我们的研究重点
组装用于内耳感官感知的亚细胞工具包
 查看全尺寸图像
查看全尺寸图像
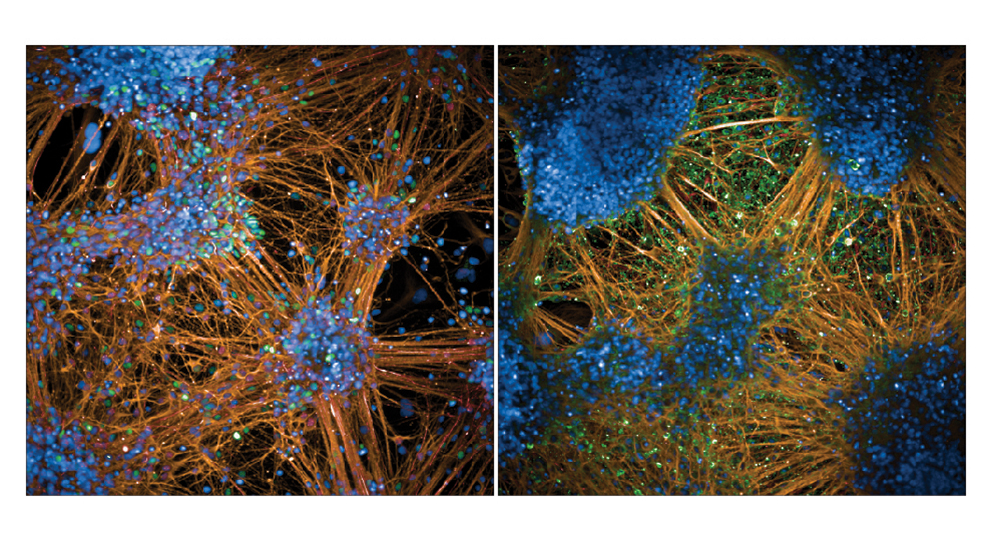
在听觉过程的最前沿,内耳半部分耳蜗中的特殊感觉细胞将声音转换为细胞信号。这些头发细胞因其顶膜上突起的巨大微绒毛束或静纤毛而得名。管束的声致挠度影响位于静纤毛尖端的阳离子通道的开放状态,使细胞去极化并触发基底部神经递质的释放结束。运动检测能力是方向敏感的,因为电流仅由束流沿上皮平面。因此,平面内静纤毛束的极化形态发生对听力至关重要两个层面:1)每个毛细胞中细胞骨架的不对称性,以静纤毛束的典型V形或半圆边缘为例,2)“平面细胞极性”(PCP)反映在相邻细胞之间的束的方向极其一致。当协调束方向时众所周知,感觉上皮需要核心PCP蛋白信号传导,其机制是在单个细胞中产生不对称性或连接不同水平的极性仍不清楚。
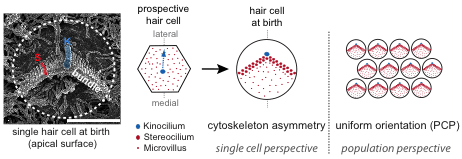
顶膜上的分子蓝图指示毛细胞的细胞骨架不对称
 查看全尺寸图像
查看全尺寸图像
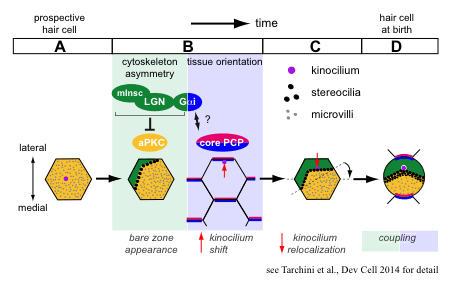
预期的毛细胞最初有一层覆盖着微绒毛的顶膜,与相邻的上皮细胞无法区分。我们专注于顶端膜的一部分,在发育过程中,在微绒毛的一个子集生长成静纤毛。这个“裸区”承载两种支架蛋白的极化定位,即哺乳动物不可分割蛋白(mInsc)和LGN(或Gpsm2),以及G蛋白质亚基Gai。这些蛋白质是产生和扩大裸区所必需的,并通过抑制极性发挥部分作用激酶aPKC连接到心尖膜的一个互补结构域。外侧mInsc、LGN和Gai以及内侧aPKC之间的界面定义了静纤毛束边缘,通常影响细胞骨架在上皮平面的不对称分布。
在毛细胞分化过程中,平面极性的最早形态学证据是初级纤毛或激毛细胞从中心到顶点的偏心位置。现有证据表明,偏心移动本身并不需要核心PCP信号,而是定向根据组织层面的提示进行翻译,以确保横向放置。我们发现,在正常毛细胞分化过程中,激毛细胞随着裸区和束的出现,平移变得越来越不偏心。我们还报告了在mInsc、LGN和Gai功能受损的这一阶段。因此,我们提出了一种新的模式,早期的翻译带来了在非常接近顶端连接处的激毛细胞后,由于细胞的膨胀而向细胞中心重新定域干预裸区。
毛细胞是如何在发育过程中整合多种极性的?
我们的研究兴趣之一是了解内在极性和组织级极性(PCP)是如何相互关联的。通过在参照静纤毛束的形成边缘,mInsc/LGN/Gai是耦合这两个平面极性水平的很好候选。有趣的是,失活Gai,而非mInsc/LGN,除了破坏每个细胞中的束不对称性外,还破坏了偏心性激肽细胞的横向定向。因此,Gai蛋白可能传递一个核心PCP信号,Gai和LGN之间特征良好的结合可能是偶联的一个中心方面机制。
在细胞内在水平上,一个研究方向是探讨未来毛细胞如何打破大多数上皮细胞的平面对称特征细胞。一个目标是确定驱动初级纤毛偏心平移的分子机制,并检查其与mInsc/LGN/Gai定义束边。在进一步的分化过程中,我们也在测试平面极性可以直接影响其他重要的细胞骨架不对称实例。其中之一是成熟发束的楼梯式结构建立高度递减的静纤毛排对声音刺激的方向敏感性很重要。
毛细胞发育研究对人类健康的意义
除了拓宽我们对细胞极性及其在内耳感觉发育中的作用的理解外,这项工作还具有重要意义为了人类健康。大多数与形成机械感觉发束有关的已知蛋白质都是通过定位与遗传相关的基因座来确定的听力损失。调节细胞骨架极性的蛋白质似乎与听力同样相关LGN公司(或每平方加仑)基因最近有报道称在无关家庭中导致非综合征性听力损失。诊断和潜在矫正遗传性听力缺陷需要建立小鼠模型,研究受影响蛋白的定位、时间需求和确切功能。
与低等脊椎动物相比,哺乳动物的毛细胞无法再生,在疾病或损伤后的生活中容易受损。也许是最治疗毛细胞丢失的有希望的治疗方法是迫使邻近的非感觉细胞采用毛细胞开发计划从头开始.尽管取得了显著进展,但前面的一个挑战是确保新生成的毛细胞进行完全的形态发生。阐明极化的方式发育过程中细胞骨架的变化将微绒毛转化为不对称的静纤毛束,调节声音感知,这无疑将有助于头发成人细胞再生策略。
{kind=link}
{kind=link}