

集锦
-
• 关于纽卡斯尔病的历史观点和全球情景。 -
• 新城疫病毒基因组特征。 -
• 新城疫病毒的复制与致病性。 -
• 使用新城疫病毒作为疫苗载体。 -
• 新城疫病毒作为溶瘤剂。
摘要
1.历史观点
图1。
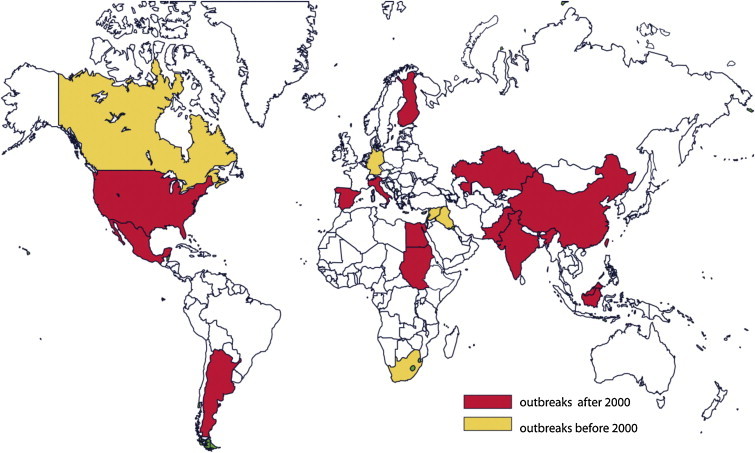
2.全球情景
图2。

3.新城疫病毒
图3。
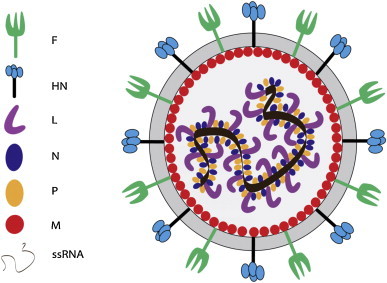
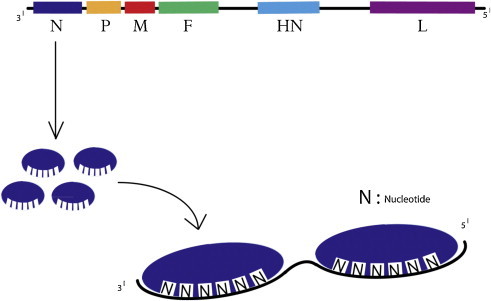
图4。
3.1. 核衣壳蛋白(N)
3.2. 磷蛋白(P)
3.3. 基质蛋白(M)
3.4. 融合蛋白(F)
3.5. 血凝素-神经氨酸酶蛋白(HN)
3.6. 大聚合酶蛋白(L)
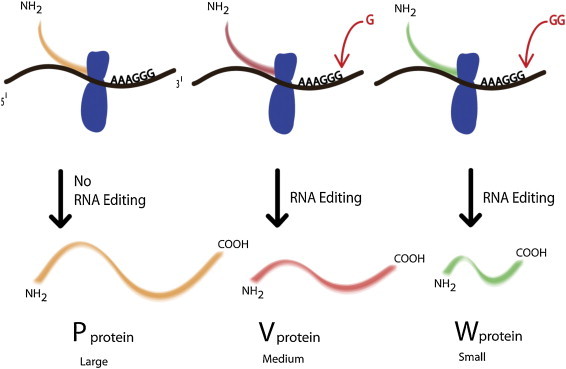
3.7. V蛋白
图5。
4.复制和致病性
图6。
5.NDV毒力和致病性研究的最新进展
6.新城疫病毒作为疫苗载体
-
一、。 NDV在鸡胚、细胞培养和鸟类和非鸟类的呼吸道中以高滴度生长。 -
二、。 NDV通过呼吸道自然感染,因此有助于传递来自呼吸道病原体的保护性抗原。 此外,它还诱导局部和全身免疫反应。 -
三、。 NDV可引起体液和细胞免疫反应。 -
四、。 NDV有一个模块化基因组,只有六个基本基因和两个辅助基因,易于操作。 -
五、。 NDV不与宿主基因组整合,因为它在宿主细胞质中复制,并且表现出最少的遗传重组。 -
不及物动词。 表达外源抗原的重组NDV经过多次传代后,外源蛋白表现出较高且稳定的表达 在体外 和 体内 ( Huang等人,2003a ). -
七、。 利用反向遗传学开发稳定的疫苗和疫苗载体可以减毒新冠病毒。 缺失突变体(V,W)和F裂解位点发生改变的突变体的发展可能同时导致更多的免疫原性和减毒重组病毒( Huang等人,2003a , Samal等人,2011年 ).
