缩写
ANS公司 自主神经系统
集成电路 心脏内的
ICNS公司 心脏内神经系统
IVC公司 下腔静脉
LCN公司 局部电路神经元
低压 左心室/心室
医疗保险 心肌梗死
RV汽车 右心室/心室
贵宾 血管活性肠肽
VIV糖蛋白 腹侧室间神经节丛
及物动词 室性快速心律失常
介绍 由于室性心律失常导致的心脏性猝死是世界上主要的死亡原因之一,估计每年导致400万至500万人死亡(Zipes&Wellens, 1998 ; 楚 等 . 2008 ). 心肌梗死(MI)后自主神经系统(ANS)的失调在心律失常的发生和心力衰竭的进展中起着至关重要的作用(Vaseghi和Shivkumar, 2008 ; Shen和Zipes, 2014 ; 福田 等 . 2015 ). 心脏神经轴负责心脏电和机械功能的动态调节(Ardell, 2004 ; 盔甲, 2004 ),涉及心脏层面的神经网络(Armour, 2008 )到岛叶皮层(奥本海默和霍普金斯, 1994 ; 灰色 等 . 2007 ).
在器官水平上,内在心脏神经系统(ICNS)由神经节和互连神经(Armour、, 2008 ). ICNS与较高的神经轴突中枢(胸内心外神经节、脊髓、脑干和皮层)共同调节心脏的兴奋性和收缩功能(Ardell, 2004 ; 盔甲, 2004 ; 博蒙特 等 . 2013 ). ICNS包含独立于高级中枢的心内反射控制所必需的所有神经元元件(墨菲 等 . 2000 )即感觉神经元、肾上腺素能和胆碱能传出节后神经元以及插入的局部回路神经元(LCN)(Armour, 2008 ; 博蒙特 等 . 2013 ). 最大的亚群LCN负责ICNS内神经元之间的节内和节间通信,并负责局部信息处理(Armour, 2008 ; 博蒙特 等 . 2013 ).
心脏病,如心肌梗死,对心肌及其相关神经成分产生不利影响(Vracko 等 . 1991 ; 曹 等 . 2000 ; 肯伯 等 . 2013 ; 阿吉约拉 等 . 2015 ). 心脏传入神经将传递心脏损伤的神经信号传递到多个水平的心脏神经轴(Armour, 1999 ). 心肌梗死后心神经轴的重塑及其对感觉信号的处理(Wang 等 . 2014 )有助于神经体液激活(扎克 等 . 2012 )以及心脏性猝死的可能性(福田 等 . 2015 ). 患有缺血性心脏病的人的内源性心脏(IC)神经元含有内含物和液泡,树突和轴突发生退行性变化(霍普金斯 等 . 2000 ). 体外 对慢性心肌梗死动物IC神经元的细胞内研究表明,兴奋性增强,突触效能改变,神经化学表型和神经调节的适应性改变(哈德威克 等 . 2014 ). 然而,关于这种变化对神经信号传导的功能后果,目前知之甚少 体内 在治愈梗死的情况下。
本研究的目的是(1)检查ICNS内的形态和表型重塑,(2)直接评估传入和传出(交感和副交感)输入的ICNS处理的重塑,以及心肌梗死后LCN对其整合。
在本研究中,我们发现心肌梗死会导致某些IC神经节内的形态学和神经化学变化。 这种结构重塑与ICNS处理传入和传出神经信号过程中的功能改变以及整体功能网络连通性或神经元对独立刺激对作出反应的能力下降平行。 传入神经信号的异质性,以及会聚神经元的重塑,可能在心律失常的发生和心力衰竭的进展中发挥重要作用。 缺血性心脏病中这种不良神经信号的特征化有可能作为疾病进展的标志,并可能用于监测和评估针对ANS的治疗。
方法 道德认可 心脏正常的约克郡猪( n个 = 16; 8名男性和8名女性; 49±3 kg)和约克郡猪治愈的根尖前心肌梗死( n个 = 16; 6名男性和10名女性; 46±2 kg)。 所有动物实验均按照美国国立卫生研究院 实验动物护理和使用指南 加州大学洛杉矶分校校长动物研究委员会批准。
心肌梗死的形成 如上所述诱发MI(Nakahara 等 . 2011 ). 简单地说,用特拉唑(8 mg kg)给动物镇静剂 −1 , i.m.), 插管和通气。 全身麻醉包括异氟醚(1-2%,吸入)。 监测12导联心电图和动脉压。 获得左股动脉通路,并在透视下将导丝(0.035英寸Amplatz Super Stiff guidewire with J‐Tip;Boston Scientific,Marlborough,MA,USA)放入左主冠状动脉。 然后将3mm血管成形术球囊导管(FoxCross PTA导管;美国加利福尼亚州特梅库拉Abbot Vascular)穿过导丝,在冠状动脉左前降支发出的大约第三条斜冠状动脉处充气。 球囊充气30秒后,将含有1 ml聚苯乙烯微球(Polybead,直径90μm;Polysciences Inc.,Warrington,PA,USA)的5 ml生理盐水悬浮液通过导管的中央管腔远端注入动脉。 造影血管造影显示动脉闭塞,肢体和心前导联ST段抬高证实急性心肌梗死。
心肌梗死后实验方案 在心肌梗死后42±2天对治愈的心肌梗死动物进行研究。 MI和年龄匹配的对照动物用特拉唑(8 mg kg)镇静 −1 , i.m.), 插管和通气。 全身麻醉用异氟醚维持(1-2%,吸入)。 麻醉深度通过血液动力学指标、下颌张力和踏板收回反射监测; 必要时调整麻醉。 获得右股静脉通路用于维持液体管理,获得右股动脉通路用于监测动脉压力。 胸骨正中切开术暴露心脏、星状神经节、下腔静脉(IVC)和降主动脉。 对颈部进行横向切开,露出颈部迷走神经和颈动脉。 在血管(下腔静脉、主动脉和颈动脉)周围放置Snare封堵器,在(迷走神经)周围或(星状神经节)自主传出神经结构内放置刺激电极。 手术完成后,全身麻醉改为α-氯醛糖(50 mg kg −1 静脉注射。 10 mg kg的丸剂 −1 小时 −1 连续的 静脉注射。 输液)。 通过加热垫监测和维持体温。 每小时评估一次酸碱状态; 调整呼吸频率和潮气量,必要时注入碳酸氢盐以维持血气平衡。 实验结束时,动物被过量的戊巴比妥钠(100 mg/kg)杀死 −1 , 静脉注射。 )其次是氯化钾(150 mg kg −1 , 静脉注射。 )来阻止心脏跳动。
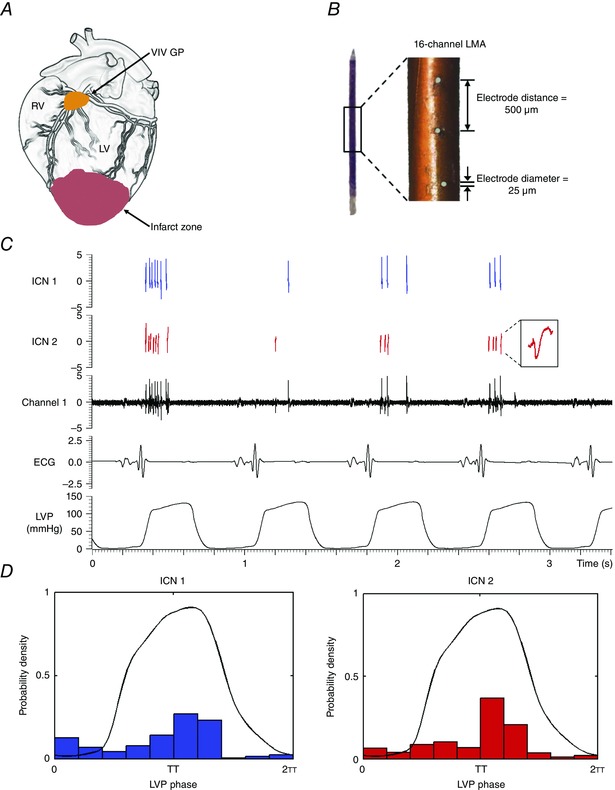
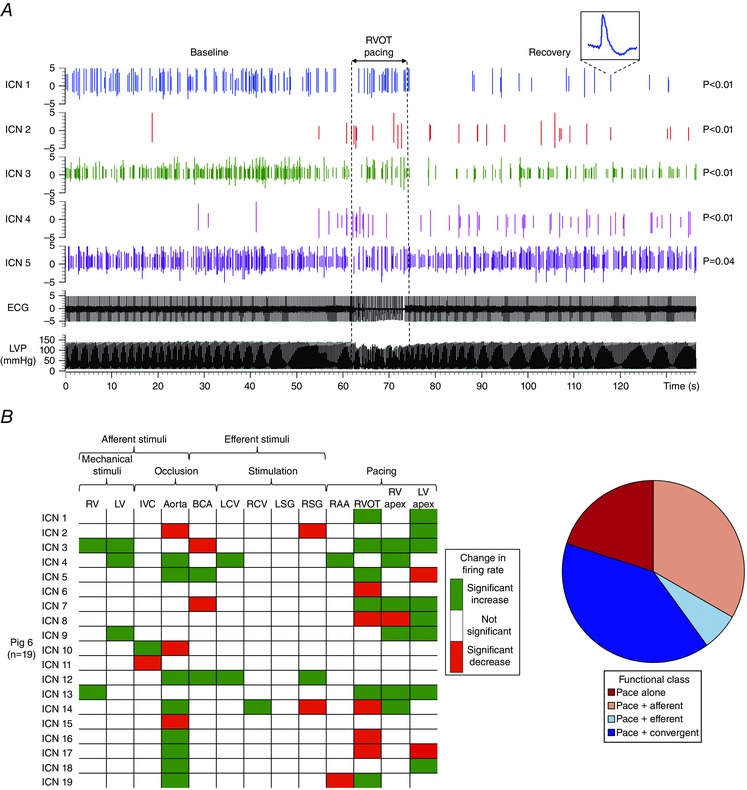
记录心脏固有神经元活动 使用线性微电极阵列(MicroProbes,Gaithersburg,MD,USA)记录 体内 腹侧室间神经节丛(VIV-GP)神经元产生的活动(图。 1 一个 ). 线性微电极阵列由16个铂-铱电极组成(直径25μm的电极,暴露尖端为2 mm;1 kHz时阻抗为0.3–0.5 mΩ)(图。 1 B类 ). 电极被嵌入VIV-GP中,该电极位于左冠状动脉主干的左前降支起点附近(图。 1 一个 )(阿罗拉 等 . 2003 ). 线性微电极阵列连接到柔性电缆上,从而使其能够半浮动。 电极线以及接地电极和参比电极与带有前置放大器的16通道微电极放大器相连(3600型;a‐M Systems Inc.,华盛顿州卡尔斯堡,美国)。 对于每个通道,滤波器设置为300 Hz至3 kHz,增益为5000。 将电极缝合到右心房心肌上,以提供参考右心房电图。 将神经波形、心电图、右心房电图和血流动力学数据输入数据采集系统(Power1401;英国剑桥电子设计公司)。 如前所述,使用Spike2(剑桥电子设计)软件离线分析数据(Beaumont 等 . 2013 ).
