材料和方法
抗体。
使用以下抗体:Tau Dako A0024;磷酸-Tau(S202)CP13细胞信号技术11834;磷酸化Tau(S396)PHF13细胞信号技术9632;ROCK1 Abcam ab45171;ROCK2 Abcam ab56661;mTOR细胞信号技术2972s;phospho-mTOR(S2448)细胞信号技术2971s;磷酸化S6K(T389)细胞信号转导技术108D2;S6K信令技术2708;p62 Abcam ab109012;LC3 Novus NB100-2220;肌动蛋白抗体ab6276。
人脑组织制备和免疫印迹。
从埃默里大学阿尔茨海默病研究中心的脑库中选择来自对照组、PSP和CBD病例的死后额叶皮层组织进行比较。本研究中的PSP和CBD病例根据既定标准进行了诊断所需的广泛神经病理学特征分析(Hauw等人,1994年;凯恩斯等人,2007年). 病例尽可能与死亡年龄、性别和死亡间隔时间相匹配(表1). Sarkosyl可溶和不可溶部分的制备与前面所述完全相同(Diner等人,2014年). 蛋白质浓度采用双钦酸法(Pierce)测定。使用前面描述的标准程序进行免疫印迹(Herskowitz等人,2011年). 每个样品50微克蛋白质用于免疫印迹。肌动蛋白被用作负荷控制。使用奥德赛图像站(Li-Cor)拍摄图像,并使用奥德塞应用软件3.0版(Li-Cor)量化波段强度。
表1。
|
PMI(小时) |
发病年龄 |
死亡年龄 |
种族/性别 |
| 病例控制(n个= 10) |
|
|
|
|
| 1 |
6 |
|
58 |
WM公司 |
| 2 |
8 |
|
60 |
空军 |
| 三 |
65 |
|
WM公司 |
|
| 4 |
10 |
|
66 |
WM公司 |
| 5 |
4.5 |
|
70 |
HM公司 |
| 6 |
三 |
|
74 |
WF公司 |
| 7 |
6.5 |
|
46 |
WF公司 |
| 8 |
6.5 |
|
53 |
上午 |
| 9 |
17 |
|
57 |
空军 |
| 10 |
10 |
|
57 |
WM公司 |
| PSP公司(n个=9) |
|
|
|
|
| 1 |
15 |
61 |
70 |
WF公司 |
| 2 |
11.5 |
74 |
82 |
WF公司 |
| 三 |
23 |
77 |
83 |
WM公司 |
| 4 |
10 |
65 |
75 |
WM公司 |
| 5 |
14 |
89 |
94 |
WF公司 |
| 6 |
10 |
70 |
83 |
WM公司 |
| 7 |
6 |
65 |
71 |
WF公司 |
| 8 |
三 |
51 |
61 |
WM公司 |
| 9 |
10 |
70 |
83 |
WM公司 |
| 中央商务区(n个= 9) |
|
|
|
|
| 1 |
24 |
91 |
M(M) |
|
| 2 |
8.5 |
72 |
78 |
WF公司 |
| 三 |
5 |
57 |
62 |
WM公司 |
| 4 |
2.75 |
56 |
61 |
HM公司 |
| 5 |
24 |
33 |
44 |
WM公司 |
| 6 |
<8 |
53 |
65 |
WM公司 |
| 7 |
三 |
72 |
82 |
WF公司 |
| 8 |
三 |
63 |
68 |
WF公司 |
| 9 |
4 |
78 |
86 |
WF公司 |
为了进行密度测定分析,将对照人脑样本随机分为两组(1-5和6-10)。对照样品1-5对所有PSP病例进行免疫印迹,对所有CBD病例进行单独免疫印迹。同时,对所有PSP病例的6-10份对照样品进行免疫印迹,对所有CBD病例进行单独免疫印迹。对于每个免疫印迹,对照样品的密度测量值被平均并归一化为1的任意单位。因为每个对照病例都使用了两个免疫印迹,所以每个对照病例获得了两个密度测定值。对每个对照样品的两次密度测定值取平均值,该平均值代表密度测定图中表示的单个数据点图1.
图1。
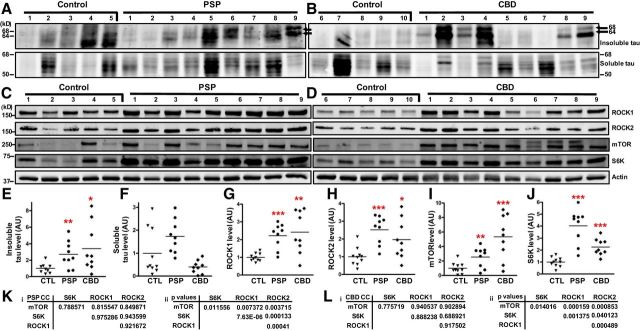
PSP和CBD大脑中ROCK、mTOR和S6K增加。A–D、来自对照组(CTL)、PSP或CBD大脑的代表性免疫印迹。E–J公司,免疫印迹密度分析。每种情况都表示为一个单独的数据点,直线表示平均值。E类, *第页= 0.0258, **第页= 0.0080.G公司, **第页= 0.0018, ***第页≤ 0.0001.H(H), *第页= 0.0235, ***第页= 0.0001.我, **第页= 0.0073, ***第页= 0.0004.J型***第页< 0.0001.K(K),L(左),相关系数(我)和第页值(ii(ii))用于PSP和CBD案例。澳大利亚,任意单位。
细胞培养、转导和转染。
在MEM Eagle(Lonza)中用10%胎牛血清和1%青霉素/链霉素保存SH-SY5Y人神经母细胞瘤细胞。对于转导或转染,等量的细胞被电镀。根据制造商的说明,分别使用Lipofectamine 2000(Invitrogen)或DharmaFECT 1 siRNA转染试剂(Dharmacon)进行DNA质粒或siRNA转导。从胚胎第17天C57BL/6小鼠胚胎中制备初级皮层神经元培养物,并将其保存在添加0.8 m米
我-如前所述的谷氨酰胺和B27(Herskowitz等人,2013年).