7.新城疫病毒作为溶瘤剂
8.结论
鸣谢
工具书类
-
Anon.初级工业园区水与环境部; 2013年,新城疫疫苗接种。 [ 谷歌学者 ] -
Alamares J.G.、Elankumaran S.、Samal S.K.、Iorio R.M.两株新城疫病毒V蛋白的干扰素拮抗活性与其已知毒力特性相关。 2010年《病毒研究》; 147(1):153–157. doi:10.1016/j.virusres.2009.10.020。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
澳大利亚维多利亚州Albiston H.、Gorrie C.纽卡斯尔病。 兽医。 J.1942年; 18:75–79。 [ 谷歌学者 ] -
亚历山大·D·J·纽卡斯尔病和其他禽副粘病毒。 作者:Swayne D.E.、Glisson J.、Jackwood M.W.、Pearson J.E.、Reed W.M.,编辑。 禽病原体分离和鉴定实验室手册。 第四版美国鸟类病理学家协会; 费城:1998年。 [ 谷歌学者 ] -
亚历山大·D·J·纽卡斯尔病和其他禽副粘病毒。 科学评论。 技术2000; 19(2):443–462. doi:10.20506/rst.192.1231。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Altomonte J.、Marozin S.、Schmid R.M.和Ebert O.将新城疫病毒作为一种改良的抗肝细胞癌溶瘤剂。 摩尔-热。 2010; 18(2):275–284. doi:10.1038/mt.2009.231。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Batliwalla F.M.、Bateman B.A.、Serrano D.、Murray D.、Macphail S.、Maino V.C.、Ansel J.C.、Gregersen P.K.、Armstrong C.A.对手术后使用新城疫病毒(NDV)溶瘤液治疗的AJCC III期恶性黑色素瘤患者进行15年随访,并确定CD8T细胞库中的变化。 《分子医学》1998; 4(12):783–794. [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Biswas M.、Johnson J.B.、Kumar S.R.、Parks G.D.、Elankumarana S.将宿主补体调节蛋白纳入新城疫病毒可增强补体逃避。 J.维罗尔。 2012; 86(23):12708–12716. doi:10.1128/JVI.00886-12。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Bukreyev A.,Collins P.L.纽卡斯尔病病毒作为人类疫苗载体。 货币。 操作。 摩尔-热。 2008; 10(1):46–55. [ 公共医学 ] [ 谷歌学者 ] -
Bukreyev A.、Huang Z.、Yang L.、Elankumaran S.、St Claire M.、Murphy B.R.、Samal S.K.、Collins P.L.表达外来病毒抗原的重组新城疫病毒在灵长类动物中减弱且具有高度免疫原性。 J.维罗尔。 2005; 79(21):13275–13284. doi:10.1128/JVI.79.21.13275-13284.2005。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Bukreyev A.、Skiadopoulos M.H.、Murphy B.R.、Collins P.L.非分段负链病毒作为疫苗载体。 J.维罗尔。 2006; 80(21):10293–10306. doi:10.1128/JVI.00919-06。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Calain P.,Roux L.六法则,仙台病毒缺陷干扰RNA高效复制的基本特征。《病毒杂志》。 1993; 67(8):4822–4830. doi:10.1128/jvi.67.8.4822-4830.1993。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Cantin C.、Holguera J.、Ferreira L.、Villar E.、Munoz Barroso I.新城疫病毒可能通过小窝介导的内吞作用进入细胞。 《病毒遗传学杂志》。 2007; 88(第2部分):559–569。 doi:10.1099/vir.0.82150-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Carnero E.,Li W.,Borderia A.V.,Moltedo B.,Moran T.,Garcia-Sastre A.用新城疫病毒载体优化人类免疫缺陷病毒gag的表达,以诱导有效的免疫反应。 J.维罗尔。 2009; 83(2):584–597. doi:10.1128/JVI.01443-08。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Cassel W.A.、Garrett R.E.新城疫病毒作为抗肿瘤药物。 癌症。 1965; 18:863–868. doi:10.1002/1097-0142(196507)18:7<863::aid-cncr2820180714>3.0.co; 2伏。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ch'ng W.C.,Stanbridge E.J.,Yusoff K.,Shafee N.新城疫病毒在常氧和缺氧条件下透明细胞肾癌细胞中的溶瘤活性:von Hippel-Lindau和干扰素β信号传导之间的相互作用。 2013年干扰素细胞因子研究杂志; 33(7):346–354. doi:10.1089/jir.2012.0095。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Chambers P.、Millar N.S.、Platt S.G.、Emmerson P.T.编码新城疫病毒基质蛋白基因的核苷酸序列。 1986年《核酸研究》; 14(22):9051–9061. doi:10.1093/nar/14.22.9051。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
陈凤,刘杰,刘丹,闫姿,纪杰,秦杰,李海,马杰,毕勇,谢奇。中国肉鸡繁殖群分离的新城疫病毒株的全基因组序列。 J.维罗尔。 2012; 86(22):12461–12462. doi:10.1128/JVI.02314-12。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Chen S.,Hao H.,Wang X.,Du E.,Liu H.,Yang T.,Liu Y.,Fu X.,Zhang P.,Yang-Z.从中国病猪中分离的一株无晶状体新城疫病毒株HX01的基因组特征。 病毒基因。 2013; 46(2):264–270. doi:10.1007/s11262-012-0844-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Choi K.S.、Kye S.J.、Kim J.Y.、To T.L.、Nguyen D.T.、Lee Y.J.、Choi J.G.、Kang H.M.、Kin K.I.、Song B.M.、Lee H.S.越南新城疫病毒分子流行病学。 特罗普。 阿尼姆。 2014年卫生生产; 46(1):271–277. doi:10.1007/s11250-013-0475-3。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Corey E.A.、Mirza A.M.、Levandowsky E.、Iorio R.M.纽卡斯尔病病毒血凝素-神经氨酸酶二聚体界面突变引起的融合缺陷是由于受体结合的温度依赖性缺陷所致。 J.维罗尔。 2003; 77(12):6913–6922. doi:10.1128/JVI.77.12.6913-6922.2003。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Cornax I.、Diel D.G.、Rue C.A.、Estevez C.、Yu Q.、Miller P.J.、Afonso C.L.Newcastle disease virus fusion和血凝素-神经氨酸酶蛋白有助于其巨噬细胞宿主范围。 《病毒遗传学杂志》。 2013; 94(第6部分):1189–1194。 doi:10.1099/vir.0.048579-0。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Cornelissen L.A.、de Vries R.P.、de Boer-Luijtze E.A.、Rigter A.、Rottier P.J.、de Haan C.A.可溶性重组三聚体血凝素的单一免疫保护鸡免受高致病性禽流感病毒H5N1的感染。 《公共科学图书馆·综合》。 2010; 5(5):e10645。 doi:10.1371/journal.pone.0010645。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