图1。 方法:心脏内神经元记录 .
一个 ,示意图显示了腹侧室间神经节丛(VIV-GP)的位置,从中记录了神经元活动。 B类 ,用于记录的16通道线性微电极阵列(LMA) 体内 VIV GP中包含的多个单个神经元的活动。 C类 ,显示从LMA的单个电极(通道1)识别的两个固有心脏(IC)神经元(ICN 1和ICN 2)的活动的代表性迹线。 ICN2插件是已识别神经元的扩展版本。 D类 ,面板神经元的基本活动 C类 与心脏周期有关。 注意,这两个神经元的活动主要集中在收缩期。 右心室; 左心室; 心电图; 左心室压。
左心室血流动力学评估 通过左股动脉将压力导管(Mikro‐Tip;Millar Instruments,美国德克萨斯州休斯顿)放入左心室(LV)室,并连接至控制单元(PCU‐2000;Millar-Instruments)。 通过收缩末期压力和最大压力变化率(d)评估左室收缩功能 P(P) /d日 t吨 最大值)。 通过舒张末期压力和最小室压变化率(d)评估左室舒张功能 P(P) /d日 t吨 最小值)。
传入神经输入评估 为了确定IC神经元传递机械感觉传入输入的能力,在以下四个部位施加心外膜机械刺激(轻触)10 s:右心室流出道、右心室心尖、左心室中前壁和左心室心尖。 然后使用圈套器封堵器对IVC和主动脉进行短暂(30s)闭塞,以确定神经元分别转导前负荷和后负荷的急性变化的能力。
传出神经输入评估 为了确定哪些IC神经元接受副交感和交感传出输入,将双极螺旋袖套电极(PerenniaFlex 304型;Cyberonics Inc.,美国德克萨斯州休斯顿)放置在颈迷走神经周围,并将双极针电极插入两侧星状神经节。 使用带有光电隔离装置的刺激器(S88和PSIU6;Grass Technologies,Warwick,RI,USA)调节IC神经元的传出输入。 对于每条迷走神经,阈值被定义为引起心率或血压下降10%所需的电流(20 Hz频率,1 ms脉宽)。 对于每个星状神经节,阈值被定义为引起心率或血压增加10%所需的电流(4 Hz频率,4 ms脉宽)。 然后以阈值电流和1 Hz的频率分别刺激每条迷走神经和星状神经节1 min。 这样做是为了评估ICNS的直接输入,而不考虑心脏功能的任何变化。 然后使用圈套封堵器短暂(1分钟)封堵双侧颈动脉(颈动脉窦尾侧),以确定颈动脉压力反射调节IC神经元输出输入的能力。
心外膜起搏 为了确定IC神经元对心脏电刺激的反应能力,将双极起搏电极(St Jude,St Paul,MN,USA)放置在不同的心外膜部位,并以高于基线心率10%的速度进行起搏(6 mA电流;2 ms脉冲宽度),共10次捕获的心跳。 对以下四个部位进行起搏:(1)右心耳,(2)右室流出道,(3)右室心尖,(4)左室心尖。
心室快速心律失常诱导性 在一组单独的控件中( n个 =8)和治愈的根尖前心肌梗死动物( n个 =8),通过程序化心室刺激(EPS320;Micropace,Canterbury,New South Wales,Australia),在两个不同的周期长度(600和400 ms),以及来自两个不同部位(RV心尖和LV前壁心外膜)的最多三个额外刺激(最短200 ms),评估室性快速心律失常(VT)的诱导性。
组织加工 IC神经元记录完成后,立即处死动物并切除心脏。 取出含有VIV GP、室间背神经节丛、右边缘动脉神经节丛和右心房神经节丛的脂肪垫,在冷(4°C)盐水中冲洗,并转移到10%的磷酸盐缓冲福尔马林(Fisher Scientific,Pittsburgh,PA,USA)中4天。 然后,将组织转移到70%乙醇中(美国密苏里州圣路易斯Sigma‐Aldrich),并在3天内将石蜡包埋。 从石蜡块上切下4μm厚的切片。
组织学染色 使用计算机形态分析(Aperio ImageScope;Leica Biosystems,Buffalo Grove,IL,USA),从苏木精和伊红染色切片(类别号H345‐25;Fisher Scientific,Pittsburgh,PA,USA。
免疫组织化学染色 通过酪氨酸羟化酶免疫反应定量IC神经元肾上腺素能表型(1:2000稀释;目录号ab112;美国马萨诸塞州剑桥市Abcam); 胆碱乙酰转移酶免疫反应的神经元胆碱能表型(1:200稀释;目录号AB144-P;Millipore,Billerica,MA,USA); 和抗VIP抗体的血管活性肠肽(VIP)免疫反应(分类号20077;ImmunoStar,Hudson,WI,USA)。 使用Dako EnVision+System–HRP标记的聚合物抗兔抗体(目录号K4003;Dako North America Inc.,Carpinetria,CA,USA)在1:500时对酪氨酸羟化酶和VIP进行二次检测,并使用多克隆兔抗羊免疫球蛋白/生物素化物(E0466,Dako)检测胆碱乙酰转移酶。 二氨基联苯胺(美国纽约州格兰德岛Life Technologies)根据制造商推荐的所有染色剂的方案检测到次要免疫反应。 然后对幻灯片进行数字扫描,并对电子图像进行分析。 使用计算机图像分析(Aperio ImageScope)在20–40倍放大率下对载玻片中的所有神经元进行量化。 以盲法对各组进行染色和量化。
数据分析:心脏多单位固有神经元活动的信号处理 如前所述,使用离线分析进行伪影消除和IC神经元识别(图。 1 C类 )(博蒙特 等 . 2013 ). 简言之,记录的神经元活动受到邻近心房和心室心肌活动产生的内源性电伪影以及刺激自主传出神经产生的外源性电伪像的污染。 在线性微电极阵列的三个以上相邻通道中同时出现的显示类似波形的活动也被认为是伪影。 识别后,通过消隐或将包含伪影的时间间隔的振幅设置为零,从所有通道中去除伪影。 该过程导致总信号的最大损失为3%。 去除伪影后,使用波形的主成分分析(Beaumont 等 . 2013 ).
数据分析:监测单个心脏固有神经元的活动 对于心外膜机械刺激和自主传出神经刺激,在刺激前1分钟比较IC神经元活性(基线) 与 刺激期间。 对于血管闭塞和起搏,比较基线时的神经元活动 与 刺激期间以及基线时 与 刺激后1分钟(恢复)。 每次刺激后,我们至少等待5分钟,让神经元活动和血流动力学恢复到基线水平,然后再继续。 根据IC神经元对心血管刺激的反应特征,将其功能分为传入、传出或会聚(图。 2 B类 和 C类 ). 传入神经元被定义为仅对心外膜机械刺激和/或下腔静脉或主动脉闭塞作出反应的神经元。 传出神经元被定义为仅对自主传出神经(迷走神经或星状神经节)刺激和/或双侧颈动脉闭塞作出反应的神经元。 对传入和传出输入的激活作出反应的神经元被定义为会聚(博蒙特 等 . 2013 ).
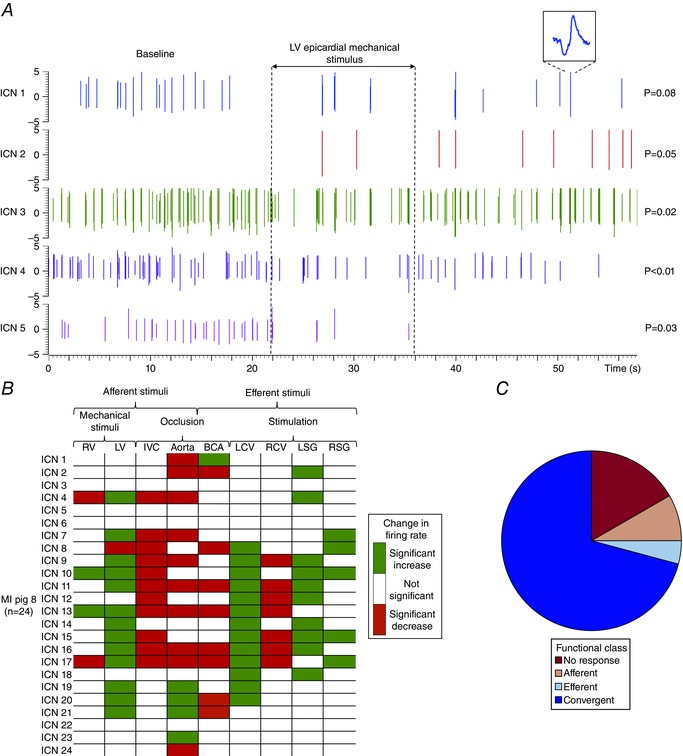
图2。 心脏固有神经元的分析和功能分类 .