DNA构建体、慢病毒和siRNA。
shRNA表达慢病毒载体的构建和生成如前所述(Herskowitz等人,2012年):ROCK1 shRNA1,5′-GCCAATACTTACTAGA和shRNA2,5′-CTACAAGTGTTGCTAGTT;ROCK2 shRNA1,5′-ATCAGACAGCATCCTTTCT和shRNA2,5′-GCAAAATCTGTTAACTCG;和加扰,5′-GGACTACTCTAGACGTATA。如前所述,产生了抗shRNA1序列的ROCK2突变体(Herskowitz等人,2013年). 为了生成ROCK2 K121G,以编码shRNA-resistance突变的人ROCK2的cDNA为模板,使用QuikChange XL Site-Directed Mutagenesis kit(Stratagene):正义引物、5′-GGCATCGCAGAAGGTTTTATGCGTAGTTAGTTTTGA和反义引物5′-TCAAACTTAGAGAGCC。通过测序验证结构。对于S6K,使用了以下内容:siRNA1,5′-CAUGGAACAUGUGUGAAA和siRNA2,5′-GGAAUGGGAUAAGUUA。
化学品。
抑制ROCKs、Fasudil(HA-1077;Sigma-Aldrich)或SR3677(N个-[2-[2-(二甲氨基)乙氧基]-4-(1H(H)-吡唑-4-基)苯基-2,3-二氢-1,4-苯并二毒素-2-甲酰胺盐酸盐;Feng等人,2008年)在H中溶解2O、 模仿是H2O.MG132(碳苯氧基-我-亮氨酸-我-亮氨酸-我-将亮氨酸)、巴非霉素和雷帕霉素(分别为西格玛目录#C2211、#B1793和#R0395)溶解在100%二甲基亚砜(DMSO)中,并在10μ米,100牛顿米,或5μ米分别是。模拟的是二甲基亚砜。
细胞裂解物的制备和活性测定。
细胞在PBS+蛋白酶抑制剂混合物(罗氏诊断)、哈尔特磷酸酶抑制剂混合物(皮尔斯)和含有0.5%诺奈德P-40、0.5%脱氧胆酸盐、150 m米氯化钠,和50 m米Tris,pH 7.4。细胞裂解液进行13000 rpm的旋转,清除的裂解液被指定为可溶部分。如上所述制备肉桂糖不溶性组分用于人脑组织分析;然而,没有使用Dounce均质机(Diner等人,2014年). 如上所述,测定所有组分的蛋白质浓度。按照制造商的说明,使用CellTiter 96非放射性细胞增殖试验(Promega)测量细胞活力。在570 nm处测量吸光度,在650 nm处测量参考波长。细胞死亡值计算为对照吸光度的百分比。
qRT-PCR。
按照制造商的说明,使用TRIzol试剂(生命技术15596-026)制备RNA。cDNA是用SuperScript III第一链合成系统(Life Technologies 18080-051)产生的。使用LightCycler 480 Probes master mix(Roche)和TaqMan基因表达分析对人类Tau(Hs00902194_m1)、GAPDH(Hs02758991_g1)、啮齿动物MAPT(Rn00691532_m1)和β-肌动蛋白(Mm00607939_s1)进行cDNA定量。在Roche LightCycler 480上进行扩增。使用ΔΔCt方法对数据进行量化(用户公告2;Applied Biosystems)。
果蝇属研究。
表达人类tau的苍蝇来自布鲁明顿果蝇种群中心(51360只)。它们在Jazzmix介质(Thermo Fisher Scientific)上保持在25°C和70%湿度。对于药物治疗,苍蝇生长在配方4-24速溶培养基(卡罗来纳州生物供应)上,该培养基用水(模拟)或指定的药物剂量配制。使用配备环形灯(徕卡)的S8APO解剖显微镜检查眼睛表型,并使用DFC295相机(徕卡相机)和徕卡软件拍摄图像。使用公布的协议在NIH ImageJ中分析眼睛图像(Costa等人,2011年). 按照上述方法制备苍蝇头匀浆,用于可溶性部分细胞裂解液。
统计分析。
使用Student’st吨独立样本测试。数据表示为相对于对照的平均值±SEM的百分比。误差条代表SEM。Prism软件(GraphPad软件)用于相关性分析和生成所有图形、相关性和第页值矩阵。
结果
为了测量研究病例中的τ水平,使用一步分离方案从10个对照组、9个PSP和9个CBD死后皮质组织匀浆中生成Sarkosyl可溶性和不溶性部分(表1). 对所有组分进行SDS-PAGE,然后使用针对243–441氨基酸产生的多克隆tau抗体进行免疫印迹,该氨基酸含有微管结合4R(图1A类,B类). 在Sarkosyl不溶性PSP和CBD组分中,在68和64kDa处显著观察到tau免疫反应性,代表过度磷酸化的全长4R-tau亚型(刘等人,2001). 密度分析表明,与对照组相比,PSP和CBD患者的Sarkosyl不溶性tau水平显著升高(图1E类). 为了量化Sarkosyl可溶性τ物种,测量了50-68kDa的免疫反应带,但在对照组、PSP和CBD样品中,可溶性τ水平的变化在统计学上不显著(图1F类).
Tau降解可以通过自噬介导,通过S6K的mTOR信号可以在神经元中启动自噬级联(Boland等人,2008年;Krüger等人,2012年). 此外,ROCK1和/或ROCK2被认为影响哺乳动物细胞的自噬(Bauer等人,2009年). 为了评估PSP或CBD中这些激酶的蛋白水平是否发生变化,对Sarkosyl可溶性样品进行SDS-PAGE和随后的免疫印迹(图1C类,D类). 密度分析表明,与对照组相比,PSP和CBD患者的ROCK1、ROCK2、mTOR和S6K显著增加(图1G–J型). 进行了统计相关分析,相关系数和第页这些值表明PSP和CBD病例中ROCK1、ROCK2、mTOR和S6K之间存在强正相关(图1K(K),L(左)).