克劳福德M.政府兽医。 外科医生; 锡兰科伦坡:1930年。 兰尼切特。 年度报告。 [ 谷歌学者 ] -
Czegledi A.、Ujvari D.、Somogyi E.、Wehmann E.、Werner O.、Lomniczi B.禽副粘病毒血清1型(新城疫病毒)第三基因组大小类别及其进化意义。 2006年病毒研究; 120(1–2):36–48. doi:10.1016/j.virusres.2005.11.009。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
de Leeuw O.,Peeters B.新城疫病毒的完整核苷酸序列:副粘病毒亚科中存在一个新属的证据。 《病毒遗传学杂志》。 1999; 80(第1部分):131–136。 doi:10.1099/0022-1317-80-1-131。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
de Leeuw O.S.、Koch G.、Hartog L.、Ravenshorst N.、Peeters B.P.新城疫病毒的毒力由融合蛋白的切割位点以及血凝素神经氨酸酶蛋白的茎区和球状头决定。 《病毒遗传学杂志》。 2005; 86(第6部分):1759-1769。 doi:10.1099/vir.0.80822-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Deng R.,Wang Z.,Mirza A.M.,Iorio R.M.通过其同源融合蛋白尖峰促进细胞融合所需的副粘病毒附着蛋白上一个结构域的定位。 病毒学。 1995; 209(2):457–469. doi:10.1006/viro.1995.1278。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Diel D.G.、Miller P.J.、Wolf P.C.、Mickley R.M.、Musante A.R.、Emanueli D.C.、Shively K.J.、Pedersen K.、Afonso C.L.2010年美国鸬鹚和海鸥分离的新城疫病毒特征。 禽疾病。 2012; 56(1):128–133. doi:10.1637/9886-081111-Reg.1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
DiNapoli J.M.、Kotelkin A.、Yang L.、Elankumaran S.、Murphy B.R.、Samal S.K.、Collins P.L.、Bukreyev A.Newcastle disease virus,一种宿主范围受限病毒,作为新发病原体鼻内免疫的疫苗载体。 程序。 国家。 阿卡德。 科学。 美国2007年; 104(23):9788–9793. doi:10.1073/pnas.0703584104。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
DiNapoli J.M.、Nayak B.、Yang L.、Finneyfrock B.W.、Cook A.、Andersen H.、Torres-Velez F.、Murphy B.R.、Samal S.K.、Collins P.L.、Bukreyev A.表达H5N1高致病性禽流感病毒血凝素或神经氨酸酶蛋白的新城疫病毒载体疫苗可保护猴子免受病毒攻击。 J.维罗尔。 2010; 84(3):1489–1503. doi:10.1128/JVI.01946-09。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Dortmans J.C.、Koch G.、Rottier P.J.、Peeters B.P.纽卡斯尔病病毒的毒力:目前已知的情况? 兽医。 2011年决议; 42:122–130. doi:10.1186/1297-9716-42-122。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Dortmans J.C.、Rottier P.J.、Koch G.、Peeters B.P.。病毒复制复合物与新城疫病毒的毒力有关。 J.维罗尔。 2010; 84(19):10113–10120. doi:10.128/JVI.00097-10。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Doyle T.一种迄今尚未记录的禽类疾病,由滤过性病毒引起。 J.公司。 病态。 治疗。 1927:144–169. [ 谷歌学者 ] -
爱德华兹·J·兽医。 穆克特斯瓦尔研究所; 1928.一种新的家禽疾病。 3月31日,第14-15页。 [ 谷歌学者 ] -
Elankumaran S.基因工程纽卡斯尔病病毒治疗前列腺癌:一颗神奇的子弹还是一种不匹配。 抗癌治疗专家评论。 2013; 13(7):769–772. doi:10.1586/14737140.2013.811062。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Elankumaran S.、Chavan V.、Qiao D.、Shobana R.、Moorkana G.、Biswas M.、Samal S.K.溶瘤病毒治疗用I型干扰素敏感重组新城疫病毒。 J.维罗尔。 2010; 84(8):3835–3844. doi:10.1128/JVI.01553-09。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Elankumaran S.、Rockemann D.、Samal S.K.纽卡斯尔病病毒通过细胞死亡的内源性和外源性半胱天冬酶依赖途径进行溶瘤。 J.维罗尔。 2006; 80(15):7522–7534. doi:10.1128/JVI.00241-06。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Errington W.,Emmerson P.T.重组新城疫病毒核衣壳蛋白组装成核衣壳样结构受到磷蛋白的抑制。 《病毒遗传学杂志》。 1997; 78(第9部分):2335–2339。 doi:10.1099/0022-1317-78-9-2335。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ferreira H.L.、Pirlot J.F.、Reynard F.、van den Berg T.、Bublot M.、Lambrecht B.对鸡新城疫H5疫苗诱导的H5N1高致病性禽流感病毒的免疫应答和保护。 禽疾病。 2012; 56(补充4):940–948。 doi:10.1637/10148-040812-ResNote.1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Forrester N.L.、Widen S.G.、Wood T.G.、Travassos da Rosa A.P.、Ksiazek T.G.,Vasilakis N.、Tesh R.B.从印度尼西亚分离的一种新的新城疫病毒的鉴定代表了II类基因型XIII的祖先血统。 病毒基因。 2013; 47(1):168–172. doi:10.1007/s11262-013-0900-8。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Freeman A.I.、Zakay-Rones Z.、Gomori J.M.、Linetsky E.、Rasooly L.、Greenbaum E.、Rozenman-Yair S.、Panet A.、Libson E.、Irving C.S.、Galun E.、Siegal T.静脉注射NDV-HUJ溶瘤病毒治疗复发性多形性胶质母细胞瘤的I/II期试验。 摩尔-热。 2006; 13(1):221–228. doi:10.1016/j.ymthe.2005.08.016。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
