 |
|
Ed Davis博士。 |
介绍
一个典型的哺乳动物基因通常不编码单个蛋白质,部分原因是选择性mRNA剪接现象。几乎所有哺乳动物的基因都包含多个内含子,90%以上的所有内含子基因都会进行选择性剪接,以生成多个转录变体,然后生成不同的蛋白质亚型(Pan等,2008年;Park等,2018年)。差异选择性剪接通常发生在组织内部和组织之间,但一些(5%)可能发生在个体之间(Kwan等人,2007;Wang等人,2008)。因此,一个基因可以潜在地表达许多不同的蛋白质。
在GeneCopoeia,我们为客户提供基于DNA的工具,用于许多不同类型的基因功能研究。这些包括用于开放阅读框(ORF)表达的质粒、通过CRISPR sgRNA的基因敲除、使用3'UTR的microRNA(miRNA)验证研究等,以及qPCR引物。当客户要求使用这些试剂时,他们经常会遇到每个基因的多个登录号,并且不知道需要订购哪一个。同样,客户可能对使用质粒研究特定基因感兴趣,但当被问及时,他们将不知道他们正在研究的变体或亚型的登录号。在本技术说明中,我们讨论了哺乳动物基因的多变异多样性,以及在向GeneCopoeia请求不同类型的质粒时,您正在使用的基因的登录号如何成为主要考虑因素。
基于基因的应用中不同登录号的考虑因素
如前所述,大多数哺乳动物基因都有多个外显子,并且选择性剪接,导致从单个基因表达出多个蛋白质,如图1所示。选择性剪接基因的一个例外是人类SOX2基因。截至2019年12月,美国国家生物技术信息中心(NCBI)表示,人类SOX2仅编码一个已知转录本,该转录本仅包含一个外显子(https://www.ncbi.nlm.nih.gov/nucore/NC_00003.12?report=genbank&from=181711925&to=181714436). 因此,研究人员在请求用于研究GeneCopoeia中该基因的质粒或qPCR引物时,不需要知道SOX2的登录号(正好是NM_003106.4)。很简单,对吧?
同样,人类SOX2是一个例外。下面,我们将讨论不同类型的基于基因的应用,以及它们如何受到多个转录变体的影响。
蛋白质表达的加入数考虑
研究基因功能最广泛使用的方法之一是从质粒上的ORF表达蛋白质。该质粒可用于直接转染培养细胞,或并入可感染细胞的病毒中。重要的是要记住,替代mRNA变异可以通过两种不同的方式影响质粒表达的蛋白质:1)产生的蛋白质的长度;和2)抗体识别的表位的可用性。
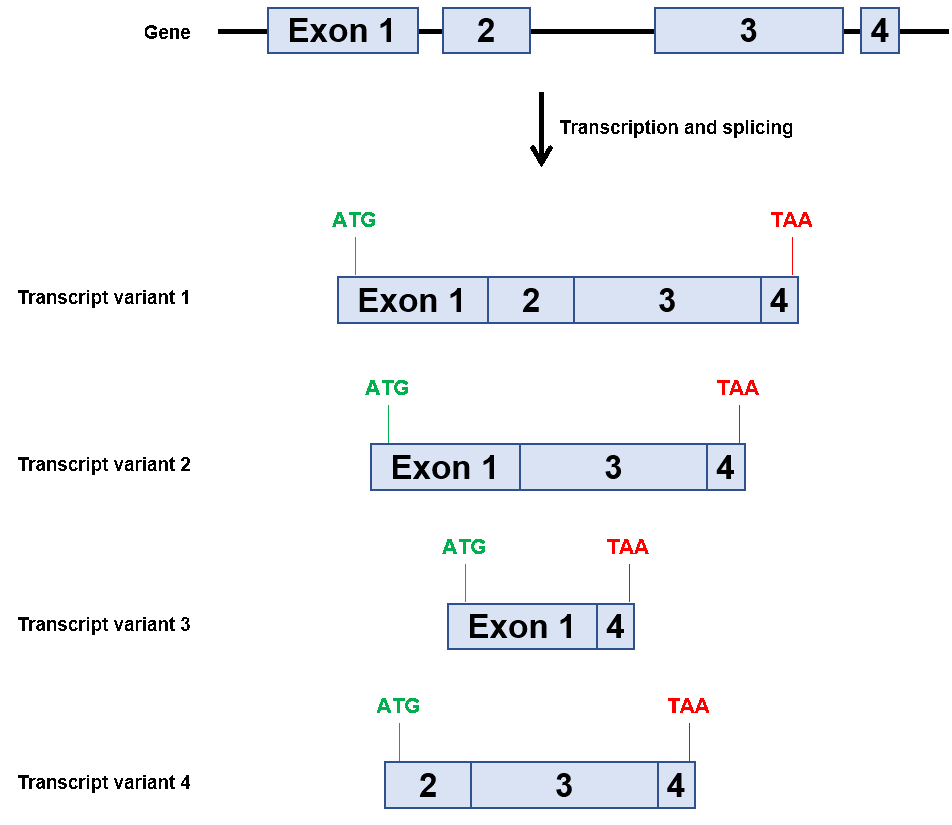
图1。假设的多xon基因的选择性剪接示例。
人类CD36就是一个受选择性剪接高度影响的基因的例子。截至2019年12月,CD36有16个已知的转录变体(https://www.ncbi.nlm.nih.gov/nucore/NC_00007.14?report=genbank&from=80602188&to=80679277). 其中一些变体仅在5'或3'UTR的长度上存在差异。然而,该基因经过选择性剪接产生6种不同的已知蛋白质,长度分别为472、433、412、396、438和317个氨基酸。其中四种亚型都具有相同的引发剂蛋氨酸,而亚型4和6具有两种不同的引发剂甲硫氨酸。所有六种蛋白质都具有相同的C末端氨基酸。
CRISPR介导淘汰赛的加入人数考虑
CRISPR的一个基本应用是基因敲除,即Cas9介导的双链断裂(DSB)以容易出错的方式进行修复,从而在蛋白质编码区产生移码突变。如果您想使用CRISPR敲除编码多种蛋白质亚型的基因,则需要考虑以下因素:
- 是否要消除所有可能的蛋白质亚型
- 你是否只想消除一种蛋白质亚型。