接下来,我们试图确定这些激酶如何影响细胞模型中的τ水平。首先,用表达ROCK1靶向、ROCK2靶向或干扰shRNA(每个靶点两个shRNA序列)的慢病毒转导SH-SY5Y人神经母细胞瘤细胞,96小时后,分别用免疫印迹或定量PCR检测内源性tau蛋白和tau mRNA的水平。在ROCK1或ROCK2敲除细胞中,Tau蛋白减少了34%和65%,而Tau mRNA分别减少了~51%和~61%,这表明较低的Tau mRNA水平可能导致Tau蛋白的丢失(图2A类,B类).
图2。
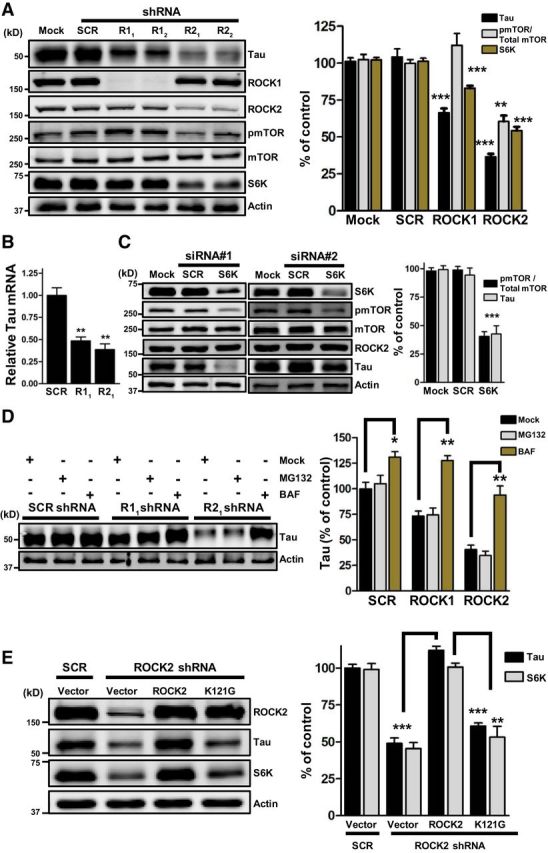
ROCK2对τ的作用需要ROCK2激酶活性。与模拟或干扰(SCR)对照相比,激酶敲除的代表性免疫印迹。除非另有说明,n个=每种条件下3次生物复制。A类,ROCK1 shRNA(R11和R12)还原τ(***第页=0.0003)和S6K(***第页= 0.0005). 岩石2 shRNA(R21和R22)减少τ,S6K(***第页<0.0001)和pMTOR(归一化为总mTOR**第页= 0.0016).B类,R1级1(**第页=0.005)和R22(**第页=0.0026)降低相对tau mRNA水平。n个=每种条件下6次生物复制。C类,S6K击倒减少pmTOR和tau(***第页< 0.0001).D类,Tau在巴非霉素处理的样品中积累。可控硅*第页= 0.0228; R1级1, **第页=0.0013;R2级1, **第页= 0.0057.E类与R2相比,ROCK2表达逆转了S6K和tau水平1加矢量控制(***第页= 0.0002, ***第页=0.0004)或L121G(**第页= 0.0039, ***第页= 0.0001). 所有数据均表示为平均值±SEM的百分比。
丝氨酸2448处mTOR的磷酸化(pmTOR)抑制自噬诱导,pmTOR水平可通过AKT或S6K的直接磷酸化通过磷脂酰肌醇3-激酶信号调节(蒋介石和亚伯拉罕,2005年;霍尔茨和布莱斯,2005年). 密度分析表明,在RNAi缺失ROCK1或ROCK2后,S6K分别下降了17%和45%,ROCK2而非ROCK1敲除降低了pmTOR,而总mTOR水平没有变化(图2A类). 基于此,我们假设ROCK2的缺失降低了S6K水平,进而抑制pmTOR,刺激自噬和τ降解。为了验证假设的前半部分,用S6K靶向或干扰siRNA转染SH-SY5Y细胞,96小时后,用免疫印迹法测定tau水平和pmTOR(图2C类). 密度分析表明,在S6K缺失细胞中,tau和pmTOR分别减少了57%和60%。值得注意的是,在S6K击倒后,ROCK2水平保持不变(图2C类). 这些结果表明,ROCK2击倒降低了S6K水平,进而降低了pmTOR和tau。接下来,用蛋白酶体抑制剂MG132处理ROCK1靶向、ROCK2靶向或干扰shRNA-表达细胞24小时,或用抑制自噬的内体酸化抑制剂bafilomycin处理6小时,并用免疫印迹法测定tau水平。MG132处理没有改变tau水平,而bafilomycin显著增加了所有样品中的tau(图2D类). 这些观察结果表明,抑制自噬而不是蛋白酶体在ROCK1或ROCK2敲低细胞中积累tau蛋白。
为了确定ROCK2激酶活性是否介导ROCK2对S6K和tau的影响,使用定点突变以甘氨酸替代赖氨酸121,生成ROCK2突变株K121G。赖氨酸121是激酶结构域中的ATP-结合位点,是ROCK2活性所必需的(Amano等人,1997年). 用表达ROCK2靶向或干扰shRNA的慢病毒转染SH-SY5Y细胞,72小时后,用指示的ROCK2质粒(抗shRNA)或空载体对照瞬时转染细胞。在ROCK2缺失细胞中,ROCK2的表达逆转了S6K和tau的作用,而K121G没有(图2E类). 这些结果表明,ROCK2对S6K和tau的作用需要ROCK2激酶活性。