葛J.、邓刚、文忠、田刚、王毅、石杰、王欣、李勇、胡S.、姜毅、杨C.、于康、布中、陈海。基于新城疫病毒的减毒活疫苗完全保护鸡和小鼠免受同源和异源H5N1禽流感病毒的致命攻击。 J.维罗尔。 2007; 81(1):150–158. doi:10.1128/JVI.01514-06。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Ghric M.,El Zowalaty M.,Omar A.R.,Ideris A.纽卡斯尔病病毒马来西亚株AF2240的血凝素-神经氨酸酶糖蛋白诱导MCF-7细胞凋亡。 昂科尔。 2013年代表; 30(3):1035–1044. doi:10.3892/或.2013.2573。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Glickman R.L.、Syddall R.J.、Iorio R.M.、Sheehan J.P.、Bratt M.A.融合糖蛋白裂解活化位点的定量基本残基要求是新城疫病毒毒力的决定因素。 J.维罗尔。 1988; 62(1):354–356. doi:10.1128/jvi.62.1.354-356.1988。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Gorman J.J.、Nestorowicz A.、Mitchell S.J.、Corino G.L.、Selleck P.W.《新城疫病毒膜糖蛋白前体蛋白水解活化位点的表征》。 生物学杂志。 化学。 1988; 263(25):12522–12531. [ 公共医学 ] [ 谷歌学者 ] -
Haddas R.、Meir R.、Perk S.、Horowitz I.、Lapin E.、Rosenbluth E.、Lublin A.以色列动物园小猫头鹰(Athene noctua)和非洲企鹅(Spheniscus demersus)的新城疫病毒。 转轨紧急疾病。 2013年doi:10.1111/tbed.12064。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Horikami S.M.、Smallwood S.、Moyer S.A.仙台病毒V蛋白与NP蛋白相互作用,调节病毒基因组RNA复制。 病毒学。 1996; 222(2):383–390. doi:10.1006/viro.1996.0435。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Hotte S.J.、Lorence R.M.、Hirte H.W.、Polawski S.R.、Bamat M.K.、O'Neil J.D.、Roberts M.S.、Groene W.S.、Major P.P.溶瘤病毒PV701的优化临床方案。 临床。 癌症研究2007; 13(3):977–985. doi:10.1158/1078-0432.CCR-06-1817。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Hu X.L.,Ray R.,Compans R.W.人类副流感病毒的融合蛋白和血凝素-神经氨酸酶之间的功能相互作用。 J.维罗尔。 1992; 66(3):1528–1534. doi:10.1128/jvi.66.3.1528-1534.1992。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
黄毅,万华强,刘华强,吴义东,刘晓峰。鹅新城疫病毒分离株基因组序列:核蛋白基因非编码区的新型六核苷酸插入。 简要报告。 架构(architecture)。 维罗尔。 2004; 149(7):1445–1457. doi:10.1007/s00705-004-0297-8。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Huang Z.、Elankumaran S.、Panda A.、Samal S.K.重组新城疫病毒作为疫苗载体。 波尔特。 科学。 2003; 82(6):899–906. doi:10.1093/ps/82.6.899。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Huang Z.、Elankumaran S.、Yunus A.S.、Samal S.K.一种表达传染性法氏囊病病毒(IBDV)VP2蛋白的重组新城疫病毒(NDV)可对抗NDV和IBDV。 J.维罗尔。 2004; 78(18):10054–10063. doi:10.128/JVI.78.18.10054-10063.2004。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Huang Z.、Krishnamurthy S.、Panda A.、Samal S.K.来自重组新城疫病毒最接近3’的基因座的外源基因的高水平表达。 《病毒遗传学杂志》。 2001; 82(第7部分):1729–1736。 doi:10.1099/0022-1317-82-7-1729。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Huang Z.、Krishnamurthy S.、Panda A.、Samal S.K.纽卡斯尔病病毒V蛋白与病毒发病相关,并作为α-干扰素拮抗剂发挥作用。 J.维罗尔。 2003; 77(16):8676–8685. doi:10.1128/JVI.77.16.8676-8685.2003。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Huang Z.、Panda A.、Elankumaran S.、Govindarajan D.、Rockemann D.D.、Samal S.K.新城疫病毒的血凝素-神经氨酸酶蛋白决定了病毒的向性和毒力。 J.维罗尔。 2004; 78(8):4176–4184. doi:10.1128/JVI.78.8.4176-4184.2004。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Iorio R.M.、Glickman R.L.、Riel A.M.、Sheehan J.P.、Bratt M.A.纽卡斯尔病病毒HN糖蛋白上七个重叠抗原位点的功能和中和特征:某些位点的单克隆抗体可防止病毒附着。 1989年《病毒研究》; 13(3):245–261. doi:10.1016/0168-1702(89)90019-1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Iorio R.M.、Mahon P.J.副粘病毒:不同受体-不同融合机制。 微生物趋势。 2008; 16(4):135-137。 doi:10.1016/j.tim.2008.01.006。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Jahanshiri F.、Eshaghi M.、Yusoff K.磷蛋白的鉴定:磷蛋白和磷蛋白:新城疫病毒的核衣壳蛋白相互作用域。 架构(architecture)。 维罗尔。 2005; 150(3):611–618. doi:10.1007/s00705-004-0439-z。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Janke M.、Peeters B.、de Leeuw O.、Moorman R.、Arnold A.、Fournier P.、Schirmacher V.带有GM-CSF插入基因编码的重组新城疫病毒(NDV)作为癌症免疫基因治疗的新载体。 基因疗法。 2007; 14(23):1639–1649。 doi:10.1038/sj.gt.3303026。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
自由生活和宠物鸟中的Kaleta E.、Baldauf C.纽卡斯尔病。 作者:Alexander D.J.,编辑。 纽卡斯尔病。 Kluwer Acedemic出版社; 马萨诸塞州波士顿:1988年。 [ 谷歌学者 ] -
Kanno T.、Ochi Y.、Hashimoto K.Dtsch。 蒂拉日尔。 Wshr。; 1929年,韩国新流感。 [ 谷歌学者 ] -
Kattenbelt J.A.、Stevens M.P.、Gould A.R.纽卡斯尔病病毒基因组中的序列变异。 2006年病毒研究; 116(1–2):168–184. doi:10.1016/j.virusres.2005.10.001。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Khattar S.K.、Collins P.L.、Samal S.K.用表达牛疱疹病毒-1(BHV-1)糖蛋白D的重组新城疫病毒免疫牛,可诱导粘膜和血清抗体反应,并对BHV-1提供部分保护。 疫苗。 2010; 28(18):3159–3170. doi:10.1016/j.vaccine.2010.02.051。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Khattar S.K.、Samal S.、Devico A.L.、Collins P.L.、Samal S.K.表达人类免疫缺陷病毒1型包膜糖蛋白的新城疫病毒在豚鼠中诱导强烈的粘膜和血清抗体反应。 J.维罗尔。 2011; 85(20):10529–10541. doi:10.1128/JVI.05050-11。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] [ 已缩回 ] -