一个 ,在一个对照心脏的5个IC神经元中记录到尖峰活动。 垂直虚线表示左室心外膜机械刺激的开始和偏移。 ICN 1插件是已识别神经元的扩展版本。 注意,与基线相比,神经元亚群表现出增加(ICN 2)、减少(ICN 3、4和5)或活动无变化(ICN 1)。 在每条迹线的右侧显示了诱发活动变化的显著性水平。 P(P) 值是根据方法中描述的分析得出的。 B类 ,总结了心肌梗死(MI)动物对心血管刺激的反应中神经元活动的诱发变化。 水平行表示单个神经元对给定刺激的响应(垂直列)。 绿色表示活动显著增加( P(P) < 0.05); 红色表示显著下降( P(P) < 0.05). C类 ,图中所示神经元的功能分类 B类 根据神经元对心血管刺激的反应,将其分为传入、传出或会聚。 传入神经元被定义为仅对RV或LV的心外膜机械刺激作出反应的神经元; 下腔静脉暂时闭塞; 和/或降主动脉暂时闭塞。 传出神经元被定义为只对电刺激左侧(LCV)或右侧颈迷走神经(RCV)作出反应的神经元; 电刺激左侧(LSG)或右侧星状神经节(RSG); 和/或双侧颈动脉(BCA)暂时闭塞。 对传入和传出输入的激活作出反应的神经元被定义为会聚神经元。
数据分析:条件概率 条件概率分析用于确定对一个刺激作出反应的IC神经元是否也对另一个刺激做出反应,如前所述(Beaumont 等 . 2013 ). 刺激之间潜在的功能关系 X(X) 和刺激 Y(Y) 在每只动物的神经元内被量化为神经元对刺激作出反应的条件概率 Y(Y) 对刺激也有反应 X(X) .条件概率(概率:对 Y(Y) |对…的响应 X(X) )被估计为对两种刺激都有反应的神经元数量 X(X) 和刺激 Y(Y) 除以对刺激作出反应的神经元数量 X(X) .
统计 基线之间每个IC神经元放电率变化的显著性水平 与 使用基于Skellam分布(Shin 等 . 2010 ). 这项测试之前已经在IC神经元的研究中得到验证(博蒙特 等 . 2013 ). Aχ 2 试验用于比较心肌梗死患者的神经元反应和VT诱导率 与 .控制动物。 Wilcoxon的签名等级测试或Mann-Whitney U型 该测试用于比较心肌梗死患者神经元的放电频率、静息血流动力学指标以及神经元的形态和表型变化 与 .控制动物。 数据表示为平均值±平均值标准误差。 P(P) ≤0.05被认为具有统计学意义。 使用SigmaPlot 12.0(Systat Software Inc.,San Jose,CA,USA)进行统计分析。
结果 治愈的心尖前心肌梗死动物处于慢性代偿状态,没有明显的心力衰竭,类似的静息血流动力学指标如左室舒张末压(3±1 与 .4±1毫米汞柱( P(P) =0.59)和LV d P(P) /d日 t吨 最大值(1436±112 与 0.1426±151毫米汞柱秒 −1 ( P(P) =1.00)(单位:MI) 与 .分别控制动物。 图 三 J型 和 K(K) 说明了微栓塞技术导致瘢痕形成的典型模式。 由于心室颤动是一种终末事件,因此在另一组动物中评估了该模型中心室颤动的诱导性。 而没有任何对照动物( n个 =8)可诱导,75%的MI动物( n个 =8)开发的VT( P(P) <0.01)。
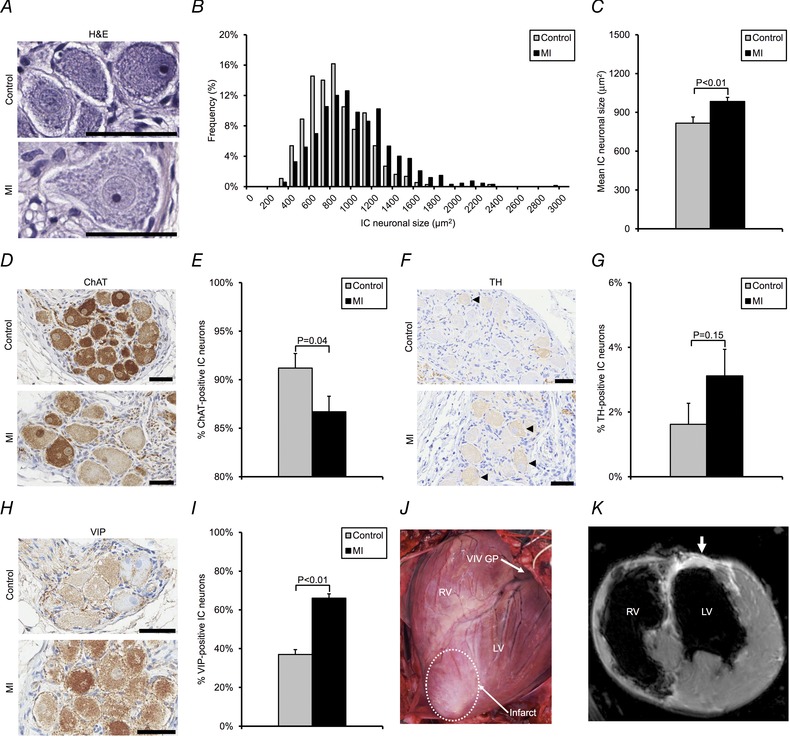
图3。 心肌梗死诱导心脏固有神经元的形态和表型重塑 .
一个 ,显示对照组VIV GP苏木精和伊红(H&E)染色神经元的显微照片 与 .MI动物。 B类 ,对照组心脏内(IC)神经元大小分布直方图 与 .MI动物。 C类 ,对照组平均神经元大小 与 .MI动物。 D类 ,显微照片显示对照组VIV-GP用胆碱乙酰转移酶(ChAT)染色 与 .MI动物。 ChAT催化乙酰胆碱的合成,并被用于鉴定可能的胆碱能神经元。 E类 ,对照组ChAT阳性神经元的百分比 与 .MI动物。 F类 ,显微照片显示对照组VIV-GP用酪氨酸羟化酶(TH)染色 与 .MI动物。 TH催化去甲肾上腺素合成中的限速步骤,并用于识别假定的肾上腺素能神经元(黑色箭头)。 G公司 ,对照组TH阳性神经元的百分比 与 .MI动物。 H(H) ,显微照片显示对照组VIV-GP被血管活性肠肽(VIP)染色 与 .MI动物。 VIP是心脏功能的调节剂和假定传入神经元的标记物。 我 ,对照组和心肌梗死动物中VIP阳性神经元的百分比。 J型 图为一个患有治愈的根尖前心肌梗死的猪心脏。图示VIV GP相对于梗死疤痕(白色虚线)的位置。 K(K) ,对应心脏的短轴心脏磁共振图像。 白色箭头表示疤痕组织导致的延迟性高强化区域。 中的数据 C类 , E类 , G公司 和 我 以平均值±平均值的标准误差(SEM)表示。 曼·惠特尼 U型 测试用于 C类 , E类 , G公司 和 我 确定各组之间的重要性。 比例尺 一个 , D类 , F类 和 H(H) 代表50μm。
心肌梗死引起不同程度的心脏内生神经元增大和表型改变 对VIV-GP神经元的大小、肾上腺素能-胆碱能表型和VIP表型进行组织学和免疫组织化学分析,以评估心肌梗死引起的潜在形态学和神经化学变化。VIP是心脏功能的调节剂和假定的传入标记物(Weihe和Reinecke, 1981 ; 布鲁姆 等 . 1986 ; 弗拉塞 等 . 1987 ; 莱特尔 等 . 2015 ).
心肌梗死动物的VIV GP神经元明显大于对照组(946±23 与 .755±22微米 2 分别为; P(P) <0.01)(图。 三 A–C ). MI和对照动物的神经元大小分布直方图如图所示。 三 B类 VIV-GP、背侧室间神经节丛和右侧边缘动脉神经节丛中观察到神经元扩大,它们对心室有优先影响,但在右心房神经节丛没有观察到,它们对心房有优先影响(图。 4 A–C ).
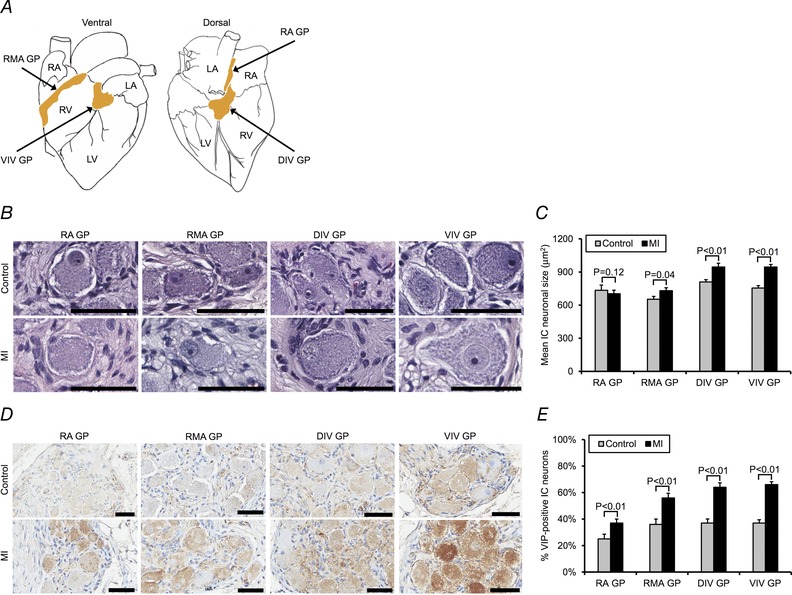
图4。 心肌梗死诱导心脏内神经元的不同形态和神经化学重塑 .