为了测试药物抑制ROCK2是否刺激自噬并降低τ水平,将小鼠初级皮层神经元暴露于SR3677,一种特征化的ROCK2小分子抑制剂(Feng等人,2008年;Herskowitz等人,2013年). 16小时后,用免疫印迹法测定内源性蛋白(图3A类). 密度测定分析表明,SR3677治疗显示可溶性tau、S6K和pmTOR蛋白水平呈线性剂量依赖性下降,这与ROCK2敲除研究一致(图3A类). 抑制S6K苏氨酸389(pS6K)mTOR磷酸化诱导神经元自噬;因此,测定了pS6K、LC3-II(自噬体的特异性标记)和p62(已知的自噬底物)(Kabeya等人,2000年;Björkoy等人,2005年;Boland等人,2008年). SR3677治疗增加了LC3-II,而p62和pS6K水平则呈剂量依赖性降低(图3A类). 还制备了Sarkosyl不溶性部分,并通过免疫印迹测定了tau。密度分析表明,SR3677处理显示Sarkosyl不溶性τ水平呈线性剂量依赖性下降,这与SR3677介导的对可溶性τ水平的影响相似(图3A类). 值得注意的是,在Sarkosyl不可溶组分的两条带上显著观察到tau免疫反应,这可能代表磷酸化的tau亚型。
图3。
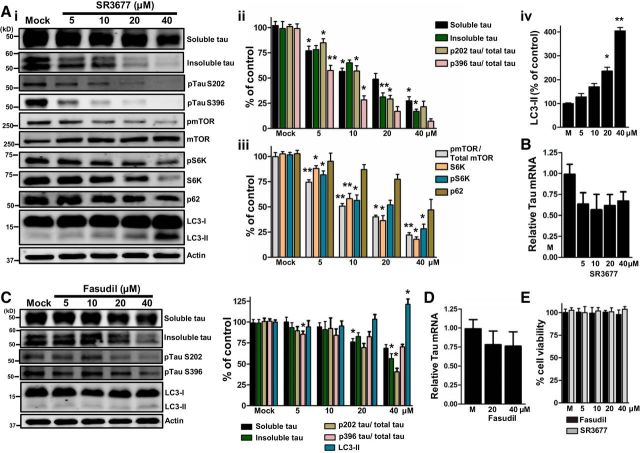
药物抑制ROCKs可降低神经元中的tau。除非另有说明,n个=每种条件下3次生物复制。tau和mTOR磷酸化的测量值分别归一化为总tau和m TOR水平。的统计数据A类,ii–iv、和C类位于表2.艾岛,显示了具有代表性的斑点。艾伊可溶性τ、不溶性τ和τ磷酸化的密度分析。Aiii公司,SR3677治疗后自噬相关标记物的密度测定分析。Aiv公司,SR3677治疗后LC3-II的密度测定分析。B类,SR3677降低tau mRNA水平(5μ米, 36%; 10 μ米, 43%; 20微米米,38%;40 μ米, 32%).n个=每种条件下6次生物复制。C类,显示了具有代表性的斑点。Fasudil治疗后可溶性τ、不溶性τ、τ磷酸化和LC3-II水平的密度分析。D类,Fasudil降低tau mRNA水平(20μ米, 22%; 40 μ米, 24%).n个=每种条件下6次生物复制。E类,在指定剂量的Fasudil或SR3677处理下,未观察到细胞活力的显著变化。所有数据均表示为相对于模拟的平均值±SEM的百分比。
过去的研究表明,ROCK的药理学抑制可以抑制丝氨酸202(pTau S202)和丝氨酸396(pTau S396;Castro-Alvarez等人,2011年;Hamano等人,2012年). 为了确定SR3677是否降低初级神经元内源性pTau S202或pTau S396,进行了免疫印迹。将pTau S202和pTau S396的水平归一化为可溶性tau,密度测定分析表明,暴露于SR3677导致pTau S202和pTiu S396呈线性剂量依赖性下降(图3A类).
定量PCR分析显示,与模拟相比,SR3677处理后tau mRNA水平较低,但没有显著降低(图3B类). 自噬可介导蛋白质聚集体降解(Verhoef等人,2002年). 基于这一点和本文的研究结果,我们假设ROCK2抑制刺激自噬,并且这种机制结合较低的tau mRNA水平有助于神经元中可溶性和不溶性tau蛋白的减少。
为了评估我们假设的临床相关性,我们将皮质神经元暴露在临床批准的ROCK抑制剂Fasudil的递增剂量下16小时(涉谷等人,2005年). 密度分析表明,当剂量≥20μ米而不溶性tau和pTau S202在40μ米(图3C类). 值得注意的是,LC3-II在40μ米与模拟相比(图3C类). 定量PCR分析显示,Fasudil≥20μ米(图3D类). 值得注意的是,Fasudil或SR3677治疗不会影响原代神经元培养的细胞存活率(图3E类). SR3677和Fasudil处理的密度测定分析的详细统计信息见表2.