Khattar S.K.、Samal S.、Labrange C.C.、Montefiori D.C.、Collins P.L.、Samal S.K.通过减毒新城疫病毒活载体表达的HIV-1 gp160、gp140和gp120的免疫原性比较。 《公共科学图书馆·综合》。 2013; 8(10):e78521。 doi:10.1371/journal.pone.0078521。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] [ 已缩回 ] -
Khattar S.K.、Yan Y.、Panda A.、Collins P.L.、Samal S.K.纽卡斯尔病病毒HN蛋白的Y526Q突变降低其功能活性,并减弱病毒复制和致病性。 J.维罗尔。 2009; 83(15):7779–7782. doi:10.1128/JVI.00536-09。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Kho C.L.、Tan W.S.、Tey B.T.、Yusoff K.新城疫病毒核衣壳蛋白:自组装和长度决定域。 《病毒遗传学杂志》。 2003; 84(第8部分):2163–2168。 doi:10.1099/vir.0.19107-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Kim S.H.、Xiao S.、Paldurai A.、Collins P.L.、Samal S.K.。C596在血凝素-神经氨酸酶蛋白的C末端延伸中在高毒力印尼新城疫病毒株复制和致病性中的作用。 《病毒遗传学杂志》。 2014年; 95(第2部分):331–336。 doi:10.1099/vir.0.055285-0。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] [ 已缩回 ] -
Kim S.H.,Yan Y.,Samal S.K.新城疫病毒血凝素神经氨酸酶蛋白细胞质尾部氨基酸序列在病毒粒子掺入、细胞融合和致病性中的作用。 J.维罗尔。 2009; 83(19):10250–10255. doi:10.1128/JVI.01038-09。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
孔丹、文中、苏宏、葛杰、陈伟、王欣、吴川、杨川、陈海、布中。新城疫病毒介导的尼帕脑炎疫苗在小鼠中诱导B和T细胞反应,在猪中诱导持久中和抗体。 病毒学。 2012; 432(2):327–335. doi:10.1016/j.virol.12.06.001。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Kortekaas J.、de Boer S.M.、Kant J.、Vloet R.P.、Antonis A.F.、Moormann R.J.副粘病毒疫苗载体提供的裂谷热病毒免疫。 疫苗。 2010; 28(27):4394–4401. doi:10.1016/j.疫苗.2010.04.048。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
克拉内菲尔德·F·在内德-印度,在赫特·普里姆夫斯的领导下。 迪尔根涅斯克内德·英迪施大道(Ned.Indisch Bl.Diergeneesk)。 1926; 38:448–450. [ 谷歌学者 ] -
Krishnamurthy S.、Huang Z.、Samal S.K.从克隆的cDNA中恢复新城疫病毒强毒株:外源基因的表达导致生长迟缓和衰弱。 病毒学。 2000; 278(1):168–182. doi:10.1006/viro.2000.0618。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Krishnamurthy S.、Samal S.K.纽卡斯尔病病毒Beaudette C株尾部的核苷酸序列、核衣壳蛋白基因和基因间区域以及整个基因组序列的完成。 《病毒遗传学杂志》。 1998; 79(第10部分):2419–2424。 doi:10.1099/0022-1317-79-10-2419。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Kumar S.、Nayak B.、Collins P.L.、Samal S.K.使用重组禽副粘病毒3型载体在鸡体内评估新城疫病毒F和HN蛋白的保护性免疫。 J.维罗尔。 2011; 85(13):6521–6534. doi:10.1128/JVI.00367-11。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] [ 已缩回 ] -
Lam H.Y.、Yeap S.K.、Rasoli M.、Omar A.R.、Yusoff K.、Suraini A.A.、Aliheen N.B.新城疫病毒在癌症治疗中的安全性和临床应用。 J.生物识别。 生物技术。 2011; 2011:718710. doi:10.1155/2011/718710。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Lamb R.,Parks G.副粘病毒科:病毒及其复制。 作者:Knipe D.M.、Howley P.M.、Griffin D.E.、Lamb R.A.、Martin M.A.、Roizman B.、Straus S.E.编辑。 菲尔德病毒学。 第五版:Lippincott Williams&Wilkins; 费城:2007年。 第1449-1496页。 [ 谷歌学者 ] -
Lawrence T.M.、Wanjalla C.N.、Gomme E.A.、Wirblich C.、Gatt A.、Carnero E.、Garcia-Sastre A.、Lyles D.S.、McGettigan J.P.、Schnell M.J.使用负链RNA病毒对抗人类免疫缺陷病毒1型gag的异源启动子策略比较。 《公共科学图书馆·综合》。 2013; 8(6):e67123。 doi:10.1371/journal.pone.0067123。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Lindh E.、Ek-Kommonen C.、Vaananen V.M.、Alasaari J.、Vaheri A.、Vapalahti O.、Huovilainen A.芬兰爆发相关和野生水禽源新城疫病毒株的分子流行病学,包括一个新的I类基因型。 临床杂志。 微生物。 2012; 50(11):3664–3673. doi:10.1128/JCM.01427-12。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Liu H.,Zhang P.,Wu P.,Chen S.,Mu G.,Duan X.,Hao H.,Du E.,Wang X.,Yang Z.从中国野生鸟类中分离的两种新城疫病毒的系统发育特征和毒力。 感染。 遗传学。 进化。 2013; 20C:215–224。 doi:10.1016/j.meegid.2013.08.021。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Lorence R.M.、Katubig B.B.、Reichard K.W.、Reyes H.M.、Phuangsab A.、Sassetti M.D.、Walter R.J.、Peeples M.E.局部新城疫病毒治疗后人类纤维肉瘤异种移植物的完全回归。 1994年癌症研究; 54(23):6017–6021. [ 公共医学 ] [ 谷歌学者 ] -
Lorence R.M.、Pecora A.L.、Major P.P.、Hotte S.J.、Laurie S.A.、Roberts M.S.、Groene W.S.、Bamat M.K.溶瘤病毒PV701静脉给药的I期研究概述。 货币。 操作。 摩尔-热。 2003; 5(6):618–624. [ 公共医学 ] [ 谷歌学者 ] -