一个 ,示意图显示了右心房神经节丛(RA GP)、右边缘动脉神经节丛、背侧室间神经节丛和VIV GP的位置。RA GP和RMA GP分别优先影响右心房和RV, 而DIV GP和VIV GP对LV具有优先影响。 B类 ,显示对照组研究的神经节丛(GPs)H&E染色神经元的显微照片 与 .MI动物。 C类 ,对照组GP中IC神经元的平均大小 与 .MI动物。 D类 ,照片显示GPs被VIP染色 与 .MI动物。 E类 ,对照组GP中VIP阳性神经元的百分比 与 .MI动物。 中的数据 C类 和 E类 表示为平均值±SEM。 曼·惠特尼 U型 测试用于 C类 和 E类 确定各组之间的重要性。 比例尺英寸 B类 和 D类 代表50μm。 RA,右心房; 左心房。
与对照组相比,MI动物表达胆碱乙酰转移酶的IC神经元百分比显著降低(87±2% 与 分别为.91±2%; P(P) =0.04)(图。 三 D类 和 E类 ). 相反,与对照动物相比,心肌梗死中酪氨酸羟化酶的表达没有显著差异(3±1% 与 分别为.2±1%; P(P) =0.15)(图。 三 F类 和 G公司 ). 心肌梗死患者VIP表达显著增加 与 .对照动物(66±2% 与 分别为.37±3%; P(P) <0.01)(图。 三 H(H) 和 我 ). VIP在所研究的所有其他神经节丛(GPs)中的表达也显著增加(图。 4 D类 和 E类 ).
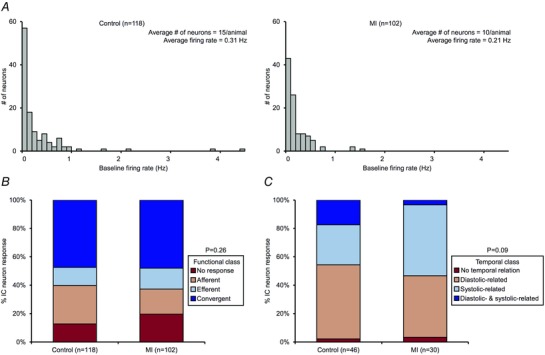
心肌梗死后心脏固有神经元的功能特征 这个 体内 使用微电极阵列在对照和MI动物中记录来自VIV GP的神经元的活动,以评估MI诱导的神经元反应特征的功能变化(图 1 和 2 ). 在8只对照动物中,研究了来自VIV GP的118个IC神经元产生的活动(平均:每只动物15±3个神经元)(图。 5 一个 ,左侧面板)。 在8只MI动物中,研究了102个神经元产生的活动(平均:每只动物10±2个神经元)(图。 5 一个 ,右侧面板)。 神经元的自发放电率来自于基线间隔的汇集数据。 对照组动物神经元的平均自发放电频率为0.31Hz(范围:0-4.42Hz),而心肌梗死组动物的平均自发发射频率为0.21Hz(区间:0-1.59Hz)。 两种状态下的分布总体相似,90%以上的神经元在1Hz以下放电。
图5。 心肌梗死不会引起心脏内神经元功能或时间特征的整体变化 .
一个 ,对照组IC神经元基线放电率直方图 与 .MI心脏。 B类 ,对照神经元的功能分类 与 .MI心脏。 C类 对照组神经元的心脏周期相关周期 与 .MI心脏。 注意,神经元亚群表现出舒张相关活动、收缩相关活动、舒张和收缩相关活动或随机行为。 心肌梗死没有显著改变神经元的功能或时间特征。 Aχ 2 测试用于 B类 和 C类 确定各组之间的重要性。
根据其对心血管刺激的反应特征,IC神经元在功能上分为传入、传出或会聚(图2 B类 和 C类 、和5 B类 ). 在对照组和心肌梗死动物中,会聚神经元代表最大的亚群(47% 与 分别为.48%),其次是传入较少(27% 与 分别为.18%和13% 与 .15%)神经元(图。 5 B类 ). 对照组动物13%的神经元和心肌梗死动物20%的神经元对任何刺激都没有反应。 这两个州的总体分类都没有显著差异( P(P) = 0.26).
将IC神经元的活动与心脏周期进行比较,以确定它们是否表现出与心脏周期相关的周期性(图 1 D类 和 5 C类 ). 根据活动直方图,如果在基线时产生至少100个动作电位的神经元有30%以上的活动发生在给定的阶段,则将其归类为与心脏周期的特定阶段相关。 对照动物中的46个神经元(39%)和MI动物中的30个神经元(29%)满足这一标准,对其进行了心动周期相关周期性分析(图 5 C类 ). 在对照组动物中,52%的神经元表现出舒张相关活动,28%表现出收缩相关活动,17%表现出舒张和收缩双重相关活动,2%表现出随机行为。 相反,在心肌梗死动物中,43%的神经元表现出舒张相关活动,50%表现出收缩相关活动,3%表现出舒张和收缩双重相关活动,以及3%表现出随机行为。
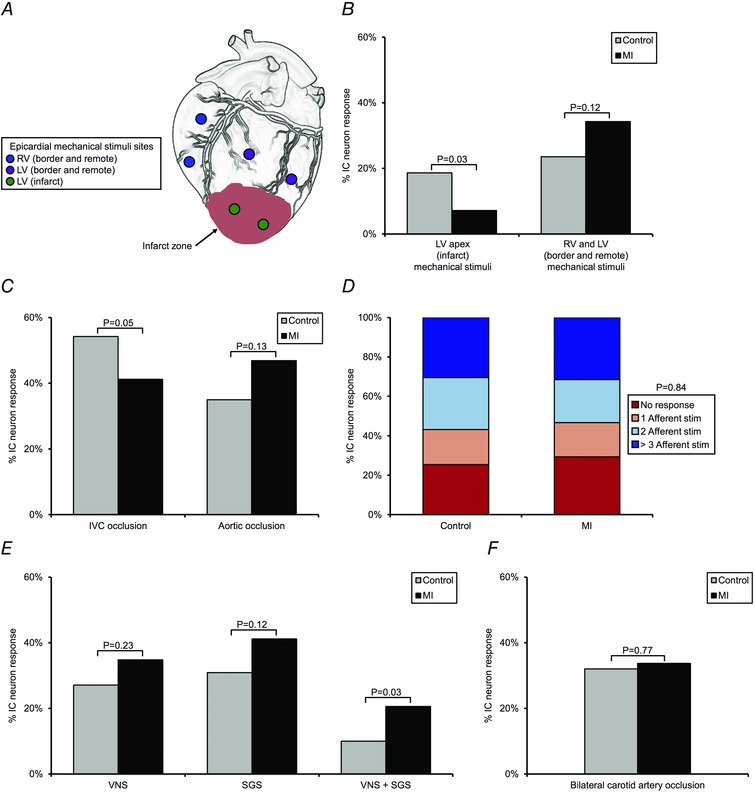
心肌梗死后心脏固有神经元的传入重塑 心肌梗死不同程度地影响了IC神经元传递梗死引起的机械敏感性传入信号的能力 与 心脏的边缘和边远区域,通过对这些区域上的心肌组织施加机械刺激进行评估(图。 6 一个 和 B类 ). 心肌梗死动物对边缘区和边远区(右心室流出道、右心室顶点和左心室中前壁)机械敏感性输入激活的神经元反应与对照组相似(心肌梗死组为34% 与 .24%为对照组; P(P) = 0.12). 相反,对MI后梗死(LV心尖)引起的机械敏感性输入激活有反应的神经元明显较少(MI为7% 与 .19%在对照组; P(P) = 0.03).
图6。 心肌梗死诱导心脏固有神经元的传入和传出重塑 .