表2。
SR3677和Fasudil治疗的免疫印迹测量和统计
| 化合物 |
可溶性τ |
不溶性τ |
p202陶/陶 |
p396τ/τ |
LC3-II型 |
pmTOR/mTOR |
60万 |
pS6K系列 |
第62页 |
| SR3677型 |
|
|
|
|
|
|
|
|
|
| 嘲弄 |
|
|
|
|
|
|
|
|
|
| % |
102.1 |
99.06 |
101.2 |
99.12 |
99.78 |
99.74 |
102.5 |
101.6 |
102.7 |
| 5 μ米
|
|
|
|
|
|
|
|
|
|
| % |
77.05 |
78.22 |
84.94 |
57.48 |
127.6 |
74.47 |
88.12 |
81.82 |
95.17 |
| 模拟vs 5μ米
|
*第页= 0.0144 |
第页= 0.061 |
*第页= 0.0234 |
**第页= 0.0039 |
第页= 0.1333 |
**第页= 0.006 |
*第页= 0.0169 |
*第页= 0.0129 |
第页= 0.4411 |
| 10 μ米
|
|
|
|
|
|
|
|
|
|
| % |
56.48 |
65.13 |
56.88 |
28.39 |
170.7 |
50.78 |
58.2 |
56.46 |
87.01 |
| 5μvs 10μ米
|
*第页= 0.0220 |
第页= 0.0574 |
*第页= 0.0144 |
*第页= 0.0111 |
第页= 0.0938 |
**第页= 0.0026 |
**第页= 0.0069 |
*第页= 0.0192 |
第页= 0.4378 |
| 20微米米
|
|
|
|
|
|
|
|
|
|
| % |
48.82 |
31.21 |
29.24 |
17.03 |
235.5 |
40.12 |
36.37 |
51.9 |
77.48 |
| 10μvs 20μ米
|
第页= 0.3122 |
**第页= 0.0024 |
*第页= 0.0125 |
第页= 0.1343 |
*第页= 0.0369 |
*第页=0.0249 |
*第页=0.0388 |
第页= 0.5571 |
第页= 0.2329 |
| 40 μ米
|
|
|
|
|
|
|
|
|
|
| % |
27.37 |
16.91 |
21.49 |
7.267 |
404.5 |
22.08 |
17.89 |
28.28 |
47.04 |
| 20μvs 40μ米
|
*第页= 0.0378 |
*第页= 0.0356 |
第页= 0.3055 |
第页= 0.1319 |
**第页= 0.0015 |
**第页= 0.0031 |
*第页= 0.0297 |
*第页= 0.0228 |
第页= 0.0561 |
| 法舒地尔 |
|
|
|
|
|
|
|
|
|
| 嘲弄 |
|
|
|
|
|
|
|
|
|
| % |
98.91 |
98.58 |
100.7 |
101.1 |
99.78 |
|
|
|
|
| 5 μ米
|
|
|
|
|
|
|
|
|
|
| % |
99.96 |
93.34 |
89.54 |
85.42 |
94.21 |
|
|
|
|
| 模拟vs 5μ米
|
第页= 0.889 |
第页= 0.5146 |
第页= 0.1534 |
*第页= 0.0268 |
第页= 0.5225 |
|
|
|
|
| 10 μ米
|
|
|
|
|
|
|
|
|
|
| % |
94.16 |
90.79 |
92.41 |
84.17 |
95.13 |
|
|
|
|
| 5μvs 10μ米
|
第页= 0.4891 |
第页= 0.8351 |
第页= 0.8415 |
第页= 0.8888 |
第页= 0.9285 |
|
|
|
|
| 20微米米
|
|
|
|
|
|
|
|
|
|
| % |
76.11 |
82.74 |
69.54 |
82.51 |
103.4 |
|
|
|
|
| 10μvs 20μ米
|
*第页= 0.049 |
第页= 0.5014 |
第页=0.1629 |
第页=0.8698 |
第页= 0.3743 |
|
|
|
|
| 40 μ米
|
|
|
|
|
|
|
|
|
|
| % |
68.54 |
56.45 |
40.66 |
70.28 |
121.2 |
|
|
|
|
| 20μvs 40μ米
|
第页= 0.2758 |
*第页= 0.024 |
*第页= 0.0104 |
第页= 0.1406 |
*第页= 0.0368#
|
|
|
|
|
确定法舒地尔是否能抑制致病性tau体内,一个果蝇属采用tau病模型。在这个模型中,玻璃多报告元件驱动编码眼睛中4R人类野生型tau的单拷贝cDNA的表达(称为gl-tau)。gl-tau眼睛的前部表现出粗糙的表型,视网膜中积聚了大量自噬空泡,表明自噬受到损害(Jackson等人,2002年;Bakhoum等人,2014年).果蝇属表达ROCK(DROCK)的单一亚型,DROCK与啮齿类ROCK的氨基酸序列比对表明DROCK激酶结构域中ATP结合位点附近存在错配(Mizuno等人,1999年). 基于此,法舒地尔被选为药物抑制DROCK。雷帕霉素诱导自噬途径并降低gl-tau苍蝇的粗糙眼表型(Bakhoum等人,2014年). 因此,在我们的实验中,雷帕霉素被用作阳性对照。gl-tau苍蝇在30μ米Fasudil或5μ米雷帕霉素,羽化后立即采集成虫头部进行显微镜和生化分析。与模拟对照组相比,法索地尔和雷帕霉素治疗组的粗糙眼表型也得到了类似的缓解(图4A类). 同时,免疫印迹和随后的密度测定分析表明,暴露于法舒地尔和雷帕霉素的果蝇4R人tau蛋白水平分别降低了62%和64%(图4B类). 其他生化评估表明,雷帕霉素处理降低了Ref(2)P、果蝇属哺乳动物p62的同源物,以及抑制的磷酸化果蝇属雷帕霉素(pTOR)在gl-tau苍蝇中的靶点(Miron等人,2003年;Nezis等人,2008年). 在暴露于法舒地尔的队列中,Ref(2)P和pTOR同样降低(图4B类). 基于这些发现,我们认为Fasudil可诱导自噬途径,从而降低gl-tau蝇体内致病性tau水平。
图4。
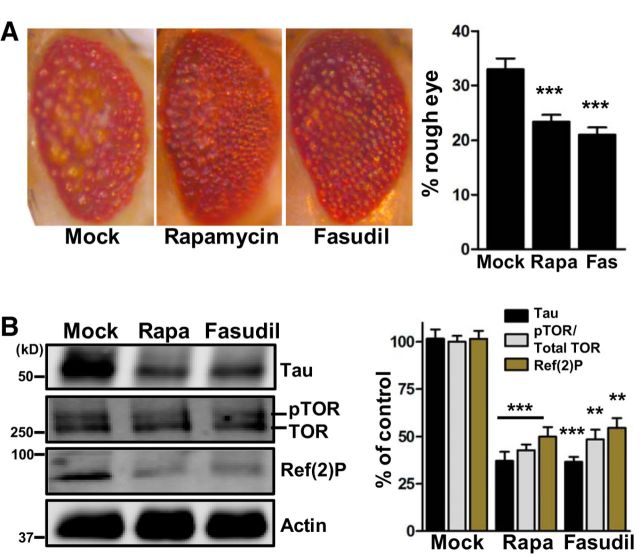
法舒地尔降低致病性τ是果蝇属tau病模型。A类,所示为典型的光学显微镜图像。雷帕霉素(Rapa)和法舒地尔(FAS)使粗糙眼表型降低33%(***第页=0.0002)和36%(***第页<0.0001)与模拟相比。B类,显示了典型的免疫印迹。雷帕霉素和法舒地尔降低4Rτ64%(***第页=0.0007)和65%(***第页=0.0003)。pTOR(归一化为总TOR)减少了57%(***第页=0.0002)和51%(***第页=0.0010)。参考(2)P减少51%(**第页=0.0014)和45%(**第页=0.0021)。n个=每种情况25–31只苍蝇。M、 模拟。所有数据均以平均值±SEM相对于模型的百分比表示。
讨论