Martinez-Sobrido L.、Gitiban N.、Fernandez-Sesma A.、Cros J.、Mertz S.E.、Jewell N.A.、Hammond S.、Flano E.、Durbin R.K.、Garcia-Sastre A.和Durbin J.E.通过重组新城疫病毒载体对抗呼吸道合胞病毒。 J.维罗尔。 2006; 80(3):1130–1139. doi:10.1128/JVI.80.3.1130-1139.2006。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
McGinnes L.W.,Morrison T.G.单个半胱氨酸残基在新城疫病毒成熟抗原HN蛋白形成中的作用。 病毒学。 1994; 200(2):470–483. doi:10.1006/viro.1994.1210。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Mebatsion T.、Koolen M.J.、de Vaan L.T.、de Haas N.、Braber M.、Romer Oberdorfer A.、van den Elzen P.、van der Marel P.新城疫病毒(NDV)标记疫苗:NDV核蛋白基因上的免疫显性表位可以被缺失或被外源表位取代。 J.维罗尔。 2002; 76(20):10138–10146. doi:10.1128/JVI.76.2010138-1046.2002。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Mebatsion T.、Weiland F.、Conzelmann K.K.狂犬病病毒基质蛋白负责子弹状颗粒的组装和出芽,并与跨膜尖峰糖蛋白G.J.病毒相互作用。 1999; 73(1):242–250. doi:10.1128/jvi.73.1.242-250.1999。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Melanson V.R.,Iorio R.M.纽卡斯尔病病毒HN蛋白茎中F特异性结构域的氨基酸替换调节融合并干扰其与F蛋白的相互作用。 J.维罗尔。 2004; 78(23):13053–13061. doi:10.1128/JVI.78.23.13053-13061.2004。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Mirza A.M.,Iorio R.M.纽卡斯尔病病毒血凝素-神经氨酸酶(HN)蛋白的柄部发生突变,尽管可以有效地形成HN–F复合物,但可以阻止F蛋白的触发。 J.维罗尔。 2013; 87(15):8813–8815. doi:10.1128/JVI.01066-13。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Molouki A.,Yusoff K.在缺乏Bax的情况下NDV诱导细胞凋亡; 线粒体上游凋亡蛋白参与的证据。 2012年7月; 9:179. doi:10.1186/1743-422X-9-179。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Morrison T.G.副粘病毒融合蛋白的结构和功能。 生物芯片。 生物物理学。 《学报》。 2003; 1614(1):73–84. doi:10.1016/s0005-2736(03)00164-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Nagai Y.,Klenk H.D.,Rott R.病毒糖蛋白的蛋白质裂解及其对新城疫病毒毒力的意义。 病毒学。 1976; 72(2):494–508. doi:10.1016/0042-6822(76)90178-1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Nakaya T.、Cros J.、Park M.S.、Nakayay Y.、Zheng H.、Sagrera A.、Villar E.、Garcia-Sastre A.、Palese P.重组新城疫病毒作为疫苗载体。 J.维罗尔。 2001; 75(23):11868–11873. doi:10.1128/JVI.75.23.11868-11873.2001。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Nakaya Y.,Nakaya.T.,Park M.S.,Cros J.,Imanishi J.,Palese P.,Garcia-Sastre A.通过两个重组负链RNA病毒载体诱导对猴免疫缺陷病毒gag的细胞免疫反应。 J.维罗尔。 2004; 78(17):9366–9375. doi:10.128/JVI.78.17.9366-9375.2004。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Nayak B.、Route S.N.、Kumar S.、Khalil M.S.、Fouda M.M.、Ahmed L.E.、Earhart K.C.、Perez D.R.、Collins P.L.、Samal S.K.对表达H5血凝素的新城疫病毒的鸡进行免疫可保护其免受高致病性H5N1禽流感病毒的侵袭。 《公共科学图书馆·综合》。 2009; 4(8):e6509。 doi:10.1371/journal.pone.0006509。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Ochi Y.,Hashimoto K.政府机构兽医。 研究(精选); 1929.韩国Uber eine neue Geflugelseuche。 第六次报告。 8月20日,16页。 [ 谷歌学者 ] -
Panda A.,Huang Z.,Elankumaran S.,Rockemann D.D.,Samal S.K.融合蛋白切割位点在新城疫病毒毒力中的作用。 微量。 Pathog公司。 2004; 36(1):1–10. doi:10.1016/j.micpath.2003.07.003。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Pantua H.D.、McGinnes L.W.、Peeples M.E.、Morrison T.G.关于装配和释放新城疫类病毒颗粒的要求。 J.维罗尔。 2006; 80(22):11062–11073. doi:10.1128/JVI.00726-06。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Park M.S.、Garcia-Sastre A.、Cros J.F.、Basler C.F.、Palese P.纽卡斯尔病病毒V蛋白是宿主范围限制的决定因素。 J.维罗尔。 2003; 77(17):9522–9532. doi:10.1128/JVI.77.17.9522-9532.2003。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Park M.S.、Steel J.、Garcia-Sastre A.、Swayne D.、Palese P.具有双重特异性的工程病毒疫苗构建物:禽流感和纽卡斯尔病。 程序。 国家。 阿卡德。 科学。 美国2006年; 103(21):8203–8208. doi:10.1073/pnas.0602566103。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
鸡新城疫病毒基质蛋白的Peeples M.E.和Bratt M.A.突变可导致融合糖蛋白融入颗粒的减少,并降低感染性。 J.维罗尔。 1984; 51(1):81–90. doi:10.1128/jvi.51.1.81-90.1984。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Peeters B.P.、de Leeuw O.S.、Koch G.、Gielkens A.L.从克隆的cDNA中拯救新城疫病毒:融合蛋白的可裂解性是毒力的主要决定因素的证据。 J.维罗尔。 1999; 73(6):5001–5009. doi:10.1128/jvi.73.6.5001-5009.1999。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Peeters B.P.、de Leeuw O.S.、Verstegen I.、Koch G.、Gielkens A.L.。重组嵌合新城疫病毒疫苗的产生,可在接种动物和感染动物之间进行血清学区分。 疫苗。 2001; 19(13-14):1616–1627. doi:10.1016/s0264-410x(00)00419-9。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Pelet T.、Delenda C.、Gubbay O.、Garcin D.、Kolakofsky D.仙台病毒复制启动子的部分特征和六法则。 病毒学。 