一个 心外膜机械刺激的部位,用于评估IC神经元传递来自RV或LV的机械敏感性传入输入的能力。 B类 ,接受来自LV心尖(梗死)的机械敏感性输入的神经元百分比 与 RV和LV(边境和偏远地区)处于受控状态 与 .MI心脏。 C类 ,对照组对短暂下腔静脉阻塞或主动脉闭塞反应的神经元百分比 与 .MI心脏。 IVC和主动脉闭塞分别用于评估神经元传递前负荷和后负荷变化的能力。 D类 ,控制中传递多个传入输入的神经元百分比 与 .MI心脏。 E类 ,分别通过颈迷走神经刺激(VNS)和星状神经节刺激(SGS)评估对照组接受副交感和/或交感神经系统传出输入的神经元百分比 与 .MI心脏。 F类 ,对照组对短暂双侧颈动脉闭塞反应的神经元百分比 与 .MI心脏。 颈动脉闭塞用于评估压力反射调节神经元传出输入的能力。 Aχ 2 测试用于 B类 , C类 , D类 , E类 和 F类 确定各组之间的重要性。
心肌梗死后,IC神经元传递心脏负荷变化的能力也受到影响(图。 6 C类 ). 在心肌梗死动物中,短暂下腔静脉闭塞引起的预负荷减少引起的神经元反应显著减弱(心肌梗死组为41% 与 对照组为.54%; P(P) = 0.05). 对降主动脉短暂部分闭塞引起的后负荷增加的神经元反应没有显著差异(MI为47% 与 .35%为对照组; P(P) = 0.13). MI同样也没有显著改变神经元传递多模态传入神经信号的整体能力(图。 6 D类 ; 两种状态下接受一个、两个或三个以上传入输入的神经元比例相似( P(P) = 0.84)). 这些传入输入包括心外膜机械刺激和短暂闭塞下腔静脉和主动脉。
心肌梗死后心脏固有神经元的传出重构 通过分别对颈迷走神经和星状神经节的低频刺激评估,心肌梗死并不影响IC神经元单独传递副交感神经和交感神经传出输入的能力(图。 6 E类 ). 以低频率刺激这些自主传出神经,以评估对ICNS的直接传出输入,而不是由心功能改变引起的间接反应。 与对照组相比,MI动物接受左侧或右侧颈迷走神经输入的神经元百分比没有显著差异(35% 与 分别为.27%; P(P) = 0.23). 从左或右星状神经节接收输入的神经元百分比也观察到类似的模式(MI中为41% 与 .31%在对照组; P(P) = 0.12). 有趣的是,与对照组相比,MI动物中接收ANS交感和副交感神经分支传出输入的神经元百分比显著增加(21% 与 分别为.10%; P(P) = 0.03). 在接受交感神经和副交感神经输入的神经元中,90%的神经元对一个或多个传入输入的刺激也有反应,这两个神经元分别是对照组(11个神经元中的10个)和MI组(21个神经元中有19个)。 因此,这些神经元被归类为会聚神经元。
为了评估MI对IC神经元传出输入的压力反射调制的影响,双侧颈动脉在颈动脉窦尾侧闭塞(图。 6 F类 ). 在两种状态下,对颈动脉闭塞反应的神经元百分比没有显著差异(心肌梗死时为34% 与 .32%为对照组; P(P) = 0.77).
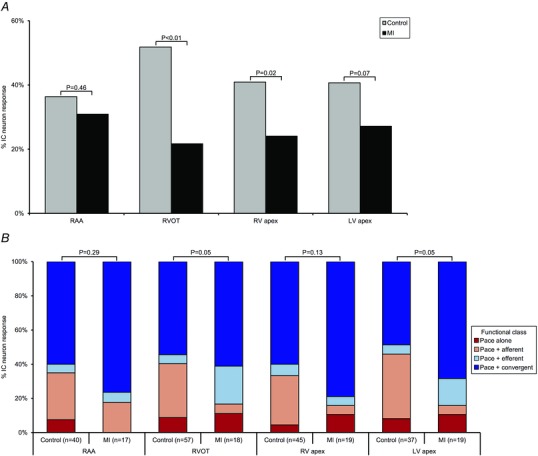
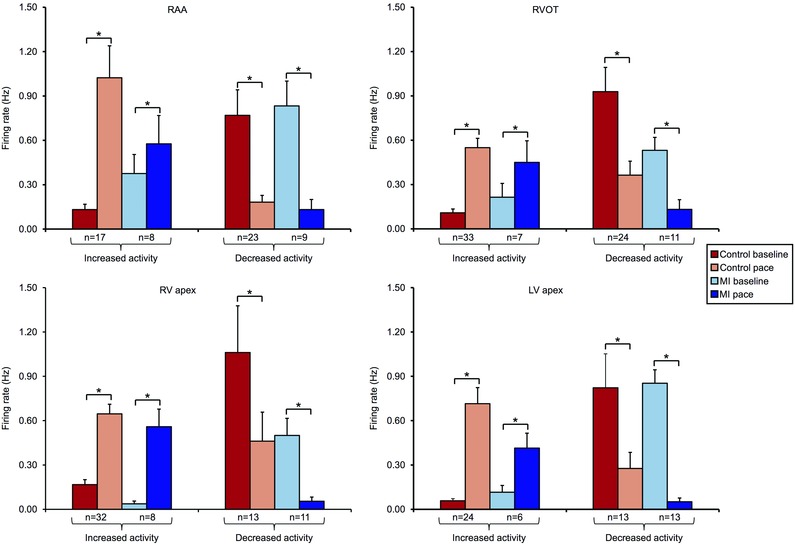
心肌梗死引起心脏内神经元对起搏反应的改变 心肌梗死不同程度地影响了IC神经元对心外膜起搏的反应(图 7 一个 和 8 一个 ). 然而,右心耳起搏(心室捕捉)的神经元反应没有改变(心肌梗死31% 与 对照组为.36%; P(P) =0.46),心室起搏反应降低(心肌梗死44% 与 对照组为.63%; P(P) < 0.01). 对起搏有反应的神经元在功能上分为传入、传出或会聚(图7 B类 和 C类 、和8 B类 ). 右心室流出道(远端)和左室心尖部(梗死)起搏诱发的神经元反应在心肌梗死后受到最显著的影响( P(P) = 0.05). 这种改变主要反映为起搏反应性会聚神经元的上调和起搏反应传入神经元的相应下调。 同样值得注意的是,在对照组动物和心肌梗死动物中评估的所有部位中,起搏参与了一个独特的神经元亚群,它只对起搏反应,而对其他传入或传出刺激没有反应(图。 8 B类 ,红色条)。
图7。 心脏固有神经元对起搏反应的分析和功能分类 .
一个 ,在对照动物的5个IC神经元中记录到尖峰活动。 垂直虚线表示右心室流出道(RVOT)心外膜起搏的开始和偏移。 ICN1插图是一个已识别神经元的扩展版本。 请注意,神经元的亚群与基线相比活动增加、减少或无变化。 轨迹右侧显示了每个神经元活动诱导变化的显著性水平。 P(P) 根据方法中所述的分析得出值。 B类 ,总结了心肌梗死动物局部心外膜起搏引起的神经元活动的诱发变化,以及对其他心血管刺激的反应。 绿色表示活动显著增加( P(P) < 0.05); 红色表示显著下降( P(P) < 0.05). C类 ,面板中描述的速度反应神经元的功能分类 B类 使用图。 2 .RAA,右心耳。
图8。 心肌梗死改变心脏固有神经元对起搏的反应和特征 .
一个 ,对照组中RAA、RVOT、RV心尖或LV心外膜起搏反应的IC神经元百分比 与 .MI心脏。 心肌梗死导致心室神经元反应的差异性降低 与 .心房起搏。 B类 ,控制中速度反应神经元的功能分类 与 .MI动物。 心肌梗死改变了梗死区(LV心尖)和边远区(RVOT)起搏的反应特征。 Aχ 2 测试用于确定组间的显著性。
心脏固有神经元的状态依赖性:对诱发反应的影响 基础活动影响IC神经元对随后心血管刺激的反应,包括起搏(图。 9 ). 在对照组和心肌梗死动物中,基础活动低的神经元往往通过起搏激活( P(P) < 0.01). 相反,基础活性高的神经元往往受到起搏的抑制( P(P) < 0.01). 这些结果表明了这些神经元的状态依赖性。
图9。 心脏固有神经元的状态依赖性 .
对照组RAA、RVOT、RV心尖部或LV心外膜起搏时基线和反应时的IC神经元活性 与 .MI动物。 神经元根据诱发的增加进行细分 与 .起搏后活动减少。 基础活性低的神经元被起搏激活,而基础活性高的神经元被抑制,这表明其具有状态依赖性。 数据以平均值±SEM表示。 Wilcoxon的符号秩检验用于确定组间的显著性* P(P) < 0.01.
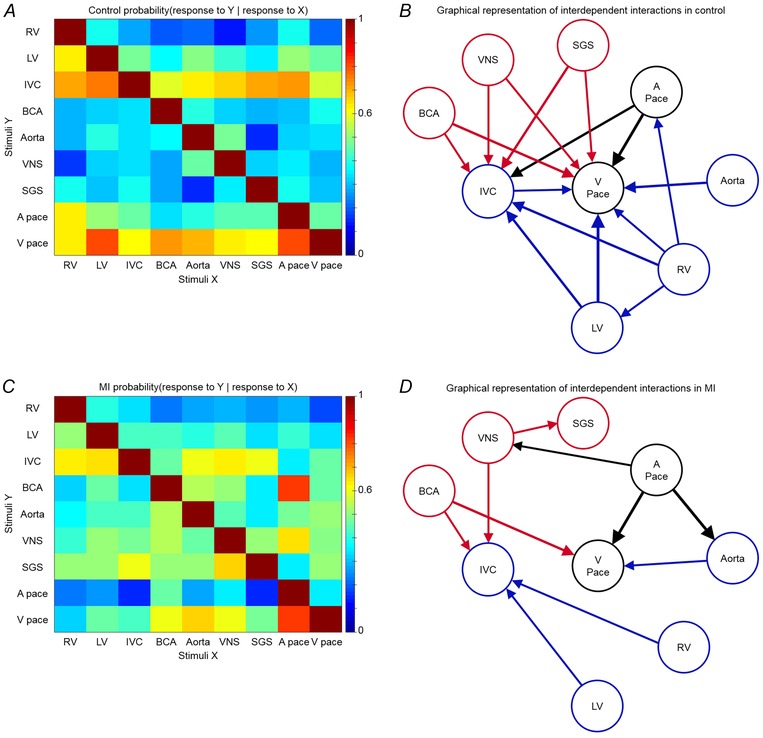
心肌梗死后心脏内神经元对刺激的相互依赖性 在对照和MI动物中确定了IC神经元对传入和传出刺激的反应以及起搏之间的关系。 对一个刺激作出反应的神经元是否也对另一个刺激做出反应的条件概率以矩阵形式表示(图。 10 一个 和 C类 ). 这些数据也以图形方式描述为网络,仅显示条件概率≥0.6的链路(图。 10 B类 和 D类 ). 这些关系反映了由成对独立刺激诱导的VIV-GP神经元群体之间的一致行为。 MI降低了ICNS内的整体功能网络连接。
图10。 心肌梗死降低了心脏固有神经系统内的功能网络连通性 .