降低致病性τ水平是治疗τ病的合理策略,但缺乏临床可用药物的治疗靶点。目前,29种小分子蛋白激酶抑制剂被用于治疗人类疾病。其中,Fasudil和Ripasudil是ROCK抑制剂。我们的研究结果表明,在PSP和CBD大脑中,ROCK1和ROCK2升高,shRNA-介导的ROCKs耗竭或药物抑制通过mRNA和蛋白质水平的组合机制降低tau。
增加的ROCK1、ROCK2、S6K和/或mTOR是否有助于潜在的致病机制或在PSP和CBD进展过程中表现出来是一个关键问题。在疾病早期对PSP和CBD大脑进行分析或在τ蛋白病小鼠模型中进行补充实验可能有助于解决这些问题。激酶水平升高可能部分归因于PSP和/或CBD脑中的反应性胶质增生(多哥和迪克森,2002年). 因此,重要的是要确定ROCKs、S6K或mTOR的变化是否仅限于特定的细胞群体,以及基因转录增强、蛋白质降解效率低下或这些效应的组合是否会提高细胞水平上的激酶。
在人类神经母细胞瘤细胞中,ROCK1敲除使tau mRNA降低~51%,但tau蛋白仅降低34%(图2A类,B类). 这种脱节可能反映了tau蛋白的半衰期很长,在SH-SY5Y细胞中约为20小时(Kim等人,2014年). 因此,较低tau mRNA的影响可能需要20小时或更长时间才能通过免疫印迹观察到tau蛋白的影响。进一步推断,ROCK2敲除降低了tau mRNA~61%,减少了tau蛋白65%,表明翻译后机制也可能在tau蛋白丢失中起作用。通过阻断自噬而非蛋白酶体,挽救了tau蛋白水平,再加上pmTOR降低,我们认为自噬刺激促进了ROCK2缺失细胞中tau蛋白的降解。类似地,在神经元中使用SR3677对ROCK2进行药物抑制,可在所有剂量下同等降低tau mRNA水平,而tau蛋白水平则以剂量依赖性的方式耗尽(图3A类,B类). SR3677处理的神经元中tau蛋白水平降低,伴随着标志着自噬诱导的蛋白变化,包括pmTOR、pS6K和p62的降低以及LC3-II的增加。这些发现支持我们的假设,即ROCK2抑制促进tau的自噬降解,但shRNA-介导的ROCKs耗竭或药物抑制如何降低tau mRNA水平是一个悬而未决的问题,需要进一步研究。值得注意的是,在PSP和CBD脑中的神经元、星形胶质细胞和少突胶质细胞中发现了tau内含物,尽管有强有力的证据支持tau降解是由神经元自噬介导的假设,但包括蛋白酶体介导的降解在内的其他机制可能对非神经元细胞中的tau处理至关重要(Wang和Mandelkow,2012年).
与具有类似效力抑制ROCK1和ROCK2的Fasudil相比,SR3677提供了大约八倍于ROCK1的ROCK2选择性(Feng等人,2008年). 这可能解释了SR3677诱导的表型更为强大以及与ROCK2敲除研究的相似性,但在本研究中使用的实验剂量下,SR3677也可能抑制ROCK1。此外,在高剂量(40μ米)在SR3677中,可能存在非靶向效应,例如直接抑制mTOR,有助于观察到的表型。SR3677的离目标命中率为1.4%,基于蜂窝的IC50对于A7r5,平滑肌细胞计算为~3n米然而,细胞培养实验中的SR3677效力可能受到细胞积累、细胞渗透性和/或ROCK1和ROCK2酶动力学的影响(Feng等人,2008年). 我们认为抑制ROCK1活性可能有治疗益处,但通过对tau mRNA水平和自噬刺激的组合效应,ROCK2抑制更能强烈地诱导tau减少。结合Fasudil的临床应用,我们的研究结果提高了人们对ROCK抑制剂的热情,将其作为对抗PSP和CBD中τ积累的潜在疗法。
脚注
本研究得到了美国国立卫生研究院/美国国家老龄研究所拨款5R00AG043552-04(J.H.H.)、美国国家神经疾病和中风研究所神经科学核心设施拨款NS055077、埃默里大学阿尔茨海默病研究中心拨款AG025688的支持,和老年痴呆症协会新研究员研究拨款2015-NIRG-339422(J.H.H.)。
工具书类
-
Amano M、Chihara K、Kimura K、Fukata Y、Nakamura N、Matsuura Y、Kaibuchi K。肌动蛋白应力纤维的形成和Rho激酶增强的局部粘连。科学。1997;275:1308–1311. doi:10.1126/science.275.5304.1308。[内政部] [公共医学] [谷歌学者]
-
An WL、Cowburn RF、Li L、Braak H、Alafuzoff I、Iqbal K、Iqbal IG、Winblad B、Pei JJ。阿尔茨海默病中磷酸化/活化p70 S6激酶的上调及其与神经原纤维病理学的关系。《美国病理学杂志》。2003;163:591–607。doi:10.1016/S0002-9440(10)63687-5。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Baker M、Litvan I、Houlden H、Adamson J、Dickson D、Perez-Tur J、Hardy J、Lynch T、Bigio E、Hutton M。tau基因中扩展单倍型与进行性核上性麻痹的关联。Hum Mol基因。1999;8:711–715. doi:10.1093/hmg/8.4.711。[内政部] [公共医学] [谷歌学者]
-
巴霍姆MF、巴霍姆CY、丁Z、卡尔顿SM、坎贝尔GA、杰克逊GR。衰老和神经变性中自噬僵局的证据。Transl Res.2014;164:1–12. doi:10.1016/j.trsl.2014.01.016。[内政部] [公共医学] [谷歌学者]
-
Bauer PO、Wong HK、Oyama F、Goswami A、Okuno M、Kino Y、Miyazaki H、Nukina N。抑制Rho激酶可增强突变型亨廷顿蛋白的降解。生物化学杂志。2009;284:13153–13164. doi:10.1074/jbc。M809229200。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Berger Z、Ravikumar B、Menzies FM、Oroz LG、Underwood BR、Pangalos MN、Schmitt I、Wullner U、Evert BO、O'Kane CJ、Rubinstein DC。雷帕霉素减轻不同聚集倾向蛋白的毒性。Hum Mol基因。2006;15:433–442. doi:10.1093/hmg/ddi458。[内政部] [公共医学] [谷歌学者]
-