1996; 224(2):405–414. doi:10.1006/viro.1996.0547。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Puhler F.、Willuda J.、Puhlmann J.、Mumberg D.、Romer-Oberdorfer A.、Beier R.。重组溶瘤性新城疫病毒的产生和两个转基因的完整IgG抗体的表达。 基因疗法。 2008; 15(5):371–383. doi:10.1038/sj.gt.3303095。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ravindra P.V.、Tiwari A.K.、Ratta B.、Chaturvedi U.、Palia S.K.、Subudhi P.K.、Kumar R.、Sharma B.、Rai A.、Chauhan R.S.新城疫病毒诱导Vero细胞凋亡需要病毒复制、从头蛋白质合成和胱天蛋白酶激活。 病毒研究2008; 133(2):285–290. doi:10.1016/j.virusres.2008.01.010。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ravindra P.V.、Tiwari A.K.、Sharma B.、Chauhan R.S.新城疫病毒作为溶瘤剂。 2009年印度医学研究杂志; 130(5):507–513. [ 公共医学 ] [ 谷歌学者 ] -
Ravindra P.V.、Tiwari A.K.、Sharma B.、Rajawat Y.S.、Ratta B.、Palia S.、Sundaresan N.R.、Chaturvedi U.、Kumar G.B.、Chindera K.、Saxena M.、Subudhi P.K.、Rai A.、Chauhan R.S.新城疫病毒HN蛋白导致鸡胚成纤维细胞凋亡。 架构(architecture)。 维罗尔。 2008; 153(4):749–754. doi:10.1007/s00705-008-0057-2。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Reichard K.W.、Lorence R.M.、Cascino C.J.、Peeples M.E.、Walter R.J.、Fernando M.B.、Reyes H.M.、Greager J.A.纽卡斯尔病病毒选择性杀死人类肿瘤细胞。 《外科学杂志》,1992年; 52(5):448–453. doi:10.1016/0022-4804(92)90310-v。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Rodier E.菲律宾禽病。 程序。 Soc.Exp.生物。 医学,1928年; 25:781–783. [ 谷歌学者 ] -
Romer-Oberdorfer A.、Mundt E.、Mebatsion T.、Buchholz U.J.、Mettenleiter T.C.从cDNA中生成重组致透镜性新城疫病毒。 《病毒遗传学杂志》。 1999; 80(第11部分):2987–2995。 doi:10.1099/0022-1317-80-11-2987。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Romer-Oberdorfer A.、Veits J.、Helferich D.、Mettenleiter T.C.以新城疫病毒为基础的减毒活载体疫苗对鸡的高致病性H5禽流感病毒保护水平取决于疫苗和挑战病毒之间H5序列的同源性。 疫苗。 2008; 26(19):2307–2313. doi:10.1016/j.vaccine.2008.02.061。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Romer-Oberdorfer A.、Werner O.、Veits J.、Mebatsion T.、Mettenleiter T.C.HN蛋白长度和F蛋白裂解位点序列对新城疫病毒致病性的贡献。 《病毒遗传学杂志》。 2003; 84(第11部分):3121–3129。 doi:10.1099/vir.0.19416-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Route S.N.,Samal S.K.。大聚合酶蛋白与新城疫病毒的毒力有关。 J.维罗尔。 2008; 82(16):7828–7836. doi:10.1128/JVI.00578-08。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Sakaguchi T.、Fujii Y.、Kiyotani K.、Yoshida T.副粘病毒F蛋白前体蛋白水解裂解与哺乳动物细胞中毛皮、PACE4和PC6基因表达的相关性。 《病毒遗传学杂志》。 1994; 75(第10部分):2821–2827。 doi:10.1099/0022-1317-75-10-2821。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Samal S.、Khattar S.K.、Kumar S.、Collins P.L.、Samal S.K.从新城疫病毒融合F蛋白的七个重复序列中协同删除N-聚糖,产生一种复制性、毒力和免疫原性增强的高融合性病毒。 J.维罗尔。 2012; 86(5):2501–2511。 doi:10.1128/JVI.06380-11。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Samal S.、Khattar S.K.、Paldurai A.、Palaniyandi S.、Zhu X.、Collins P.L.、Samal S.K.在新城疫病毒融合蛋白的细胞质域中的突变赋予了调节病毒复制和致病性的高融合表型。 J.维罗尔。 2013; 87(18):10083–10093. doi:10.128/JVI.01446-13。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Samal S.、Kumar S.、Khattar S.K.、Samal S.K.纽卡斯尔病病毒融合蛋白裂解位点序列中的单一氨基酸改变Q114R可减弱病毒复制和致病性。 《病毒遗传学杂志》。 2011; 92(第10部分):2333–2338。 doi:10.1099/vir.0.033399-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Samal S.K.In:新城疫和相关禽副粘病毒。 第一卷副粘病毒生物学。 Samal S.K.,编辑。 凯斯特学术出版社; 英国诺福克:2011年。 [ 谷歌学者 ] -
Sanchez-Felipe L.,Villar E.,Munoz Barroso I.新城疫病毒进入宿主细胞:酸性pH和内吞作用。 生物芯片。 生物物理学。 《学报》。 2014年; 1838(1):300–309. doi:10.1016/j.bbamem.2013.08.008。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Schirrmacher V.,Griesbach A.,Ahlert T.纽卡斯尔病病毒体内抗肿瘤作用:局部与全身效应。 国际期刊Oncol。 2001; 18(5):945–952. doi:10.3892/ijo.18.5.945。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Schroer D.、Veits J.、Grund C.、Dauber M.、Keil G.、Granzow H.、Mettenleiter T.C.、Romer-Oberdorfer A.接种新城疫病毒载体疫苗可保护鸡免受高致病性H7禽流感病毒的感染。 禽疾病。 2009; 53(2):190–197. doi:10.1637/8416-072308-Reg.1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Seal B.S.、King D.J.、Meinersmann R.J.新城疫病毒基质蛋白基因的分子进化和副粘病毒科之间的系统发育关系。 病毒研究2000; 66(1):1–11. doi:10.1016/s0168-1702(99)00119-7。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Sergel T.、McGinnes L.W.、Morrison T.G.NDV HN蛋白的融合促进活性与神经氨酸酶活性无关。 病毒学。 1993; 196(2):831–834. doi:10.1006/viro.1993.1541。