如果你需要敲除基因中所有已知的蛋白质亚型,那么可以设计一个sgRNA来同时敲除所有亚型。这种sgRNA的设计必须使其靶向与所有已知转录变体相同的mRNA区域。设计CRISPR sgRNAs的一种方法是尽可能以基因启动子ATG为靶点,因为基因该位置的移码突变可能会导致基因序列早期出现终止密码子并将其敲除。另外,sgRNAs也被设计用于靶向编码保守和/或已知基本功能域的序列。如果您的基因编码多个剪接变体,并且所有剪接变体都具有相同的启动子ATG或关键结构域,那么使用此策略设计单个敲除sgRNA将没有问题。
但是CD36基因的情况如何呢?如前所述,CD36用三种可能的启动子ATG编码转录变体。蛋白质亚型1、2、3和5均由具有完全相同的第一个蛋白质编码外显子的转录物编码。该外显子被排除在亚型4和6之外。此外,亚型4的第一个蛋白编码外显子与亚型1、2、3和5的蛋白编码区重叠,被排除在亚型6之外。然而,亚型6的第二个蛋白编码外显子也存在于其他五种亚型中。因此,以编码亚型6的转录物的第二个蛋白编码外显子为靶点的一个sgRNA应该足以敲除CD36的所有已知蛋白亚型。
然而,在某些情况下,您可能无法在所有转录变体中找到共同的区域。如果是这样,那么就需要使用多个sgRNA序列来同时敲除它们。
相反,您可能只想去掉其中一种亚型,而保留其他亚型。这样做需要您感兴趣的亚型包含该亚型特有的区域。就CD36而言,已知的六种亚型中没有一种包含唯一的蛋白质编码序列,因此,如果您正在处理的细胞中表达了多个亚型,则不可能单独敲除任何一种特定的蛋白质亚型。
shRNA-介导击倒的加入数考虑
shRNA介导的基因敲除在两个基本方面与CRISPR不同:1)CRISPR会导致染色体中遗传密码的永久性改变,而shRNA不会,因为前者作用于DNA,而后者作用于RNA;和2)CRISPR能够100%消除基因产物,但shRNA通常不会。然而,尽管存在这些差异,但对转录变体的考虑是相同的。在某些情况下,单个shRNA能够一次击倒一个基因的所有转录变体,而在其他情况下,需要同时使用多个shRNA。同样,只有在一个转录物变体包含唯一序列的情况下,才能选择性地敲除该变体,同时保留其他变体的完整性。
融合标签的接入号码注意事项
另一个常见的基因功能研究应用是融合标签,它通常涉及在感兴趣的蛋白质的N或C末端添加另一个蛋白质序列,如绿色荧光蛋白(GFP)或小多肽如FLAG。
融合标记通常通过两种方式之一完成。第一种方法是使用ORF表达质粒,如前所述。如果该基因表达多种蛋白质亚型,那么您需要知道您使用的是哪种亚型,以便选择正确的ORF质粒。
融合标签的第二种方法是使用CRISPR基因编辑将标签放在基因的天然染色体位点上。这种方法优于ORF质粒方法,因为它允许基因在其天然基因调控元件下表达。然而,由于存在替代的mRNA剪接,如果您想使用CRISPR在您的蛋白质上放置融合标签,您需要再次注意该基因的多个转录剪接变体和蛋白质亚型的存在,因为在某些情况下,同一蛋白质的不同亚型可能具有不同的N或C末端。人类ASXL1就是一个很好的例子。截至2019年12月,该基因编码三种已知的转录变体,以及1541、85和1480个氨基酸的相应蛋白质亚型(https://www.ncbi.nlm.nih.gov/nucore/NC_0020.11?report=genbank从32358062&到32439319). 在这三种变体中,存在两种不同的终止密码子。如果研究人员有兴趣在ASXL1上放置C末端融合标签,他们需要决定同时标记亚型1和3(共享相同的C末端氨基酸),或者标记亚型2。如果他们想使用CRISPR标记C末端的所有三种ASXL1亚型,则需要使用两套标记试剂。此外,在三种不同的变体中有两种不同的启动器ATG。其中一个在亚型1和亚型2之间共享。因此,单个N末端标记也不会同时标记所有三种亚型。然而,请注意,标记N末端并不总是一个好主意。如果蛋白质含有信号肽,则在翻译后处理过程中,位于信号肽上游的N末端标记可能会被裂解,因此可能需要在信号肽和蛋白质其余部分之间放置N末端标记