一个 ,IC神经元响应一个刺激的条件概率( X(X) , x个 轴)也对另一刺激作出反应( Y(Y) , 年 ‐轴)。 B类 ,对照动物中刺激物之间相互依赖的相互作用的图形表示。 C类 ,一个神经元对一个刺激作出反应的条件概率( X(X) , x个 轴)也对另一刺激作出反应( Y(Y) , 年 心梗动物。 D类 ,心肌梗死动物刺激之间相互依赖的相互作用的图形表示。 色标输入 一个 和 C类 表示每次发生的概率级别。 箭头厚度in B类 和 D类 与条件概率的强度成正比。 仅显示概率≥0.6的链接。 传入和传出刺激分别用蓝色和红色表示。 心房(A步)和心室起搏(V步)用黑色表示。
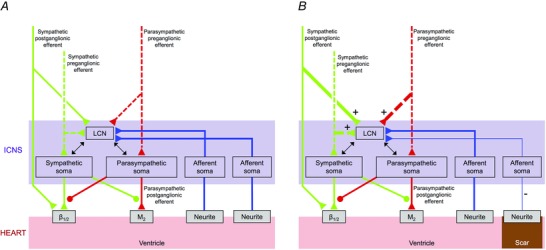
讨论 本研究以结构和 体内 心肌梗死后ICNS内神经元元件的功能重塑,代表了慢性梗死心脏的第一个“心脏神经电图”。 在VIV GP中评估ICNS的网络功能,该神经丛主要与心室功能的控制有关(Cardinal 等 . 2009 ). 这项研究有几个主要发现。 首先,心肌梗死后IC神经节经历形态和表型重塑。 损伤部位决定了哪一个神经节重塑。 其次,梗死区至IC神经元的传入神经信号减弱,而来自边缘和边远区的信号在MI后保留下来,从而形成“神经感觉边界区”,或损伤传入信息的异质性 与 相邻的非损伤心肌组织(图。 11 ). 传入神经信号的改变还表现为IC神经元传递前负荷变化的能力降低。 第三,心肌梗死后维持ICNS的自主传出输入(图。 11 ). 第四,收敛的IC LCN,即既接受传入输入又接受传出输入的IC-LCN,在心肌梗死后具有增强的转导能力(图。 11 ). 第五,在MI后,ICNS内的功能网络连通性降低。 最后,MI降低了对心室起搏的反应并改变了IC神经元的特性。
图11。 心肌梗死后心内神经系统的功能重塑 .
一个 ,示意图,显示了内在心脏神经系统(ICNS)和心脏之间的神经连接,以及来自健康心脏神经轴高级中心的输入。 B类 示意图显示了心肌梗死后ICNS与心脏之间的神经连接发生的变化。与心脏的边缘和边远区域相比,收敛性局部回路神经元(LCN)的交感和副交感输入增加,而梗死的传入输入减少。 绿色和红色虚线和连续线分别代表节前和节后纤维。
对照组和心肌梗死组动物之间的唯一区别是存在梗死疤痕。 因此,我们注意到的结构和功能变化可能归因于心肌梗死。我们研究了心肌梗死发生6周后ICNS的重塑。这是自主适应的一个稳定阶段,超出了以心肌细胞死亡和神经退行性变为特征的急性期重塑(Hardwick 等 . 2014 ). 根据左室舒张末期压力和收缩力等血流动力学指标,这些动物处于慢性代偿状态,并且没有转变为明显的心力衰竭。 我们记录了VIV GP中的神经元活动,因为它主要参与心室功能的控制(Cardinal 等 . 2009 )其神经元胞体位于梗死区上游(Arora 等 . 2003 ). 因此,我们观察到的神经重塑并不是由于神经元的直接缺血性损伤。
我们评估了VT的诱导性,并证明该模型中的MI动物具有明显更高的诱导性。 由于心室颤动是一种终末事件,因此在另一组动物中评估了诱导性。 具体而言,复苏动物所需的心脏复律电击会破坏和/或移位我们的神经元记录界面,并妨碍我们对ICNS的详细描述。 此外,电击本身有可能改变随后的神经活动。 我们之前也证明了ICNS的异质药理学或电激活是致心律失常的(Armour 等 . 2005 ; 博蒙特 等 . 2013 )神经反应迟钝会降低心律失常的可能性(Richer 等 . 2008 ; 吉本斯 等 . 2012 ; 艾黛儿 等 . 2014 ). 综上所述,我们现在可以假设,心肌梗死后我们观察到的改变的神经信号可能与室性心律失常的发生有关。
心脏内神经系统神经元的不同形态和神经化学重塑 MI导致VIV GP内神经元的形态学改变。在对心室产生优先影响的GP中观察到IC神经元增大,而对心房产生优先影响(红雀 等 . 2009 ). 据报道,反应性损伤导致神经元肥大(Barr&Hamilton, 1948 ; 亨德里克森和迪宁, 1982 ; 吉纳 等 . 1998 )和慢性信号(周 等 . 2004 ). 然而,由于我们研究的GPs位于梗死的上游,因此后一种机制可能就是这种情况。 神经元大小的差异性增大表明,MI后传入和传出神经信号存在异质性,神经元直接参与对受影响最大的梗死区的控制。 此外,损伤部位决定了哪一个神经节重塑。 在我们的结构和功能数据中观察到的这种异质性,可能在心律失常发生中起着重要作用(陈 等 . 2014 ). 这些结构变化与缺血性心肌病的人类和动物模型中胸外神经节的结构变化相似(Zhou 等 . 2004 ; 阿吉约拉 等 . 2012 ; 汉族 等 . 2012 ; 阿吉约拉 等 . 2015 ).
心肌梗死导致VIV-GP内胆碱能表型的减少。胆碱能神经元的减少可能与心肌梗死后中枢介导的副交感神经退缩有关(Vanoli 等 . 1991 ; 比尔曼, 2006 ). 虽然我们没有评估肾上腺素能-胆碱能转分化,但有强有力的证据表明,这发生在疾病状态下的心神经轴的其他神经节内(金泽 等 . 2010 ; 福田 等 . 2015 ). 心肌梗死后所有GP中VIP的表达也增加(图。 4 D类 和 E类 ). VIP被认为是一种冠状血管扩张剂,具有变力和变时作用(Weihe和Reinecke, 1981 ; 布鲁姆 等 . 1986 ; 弗拉塞 等 . 1987 ). 心肌梗死后ICNS内VIP表达增加可能是一种代偿机制,通过血管扩张来维持心脏功能,从而改善冠状动脉血流量,以及增强肌力和计时性。 VIP也与伤害有关(Rytel 等 . 2015 ). VIP阳性神经元的增加也可能是心肌梗死后传入信号增强的结果。
梗死边缘区的神经感觉输入 虽然心肌梗死后传出神经信号的改变已被广泛记录,但很少注意来自受损和相邻非受损心肌组织的传入神经信号。 解剖和功能研究已确定IC神经节中的单极神经元和位于心房和心室组织(Armour, 2008 ). 这些传入神经元传递心脏的局部机械和化学环境(Armour, 2008 ). 每个GP中包含的传入神经元具有空间上不同的感受野(Waldmann 等 . 2006 ; 红衣主教 等 . 2009 )允许从广泛的心脏区域传递感觉信息。 我们发现VIV GP神经元位于左冠状动脉主干(Arora)的左前降支起源附近 等 . 2003 ),转导来自覆盖RV和LV的不同心脏区域的感觉输入。
心肌梗死后,继发于缺血的梗死区发生心肌坏死。 此外,能量底物的缺乏和活性氧等分子的积累触发了细胞内信号传递的级联过程,导致梗死边缘区心肌细胞的重塑(Sutton&Sharpe, 2000 ). 在心肌细胞重塑的同时,在心脏神经轴的多个水平上发生适应性和不适应性变化,包括ICNS(Armour, 1999 ). 在急性状态下,传递损伤心肌组织传入信号的IC神经元过度异常激活(Huang 等 . 1993 ; 盔甲, 1999 ). 我们的数据表明,在慢性状态下,梗死区向ICNS的传入信号减少,但并未完全消除,而来自边缘和偏远地区的传入信号则得以保留。 传入神经信号的异质性导致边界条件和类似于瘢痕形成引起的心肌边界区的“神经感觉边界区”。 感觉边界条件的重要性已经在其他神经回路中得到了体现,例如视觉系统,在视觉系统中,视网膜神经节细胞检测非均匀光场的能力增强(Tessier‐Lavigne, 2000 ). 我们假设心肌梗死诱导的ICNS传入输入的不对称性可能是ANS反射激活的基础,包括交感神经兴奋。 在这方面,应用树脂毒素(瞬时受体电位香草醛1的一种有效激动剂)可降低心肌梗死后的心脏传入伤害性信号,降低交感神经兴奋,并与保持心脏功能有关(Wang 等 . 2014 ). 这些数据指出了传入神经信号在心脏病进展中的根本重要性,以及它们作为控制疾病过程的治疗靶点的作用。
收敛的局部回路神经元:心脏内神经系统的信息处理 在本研究中,我们发现IC神经元的一个亚群,称为会聚LCN,同时接收传入和传出输入。 收敛的LCN与传入和传出神经元一起构成IC神经回路的基本组成部分(Armour, 2008 ; 博蒙特 等 . 2013 ). 在这个电路中,收敛的LCN集成并处理信息,即使在MI状态下,这些神经元也有大量的亚群,这表明保持了局部信息处理的能力。 虽然心肌梗死后神经元的总体功能分类(传入、传出和收敛)没有差异,但对心血管刺激的综合神经网络反应适应和/或重塑了。 这在神经元对预负荷变化和区域起搏的反应以及通过条件概率分析评估的功能性神经网络连接的改变中很明显。 虽然大多数对起搏有反应的神经元也被发现能感应心血管动态变化,但有一个独特的亚群只对起搏起反应。 ICNS对心肌梗死区和心脏远端起搏反应的改变表明,起搏可能会在心肌梗死施加的压力之上对ICNS施加额外压力。在临床环境中,心室起搏已被证明对心脏功能有不利影响,导致心室不同步(Tops 等 . 2009 ). 据报道,在有梗死疤痕的情况下起搏也会引起电暴(Roque 等 . 2014 ). 此外,它可以加速心力衰竭的进展并增加死亡率(威尔科夫 等 . 2002 ; 斯威尼 等 . 2003 ). 这些不良事件的潜在机制与神经内分泌系统的激活有关(张 等 . 2013 ). 令人感兴趣的是,ICNS活性的调节,以及MI诱导的传入神经信号的重塑,可能是起搏和神经内分泌系统激活之间的基本联系。
传出神经控制 ICNS被经典地视为副交感神经节前向心脏传出投射的简单中继站(兰利, 1921 ). 与此相反,并支持在犬模型中获得的数据(Beaumont 等 . 2013 )我们发现,大部分猪IC神经元接受交感或副交感神经系统的输入,以及复杂的心血管传入输入。 事实上,IC神经元的一个子集接收到传出输入(星状神经节和迷走神经的输入)的汇合,这意味着ICNS内发生了大量的交感-副交感相互作用(McGuirt 等 . 1997 ; 兰德尔 等 . 2003 ).