Björköy G、Lamark T、Brech A、Outzen H、Perander M、Overvatn A、Stenmark H、Johansen T。p62/SQSTM1形成通过自噬降解的蛋白质聚集体,对亨廷顿蛋白诱导的细胞死亡具有保护作用。细胞生物学杂志。2005年;171:603–614. doi:10.1083/jcb.200507002。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Boland B、Kumar A、Lee S、Platt FM、Wegiel J、Yu WH、Nixon RA。神经元自噬诱导和自噬体清除:与阿尔茨海默病自噬病理学的关系。神经科学杂志。2008年;28:6926–6937. doi:10.1523/JNEUROSCI.0800-08.2008。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Cairns NJ、Bigio EH、Mackenzie IR、Neumann M、Lee VM、Hatanpaa KJ、White CL、3rd、Schneider JA、Grinberg LT、Halliday G、Duyckaerts C、Lowe JS、Holm IE、Tolnay M、Okamoto K、Yokoo H、Murayama S、Woulfe J、Munoz DG、Dickson DW、Ince PG、Trojanowski JQ、Mann DM。额颞叶变性的神经病理诊断和病理学标准:额颞叶退化联盟的共识。神经病理学学报。2007;114:5–22. doi:10.1007/s00401-007-0237-2。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Castro-Alvarez JF,Gutierrez-Vargas J,Darnaudéry M,Cardona-Gómez GP.ROCK抑制可防止全脑缺血后tau过度磷酸化和p25/CDK5增加。行为神经科学。2011;125:465–472. doi:10.1037/a0023167。[内政部] [公共医学] [谷歌学者]
-
Chiang GG,Abraham RT。雷帕霉素哺乳动物靶点(mTOR)在Ser-2448的磷酸化由p70S6激酶介导。生物化学杂志。2005年;280:25485–25490. doi:10.1074/jbc。M501707200。[内政部] [公共医学] [谷歌学者]
-
Conrad C、Andreadis A、Trojanowski JQ、Dickson DW、Kang D、Chen X、Wiederholt W、Hansen L、Masliah E、Thal LJ、Katzman R、Xia Y、Saitoh T。τ参与进行性核上性麻痹的遗传证据。Ann Neurol公司。1997;41:277–281. doi:10.1002/ana.410410222。[内政部] [公共医学] [谷歌学者]
-
Costa R,Speretta E,Crowther DC,Cardoso I.测试多西环素对阿尔茨海默病果蝇模型的治疗潜力。生物化学杂志。2011;286:41647–41655. doi:10.1074/jbc。M111.274548。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Diner I、Hales CM、Bishof I、Rabenold L、Duong DM、Yi H、Laur O、Gearing M、Troncoso J、Thambisetty M、Lah JJ、Levey AI、Seyfried NT。阿尔茨海默病中小核核糖核蛋白U1–70K的聚集特性。生物化学杂志。2014;289:35296–35313. doi:10.1074/jbc。M114.562959。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Feng Y,Yin Y,Weiser A,Griffin E,Cameron MD,Lin L,Ruiz C,Schürer SC,Inoue T,Rao PV,Schröter T,Lograsso P.发现取代的4-(吡唑-4-基)-苯并二氧六环-2-甲酰胺是强效和高选择性Rho激酶(ROCK-II)抑制剂。医学化学杂志。2008年;51:6642–6645. doi:10.1021/jm800986w。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Hamano T、Yen SH、Gendron T、Ko LW、Kuriyama M.Pitavastatin通过Rho/ROCK失活降低tau水平。神经生物老化。2012;33:2306–2320. doi:10.1016/j.neurobiolaging.2011.10.020。[内政部] [公共医学] [谷歌学者]
-
Hauw JJ、Daniel SE、Dickson D、Horoupian DS、Jellinger K、Lantos PL、McKee A、Tabaton M、Litvan I。Steele Richardson Olszewski综合征(进行性核上性麻痹)神经病学的初步NINDS神经病理学标准。1994;44:2015–2019. doi:10.1212/WNL44.11.2015。[内政部] [公共医学] [谷歌学者]
-
Herskowitz JH、Seyfried NT、Gearing M、Kahn RA、Peng J、Levey AI、Lah JJ。脂蛋白受体LR11/SORLA的Rho激酶II磷酸化改变淀粉样β的产生。生物化学杂志。2011;286:6117–6127. doi:10.1074/jbc。M110.167239。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Herskowitz JH、Offe K、Deshpande A、Kahn RA、Levey AI、Lah JJ。GGA1介导的LR11/SorLA内吞交通改变APP细胞内分布和淀粉样β的产生。分子生物学细胞。2012;23:2645–2657. doi:10.1091/mbc。E12-01-0014。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Herskowitz JH、Feng Y、Mattheyses AL、Hales CM、Higginbotham LA、Duong DM、Montine TJ、Troncoso JC、Thambisetty M、Seyfried NT、Levey AI、Lah JJ。药物抑制ROCK2抑制阿尔茨海默病小鼠模型中淀粉样β的产生。神经科学杂志。2013;33:19086–19098. doi:10.1523/JNEUROSCI.2508-13.2013。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Holz MK,Blenis J.将S6激酶1鉴定为雷帕霉素(mTOR)磷酸化激酶的新哺乳动物靶点。生物化学杂志。2005年;280:26089–26093. doi:10.1074/jbc。M504045200。[内政部] [公共医学] [谷歌学者]
-