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Sergel T.、McGinnes L.W.、Peeples M.E.、Morrison T.G.纽卡斯尔病病毒血凝素-神经氨酸酶蛋白的附着功能可以通过突变从促进融合中分离出来。 病毒学。 1993; 193(2):717–726. doi:10.1006/viro.1993.1180。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Sharma B.、Pokhriyal M.、Rai G.K.、Saxena M.、Ratta B.、Chaurasia M.、Yadav B.S.、Sen A.、Mondal B.从非禽类宿主(绵羊)中分离新城疫病毒及其影响。 架构(architecture)。 维罗尔。 2012; 157(8):1565–1567. doi:10.1007/s00705-012-1317-8。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Sheehan J.P.,Iorio R.M.纽卡斯尔病病毒血凝素-神经氨酸酶中的单一氨基酸替换导致两种功能中的蛋白质均缺失。 病毒学。 1992; 189(2):778–781. doi:10.1016/0042-6822(92)90605-o。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Shobana R.、Samal S.K.、Elankumaran S.前列腺特异性抗原再靶向重组新城疫病毒用于前列腺癌病毒治疗。 J.维罗尔。 2013; 87(7):3792–3800。 doi:10.1128/JVI.02394-12。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Snoeck C.J.、Owoade A.A.、Couacy-Hymann E.、Alkali B.R.、Okwen M.P.、Adeyanju A.T.、Komoyo G.F.、Nakoune E.、Le Faou A.、Muller C.P.西非和中非家禽新城疫病毒的高遗传多样性:基因型XIV和新定义基因型XVII和XVIII的共循环。 临床杂志。 微生物。 2013; 51(7):2250–2260. doi:10.1128/JCM.00684-13。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Steward M.、Vipond I.B.、Millar N.S.、Emmerson P.T.《纽卡斯尔病病毒的RNA编辑》。 《病毒遗传学杂志》。 1993; 74(第12部分):2539–2547。 doi:10.1099/0022-1317-74-12-2539。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Stojdl D.F.、Lichty B.、Knowles S.、Marius R.、Atkins H.、Sonenberg N.、Bell J.C.利用以前未知的溶瘤病毒利用干扰素途径中的肿瘤特异性缺陷。 《自然医学》,2000年; 6(7):821–825. doi:10.1038/77558。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Stone-Hulslander J.、Morrison T.G.《新城疫病毒感染细胞中HN和F蛋白相互作用的检测》。 J.维罗尔。 1997; 71(9):6287–6295. doi:10.1128/jvi.71.9.6287-6295.1997。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Stone-Hulslander J.,Morrison T.G.纽卡斯尔病病毒HN蛋白膜近端区域七肽重复序列的突变分析。 J.维罗尔。 1999; 73(5):3630–3637. doi:10.1128/jvi.73.5.3630-3637.1999。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Swayne D.E.、King D.J.禽流感和纽卡斯尔病。 美国兽医杂志。 2003年医学协会; 222(11):1534–1540. doi:10.2460/javma.2003.222.1534。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ten R.M.、Blank V.、Le Bail O.、Kourilsky P.、Israel A.两个因子,IRF1和KBF1/NF-kappa B,在干扰素α-β或新城疫病毒诱导MHC I类基因表达的过程中相互配合。 C.R.学院。 科学。 三、 1993年; 316(5):496–501. [ 公共医学 ] [ 谷歌学者 ] -
Tolf C.、Wille M.、Haidar A.K.、Avril A.、Zohari S.、Waldenstrom J.波罗的海西部地区秋季迁徙期间Mallards 1型禽副粘病毒的流行情况。 维罗尔。 2013年7月; 10(1):285. doi:10.1186/1743-422X-10-285。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Tong S.,Compans R.W.同型和异型副流感病毒HN和F蛋白之间相互作用的替代机制。 《病毒遗传学杂志》。 1999; 80(第1部分):107–115。 doi:10.1099/0022-1317-80-1-107。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
丰田章男、坂口章男、Imai K.、Inocencio N.M.、Gotoh B.、Hamaguchi M.、Nagai Y.新城疫病毒强毒株和弱毒株之间融合糖蛋白裂解活化位点的结构比较。 病毒学。 1987; 158(1):242–247. doi:10.1016/0042-6822(87)90261-3。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Vigil A.、Park M.S.、Martinez O.、Chua M.A.、Xiao S.、Cros J.F.、Martinez-Sobrido L.、Woo S.L.、Garcia-Sastre A.使用反向遗传学增强新城疫病毒的溶瘤特性。 癌症研究2007; 67(17):8285–8292. doi:10.1158/008-5472.CAN-07-1025。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
韦斯特伯里H.纽卡斯尔病病毒:一种进化中的病原体? 鸟类病理学。 2001; 30(1):5–11. doi:10.1080/03079450020023131。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
谢忠、谢磊、陈安、刘杰、庞毅、邓欣、范奇。一种从野生白鹭中分离的新城疫病毒的全基因组序列分析。 J.维罗尔。 2012; 86(24):13854–13855. doi:10.1128/JVI.02669-12。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Yan Y.,Rout S.N.,Kim S.H.,Samal S.K.血凝素-神经氨酸酶基因非翻译区在新城疫病毒复制和致病性中的作用。 J.维罗尔。 2009; 83(11):5943–5946. doi:10.128/JVI.00188-09。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Zamarin D.、Martinez-Sobrido L.、Kelly K.、Mansour M.、Sheng G.、Vigil A.、Garcia-Sastre A.、Palese P.、Fong Y.通过对抗细胞固有免疫反应增强重组新城疫病毒的溶瘤特性。 摩尔-热。 2009; 17(4):697–706. doi:10.1038/mt.2008.286。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
张志勇,王力强,傅春芳,李霞,崔志乐,张建勇,薛S.H.,孙宁,徐凤。基因靶向治疗与HN和VP3基因重组鸡痘病毒联合治疗小鼠骨肉瘤。 欧洲药理学评论。 科学。 2013; 17(6):767–776. [ 公共医学 ] [ 谷歌学者 ] -
赵伟,胡宏,扎克·L,于强,杨振华HN基因C-末端延伸不是新城疫病毒肠道向性的决定因素。 病毒基因。 2013; 47(1):27–33. doi:10.1007/s11262-013-0903-5。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]