心肌梗死后传出神经信号的改变发生在心脏神经轴的多个水平(Vracko 等 . 1991 ; 装甲, 1999 ; 曹 等 . 2000 ; 周 等 . 2008 ; 阿吉约拉 等 . 2012 , 2013 , 2015 ; 汉族 等 . 2012 ; 哈德威克 等 . 2014 ). 在器官水平上,观察到梗死心肌的交感神经去神经支配和边界区的高神经支配(Vracko 等 . 1991 ; 曹 等 . 2000 ). 交感神经节(如星状神经节(Ajijola 等 . 2012 , 2015 ). 交感神经影响的增加伴随着中枢介导的副交感神经(瓦诺利 等 . 1991 ; 比尔曼, 2006 ). 尽管交感-迷走神经平衡发生了这些变化,但我们的数据表明,心肌梗死后ICNS的交感和副交感神经输入保持不变。 事实上,心肌梗死后接受收敛性传出输入的IC神经元的百分比增加了一倍。反过来,这些IC神经元中的绝大多数本质上是收敛性局部回路,这一事实证明,90%的IC神经元受到传入刺激的影响。 这种适应可能是为了维持外周神经网络的稳定性,以应对交感迷走神经失衡和梗死引起的不同传入输入所造成的不稳定影响 与 心脏的边缘和边远区域。
限制 全身麻醉可以抑制神经系统的活动。 为了减少吸入麻醉剂的影响,我们在完成手术准备后立即改用α-氯醛糖输注。 我们使用轻触作为机械刺激,以确定VIV GP内传入神经元的感觉场。虽然有记录和标记神经元的方法,但我们是从跳动的心脏记录神经元的,这需要使用浮动微电极阵列。 因此,我们专注于描述 体内 神经元的功能 事后(post-hoc) 结构变化评估。 我们研究了猪心脏(Arora)七个GP中只有一个发生的功能性神经重塑 等 . 2003 ); 然而,这种GP主要与心室功能的控制有关(Cardinal 等 . 2009 )我们的组织学数据支持,它很可能受到梗死的影响最大。 我们评估了心肌梗死后固定时间点ICNS的结构和功能变化。在心脏病的演变过程中,心神经轴发生了动态变化(Hardwick 等 . 2014 ; 福田 等 . 2015 )因此,随着疾病的进展,神经信号可能会显示出不同的特征。 最后,虽然我们证明了该模型中的心肌梗死动物具有更强的致心律失常潜能,但我们并没有在接受心室颤动诱导的相同动物中获得神经元记录,并试图描述心室颤动期间ICNS神经特征。 复苏动物所需的心律转复电击会破坏和/或移动我们的神经元记录界面,并排除ICNS的详细特征。
其他信息 作者贡献 P.S.R.、J.L.A.和K.S.设计了所有实验; P.S.R.进行了所有神经元记录实验和相关分析; K.N.和O.A.A.进行了所有免疫组织化学实验和相关分析; P.S.R.和M.V.进行了所有VT诱导性研究; 所有作者都为撰写这篇论文做出了贡献。 所有作者都已经批准了手稿的最终版本,并同意对作品的各个方面负责。所有被指定为作者的人都有资格获得作者资格,所有符合作者资格的人都会被列出。
基金 这项工作得到了美国国立卫生研究院(NIH)国家心脏、肺和血液研究所(NHLBI)拨款(R01HL071830至J.L.A.;R01HL084261至K.S.)的支持。 P.S.R.得到了NIH国家普通医学科学研究所拨款(2T32GM065823)、美国心脏协会拨款(15PRE22230011)和NIH NHLBI拨款(F31HL127974)的支持。 O.A.A.得到了NIH NHLBI拨款(K08HL125730)的支持。 M.V.得到了美国心脏协会的资助(11FTF7550004)。
转换视角 本研究提供了直接证据,证明在临床相关的大动物模型中,心肌梗死时心脏内神经系统(ICNS)的功能性神经特征发生改变,并强调了神经元记录在阐明神经适应和/或心脏病适应不良方面的实用性。 传入神经信号的异质性可能是自主神经系统反射激活的基础,从而影响心律失常的可能性(陈 等 . 2014 ; 福田 等 . 2015 )以及进展为心力衰竭(Zucker 等 . 2012 ). 作为缓解缺血性心脏病的一种新的治疗方法,应研究从病变心肌到ICNS、胸外神经节和心神经轴的更高中心的传入神经信号的调制。 这些发现还提高了“心脏神经电图”作为研究心脏生理学和病理生理学的一种方式的可能性,因为它提供了一种潜在的方法来监测疾病进展和干预效果。
工具书类
Ajijola OA、Wisco JJ、Lambert HW、Mahajan A、Stark E、Fishbein MC和Shivkumar K(2012)。 心肌病患者的心外神经重构。 圆形心律失常电生理 5, 1010–1116. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Ajijola OA、Yagishita D、Patel KJ、Vaseghi M、Zhou W、Yamakawa K、So E、Lux RL、Mahajan A和Shivkumar K(2013)。 局灶性心肌梗死诱导心脏交感神经支配的整体重塑:空间背景下的神经重塑。 美国生理学杂志心脏循环生理学 305,H1031–1040。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Ajijola OA、Yagishita D、Reddy NK、Yamakawa K、Vaseghi M、Downs AM、Hoover DB、Ardell JL和Shivkumar K(2015)。 空间靶向心肌梗死后星状神经节神经元的重塑:神经肽和形态学变化。 心脏节律 12, 1027–1035. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Ardell J(2004)。 《基础和临床神经心脏病学》(Basic and Clinical Neurocardiology)胸内神经对心脏功能的调节。 牛津大学出版社,纽约。 [ 谷歌学者 ]
Ardell JL、Cardinal R、Beaumont E、Vermeulen M、Smith FM&Armour JA(2014年)。 慢性脊髓刺激改变了抑制心房颤动的内在心脏突触效能。 自主神经科学 186, 38–44. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
装甲JA(1999)。 心肌缺血和心脏神经系统。 心血管研究 41, 41–54. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Armour JA(2004)。 健康和疾病中的心脏神经元层次。 美国生理学杂志Regul Integr Comp Physiol 287,R262-271。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Armour JA(2008)。 哺乳动物心脏“小脑”的潜在临床相关性。 实验生理学 93, 165–176. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Armour JA、Richer LP、PagéP、Vinet A、Kus T、Vermeulen M、Nadeau R和Cardinal R(2005)。 纵隔神经激活引起的犬房性快速心律失常的起源和药理反应。 自主神经科学 118, 68–78. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Arora RC、Waldmann M、Hopkins DA和Armour JA(2003年)。 猪心内神经节。 Anat Rec A发现分子细胞进化生物学 271, 249–258. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Barr ML和Hamilton JD(1948年)。 轴突反应期间脊髓运动神经元某些形态学变化的定量研究。 《计算机神经学杂志》 89, 93–121. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Beaumont E、Salavatian S、Southerland EM、Vinet A、Jacquemet V、Armour JA和Ardell JL(2013)。 犬内在心脏神经系统内的网络相互作用:对局部心脏功能反射控制的影响。 生理学杂志 591, 4515–4533. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Billman GE(2006)。 全面回顾和分析25年来在体犬心脏性猝死模型的数据:对未来抗心律失常药物开发的影响。 药物治疗学 111, 808–835. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Brum JM、Bove AA、Sufan Q、Reilly W&Go VL(1986年)。 血管活性肠肽在冠状动脉循环中的作用和定位:非肾上腺素能、非胆碱能冠状动脉调节的证据。 美国心脏病学会杂志 7, 406–413. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Cao JM、Fishbein MC、Han JB、Lai WW、Lae AC、Wu TJ、Czer L、Wolf PL、Denton TA、Shintaku IP、Chen PS和Chen LS(2000)。 心脏局部超神经支配与室性心律失常的关系。 循环 101, 1960–1969. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
红衣主教R、PagéP、Vermeulen M、Ardell JL和Armour JA(2009年)。 犬心脏神经节丛神经元对烟碱刺激的空间发散性心脏反应。 自主神经科学 145, 55–62. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Chen PS、Chen LS、Fishbein MC、Lin SF和Nattel S(2014)。 自主神经系统在心房颤动中的作用:病理生理学和治疗。 循环研究 114, 1500–1515. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Chugh SS、Reinier K、Teodorescu C、Evanado A、Kehr E、Al Samara M、Mariani R、Gunson K和Jui J(2008)。 心脏性猝死的流行病学:临床和研究意义。 心血管疾病进展 51, 213–228. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Frase LL、Gaffney FA、Lane LD、Buckey JC、Said SI、Blomqvist CG和Krejs GJ(1987)。 血管活性肠肽对健康受试者的心血管影响。 美国心脏病杂志 60, 1356–1361. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Fukuda K、Kanazawa H、Aizawa Y、Ardell JL和Shivkumar K(2015)。 心脏神经支配与心脏性猝死。 循环研究 116, 2005–2019. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Geuna S、Borrione P、Poncino A和Giacobini‐Robecchi MG(1998)。 支配再生蜥蜴尾巴的背根神经节神经元的形态学和形态计量学变化。 国际神经科学发展杂志 16, 85–95. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Gibbons DD、Southerland EM、Hoover DB、Beaumont E、Armour JA和Ardell JL(2012)。 神经调节靶向内在的心脏神经元,以减弱神经介导的心房心律失常。 美国生理学杂志Regul Integr Comp Physiol 302,R357-364。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Gray MA、Taggart P、Sutton PM、Groves D、Holdright DR、Bradbury D、Brull D&Critchley HD(2007年)。 反映心脏功能的皮层电位。 《美国科学院院刊》 104, 6818–6823. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Han S、Kobayashi K、Joung B、Piccirillo G、Maruyama M、Vinters HV、March K、Lin SF、Shen C、Fishbein MC、Chen PS和Chen LS(2012)。 心肌梗死后左星状神经节的电解剖重建。 美国心脏病学会杂志 59, 954–961. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Hardwick JC、Ryan SE、Beaumont E、Ardell JL和Southerland EM(2014)。 慢性心肌梗死诱导豚鼠心内神经丛的动态重塑。 自主神经科学 181, 4–12. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Hendrickson A&Dineen JT(1982)。 幼年猴纹状体皮质损伤后背外侧膝状体核神经元肥大。 神经科学快报 30, 217–222. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Hopkins DA、Macdonald SE、Murphy DA&Armour JA(2000)。 缺血人类心脏固有神经元的病理学。 Anat Rec公司 259, 424–436. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Huang MH、Ardell JL、Hanna BD、Wolf SG和Armour JA(1993)。 暂时性冠状动脉闭塞对犬心脏固有神经元活动的影响。 综合生理行为科学 28, 5–21. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Kanazawa H、Ieda M、Kimura K、Arai T、Kawaguchi‐Manabe H、Matsuhashi T、Endo J、Sano M、Kawakami T、Kimula T、Monkawa T、Hayashi M、Iwanami A、Okano H、Okada Y、Ishibashi‐Ueda H、Ogawa S和Fukuda K(2010)。 心力衰竭通过gp130信号细胞因子导致啮齿动物心脏交感神经胆碱能转分化。 临床研究杂志 120, 408–421. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Kember G、Armour JA和Zamir M(2013年)。 心脏的神经控制层次尚未进化到处理心肌缺血的程度。 基因组学杂志 45, 638–644. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
兰利JN(1921)。 英国剑桥W.Heffer《自主神经系统》。 [ 谷歌学者 ]
McGuirt AS、Schmacht DC和Ardell JL(1997)。 SA结副交感神经切除术后,维持控制心房率的自主相互作用。 美国生理学杂志 272,H2525–2533。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Murphy DA、Thompson GW、Ardell JL、McCraty R、Stevenson RS、Sangalang VE、Cardinal R、Wilkinson M、Craig S、Smith FM、Kingma JG&Armour JA(2000)。 心脏移植后会重新神经化。 胸外科年鉴 69, 1769–1781. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Nakahara S、Vaseghi M、Ramirez RJ、Fonseca CG、Lai CK、Finn JP、Mahajan A、Boyle NG和Shivkumar K(2011年)。 心肌瘢痕的特征:猪梗死模型中的电生理成像相关性。 心脏节律 8,1060–1067之间。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Oppenheimer S&Hopkins D(1994)。 心脏的延髓上神经元调节,《神经病学》,阿莫尔·J和阿德尔·J主编,第309-342页。 牛津大学出版社,纽约。 [ 谷歌学者 ]
Randall DC、Brown DR、McGuirt AS、Thompson GW、Armour JA和Ardell JL(2003)。 心脏固有神经系统内的相互作用有助于变时调节。 美国生理学杂志Regul Integr Comp Physiol 285兰特,1066–1075兰特。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Richer LP、Vinet A、Kus T、Cardinal R、Ardell JL和Armour JA(2008)。 α‐肾上腺素受体阻断可改善神经诱导的心房心律失常。 美国生理学杂志Regul Integr Comp Physiol 295,R1175–1180。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Roque C、Trevisi N、Silberbauer J、Olorize T、Mizuno H、Baratto F、Bisceglia C、Sora N、Marzi A、Radinovic A、Guaracini F、Vergara P、Sala S、Paglino G、Gulletta S、Mazzone P、Cireddu M、Maccabelli G和Della Bella P(2014)。 心脏再同步化治疗引起的电风暴是由心外膜瘢痕上的起搏决定的,可以通过导管消融成功控制。 圆形心律失常电生理 7, 1064–1069. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Rytel L,Palus K&Całka J(2015)。 猪结节神经节感觉神经元中PACAP与VIP、SP和CGRP的共表达。 Anat Histol胚胎 44, 86–91. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Shen MJ和Zipes DP(2014)。 自主神经系统在调节心律失常中的作用。 循环研究 114, 1004–1021. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Shin HC、Aggarwal V、Acharya S、Schieber MH和Thakor NV(2010年)。 使用基于Skellam的最大似然解码对手指运动进行神经解码。 IEEE Trans生物医学工程 57, 754–760. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Sutton MG和Sharpe N(2000)。 心肌梗死后左室重构:病理生理学和治疗。 循环 101, 2981–2988. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Sweeney MO、Hellkamp AS、Ellenbogen KA、Greenspon AJ、Freedman RA、Lee KL、Lamas GA和Investigators MST(2003)。 在窦性结功能障碍起搏器治疗的临床试验中,在基线QRS持续时间正常的患者中,心室起搏对心力衰竭和心房颤动的不利影响。 循环 107, 2932–2937. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Tessier‐Lavigne M(2000)。 视网膜的视觉处理《神经科学原理》,第4版,编辑:坎德尔ER,施瓦茨JH。& Jessell TM,第507-522页。 纽约州麦格劳-希尔。 [ 谷歌学者 ]
Tops LF,Schalij MJ&Bax JJ(2009)。 右心室心尖部起搏对心室功能的影响以及对治疗的不同步影响。 美国心脏病学会杂志 54, 764–776. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Vanoli E、De Ferrari GM、Stramba‐Badiale M、Hull SS、Foreman RD&Schwartz PJ(1991)。 清醒犬心肌梗死治愈后迷走神经刺激和猝死预防。 循环研究 68, 1471–1481. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Vaseghi M&Shivkumar K(2008)。 自主神经系统在心脏性猝死中的作用。 心血管疾病进展 50, 404–419. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Vracko R、Thorning D和Frederickson RG(1991年)。 人心肌瘢痕中的神经纤维。 人体病理学 22, 138–146. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Waldmann M、Thompson GW、Kember GC、Ardell JL和Armour JA(2006年)。 心房和心室固有心脏神经元的随机行为。 应用生理学杂志 101, 413–419. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Wang HJ、Wang W、Cornish KG、Rozanski GJ和Zucker IH(2014)。 在心力衰竭大鼠中,心脏交感传入神经失神经可减轻心脏重塑并改善心血管功能障碍。 高血压 64, 745–755. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Weihe E&Reinecke M(1981)。 哺乳动物窦房结的肽能神经支配:血管活性肠多肽、神经降压素、P物质 26, 283–288. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Wilkoff BL、Cook JR、Epstein AE、Greene HL、Hallstrom AP、Hsia H、Kutalek SP和Sharma A; 双腔和VVI植入式除颤器试验研究人员 (2002). 植入除颤器患者的双腔起搏或心室备用起搏:双腔和VVI植入式除颤器(DAVID)试验。 日本汽车制造商协会 288, 3115–3123. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Zhang YY、Wu DY、Fu NK、Lu FM和Xu J(2013)。 单导联心房起搏和双腔起搏模式下的神经内分泌和血流动力学变化。 国际医学研究杂志 41, 1057–1066. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Zhou S、Chen LS、Miyauchi Y、Miyaughi M、Kar S、Kangavari S、Fishbein MC、Sharifi B和Chen PS(2004)。 犬心肌梗死后心脏神经萌芽的机制。 循环研究 95,76–83。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Zhou S、Jung BC、Tan AY、Trang VQ、Gholmieh G、Han SW、Lin SF、Fishbein MC、Chen PS和Chen LS(2008)。 犬猝死模型中的自发性星状神经节神经活动和室性心律失常。 心脏节律 5, 131–139. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Zipes DP和Wellens HJ(1998)。 心脏性猝死。 循环 98, 2334–2351. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Zucker IH、Patel KP&Schultz HD(2012年)。 神经体液刺激。 心力衰竭临床 8, 87–99. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]