启动子研究的加入数考虑
下一种需要考虑的登录号情况涉及基因启动子研究。GeneCopoeia提供表达启动子的质粒,以控制下游报告基因的表达,如Gaussia荧光素酶或GFP。一些具有多个转录变体的基因是从单个启动子表达的。然而,其他人可能有许多潜在的推动者。其中一个例子是人类Smarca2基因。截至2019年12月,该基因有7个已知的转录变体(https://www.ncbi.nlm.nih.gov/nucore/NC_00009.12?report=genbank&from=2015347&to=2193624)和五种不同的预测启动子(https://www.genecopoeia.com/product/search3/?s=SMARCA2&search_type=1).
3’UTR研究的入学人数考虑
基因的3'非翻译区(3'UTR)包含重要的调控元件,包括多聚腺苷酸化信号和微RNA(miRNAs)的结合位点。GeneCopoeia还提供位于报告基因下游的3’UTR质粒,如Gaussia荧光素酶。这些3'UTR报告质粒还受到多个转录变体的影响,因为来自同一基因的不同转录物可能具有不同的3'UTRs。人类ASXL1基因也是一个很好的例子。如上所述,该基因有三种不同的已知转录变体,在这些转录物中至少存在两种不同的3’UTR。
qPCR引物的接入数考虑
受多种转录变体影响的基因功能研究工具的最后一个例子是qPCR引物对,用于测量细胞中单个基因产生的mRNA的相对丰度。GeneCopoire的基因qPCR引物通常设计用于识别仅存在于成熟剪接mRNA中的外显子-外显子连接。因此,与其他应用一样,qPCR引物可能设计用于扩增所有转录变体,或仅扩增一种特定的转录变体。
对于前面提到的CD36基因,可以设计一对或多对qPCR引物,同时扩增编码所有已知蛋白质亚型的转录变体。相反,一对引物可以设计成只扩增编码四种亚型的转录变体,而不能扩增其他两种亚型,而一对引子可以设计成仅扩增编码亚型2的转录变体。
结论
生物体的表型特征不仅受其基因组中基因的同一性影响,还受遗传位点中发生的选择性剪接变异所引起的多样性影响。虽然这种多样性具有明显的积极进化优势,但它也是基于基因的研究中潜在的混淆和错误来源。因此,了解您正在研究的基因可能表达多种蛋白质亚型,并且您应该了解您正在使用的亚型的身份,这一点非常重要。在GeneCopoeia,我们的科学家在哺乳动物系统的功能基因组应用方面拥有丰富的专业知识,可以帮助您找到适合您研究要求的基于基因的产品。我们提供序列验证的质粒DNA、纯化慢病毒或纯化腺相关病毒,甚至提供完整的、从开始到完成的定制细胞系构建服务。
想了解更多关于GeneCopoeia基于基因的产品和服务的信息或下单吗?访问我们的网站,致电1-301-762-0888,或发送电子邮件inquiry@genecopoeia.com.
工具书类
- 关,等。(2007年)。人类基因组中选择性剪接的遗传性。基因组研究 17, 1210.
- 平移,等., 2008. 通过高通量测序深入调查人类转录组中的选择性剪接复杂性。自然遗传学 40, 1413.
- 公园,等. (2018). 人类种群中选择性剪接变异的扩展前景。美国人类遗传学杂志 102, 11.
- 王,等. (2008). 人类组织转录体中的替代亚型调控。自然 456, 470.
版权所有©2019 GeneCopoeia,Inc。
电子邮件:inquiry@genecopoeia.com
电话:+1(866)360-9531
TNGE10-120319号 |