Houlden H、Baker M、Morris HR、MacDonald N、Pickering-Brown S、Adamson J、Lees AJ、Rossor MN、Quinn NP、Kertesz A、Khan MN、Hardy J、Lantos PL、St George-Hyslop P、Munoz DG、Mann D、Lang AE、Bergeron C、Bigio EH、Litvan I、Bhatia KP、Dickson D、Wood NW、Hutton M。皮质基底细胞变性和进行性核上性麻痹具有共同的tau单倍型。神经病学。2001年;56:1702–1706. doi:10.1212/WNL.56.12.1702。[内政部] [公共医学] [谷歌学者]
-
Jackson GR、Wiedau-Pazos M、Sang TK、Wagle N、Brown CA、Masachi S、Geschwind DH。人类野生型tau与无翼路径成分相互作用,并在果蝇中产生神经原纤维病理学。神经元。2002;34:509–519. doi:10.1016/S0896-6273(02)00706-7。[内政部] [公共医学] [谷歌学者]
-
Kabeya Y、Mizushima N、Ueno T、Yamamoto A、Kirisako T、Noda T、Kominami E、Ohsumi Y、Yoshimori T。LC3是酵母Apg8p的哺乳动物同源物,加工后定位于自噬体膜。EMBO J.2000;19:5720–5728. doi:10.1093/emboj/192.15720。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Kim BM、You MH、Chen CH、Lee S、Hong Y、Hong Y、Kimchi A、Zhou XZ、Lee TH。死亡相关蛋白激酶1在异常tau蛋白调节和功能中起着关键作用。细胞死亡疾病。2014;5:e1237。doi:10.1038/cddis.2014.216。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Koch JC、Tönges L、Barski E、Michel U、Bähr M、Lingor P.ROCK2是中枢神经系统轴突变性、神经元死亡和轴突再生的主要调节器。细胞死亡疾病。2014;5:e1225。doi:10.1038/cddis.2014.191。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Kouri N、Whitwell JL、Josephs KA、Rademakers R、Dickson DW。皮质基底部变性:一种病理上独特的4Rτ蛋白病。Nat Rev Neurol公司。2011;7:263–272. doi:10.1038/nrneurol.2011.43。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Krüger U,Wang Y,Kumar S,Mandelkow EM.初级神经元tau的自噬降解及其被海藻糖增强。神经生物老化。2012;33:2291–2305. doi:10.1016/j.neurobiolaging.2011.11.009。[内政部] [公共医学] [谷歌学者]
-
Liu WK、Le TV、Adamson J、Baker M、Cookson N、Hardy J、Hutton M、Yen SH、Dickson DW。进行性核上性麻痹中扩展τ单倍型与τ生化和神经病理学的关系。Ann Neurol公司。2001年;50:494–502。doi:10.1002/ana.1159。[内政部] [公共医学] [谷歌学者]
-
Miron M,Lasko P,Sonenberg N.从Akt到FRAP/TOR的信号传递针对果蝇的4E-BP和S6K。分子细胞生物学。2003;23:9117–9126. doi:10.1128/MCB.23.24.9117-9126.2003。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Mizuno T,Amano M,Kaibuchi K,Nishida Y。果蝇Rho激酶同源物的鉴定和表征。基因。1999;238:437–444. doi:10.1016/S0378-1119(99)00351-0。[内政部] [公共医学] [谷歌学者]
-
Nakagawa O、Fujisawa K、Ishizaki T、Saito Y、Nakao K、Narumiya S.ROCK-I和ROCK-II,小鼠体内Rho-相关线圈形成蛋白丝氨酸/苏氨酸激酶的两种亚型。FEBS信函。1996;392:189–193. doi:10.1016/0014-5793(96)00811-3。[内政部] [公共医学] [谷歌学者]
-
Nezis IP、Simonsen A、Sagona AP、Finley K、Gaumer S、Contamine D、Rusten TE、Stenmark H、Brech A.参考(2)P,哺乳动物p62的果蝇同源物,是在成人大脑中形成蛋白质聚集物所必需的。细胞生物学杂志。2008年;180:1065–1071. doi:10.1083/jcb.200711108。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Shibuya M、Hirai S、Seto M、Satoh S、Ohtomo E。法舒地尔对急性缺血性卒中的影响:一项前瞻性安慰剂对照双盲试验的结果。神经科学杂志。2005年;238:31–39. doi:10.1016/j.jns.2005.06.003。[内政部] [公共医学] [谷歌学者]
-
Tang Z、Bereczki E、Zhang H、Wang S、Li C、Ji X、Branca RM、LehtiöJ、Guan Z、Filipcik P、Xu S、Winblad B、Pei JJ。雷帕霉素(mTor)的哺乳动物靶点介导τ蛋白失衡:对阿尔茨海默病的影响。生物化学杂志。2013;288:15556–15570. doi:10.1074/jbc。M112.435123。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
多哥T,Dickson DW。进行性核上性麻痹星形胶质细胞中Tau的积累是一个退化过程,而不是一个反应过程。神经病理学学报。2002;104:398–402. doi:10.1007/s00401-002-0569-x。[内政部] [公共医学] [谷歌学者]
-
Verhoef LG,Lindsten K,Masucci MG,Dantuma NP。聚集体形成抑制聚谷氨酰胺蛋白质的蛋白酶体降解。Hum Mol基因。2002;11:2689–2700. doi:10.1093/hmg/11.22.2689。[内政部] [公共医学] [谷歌学者]
-
Wang Y,Mandelkow E.通过自噬和蛋白酶体途径降解tau蛋白。生物化学Soc Trans。2012;40:644–652. doi:10.1042/BST20120071。[内政部] [公共医学] [谷歌学者]

