特发性肺纤维化/普通间质性肺炎(IPF/UIP)是特发性间质性肺病中最常见和最严重的形式。1-4从病因和自然史来看,关于该病的许多问题仍未解决,最近的对比意见引发了对其发病机制的热烈讨论。5-13IPF/UIP的炎症理论受到了挑战,并基于以下假设提出了新的模型:组织损伤后间充质和上皮肺成分之间的失调交流是纤维化和组织重塑不可逆过程的关键。7,9,10这种观点的改变似乎特别有趣,因为它可能为旨在对比成纤维细胞增殖和/或诱导成纤维细胞凋亡的新治疗方法提供了理论基础。7月14日,15日然而,成纤维细胞/肌成纤维细胞在IPF/UIP中的中心地位仍然存在争议和未经证实,并且对参与该疾病发病机制的分子机制知之甚少。关键的论据涉及早期损伤的上皮靶点,以及异常间充质/上皮相互作用的分子特征。在这方面,我们最近观察到,细支气管上皮细胞和支气管-肺泡连接是IPF/UIP中细胞损伤的相关靶点,而异常的细支气管增生(包括增生、化生、细支气管化和蜂窝状)实际上可能代表了该病的实质性特征。16
许多证据表明,对调节肺发育和形态发生的分子途径的研究可能提供有关肺部疾病发病机制的重要信息。8,16肺发育的早期阶段依赖于复杂的分子网络,包括一系列刺激和抑制途径,包括胎儿生长因子,埃格夫,转化生长因子β/激活素,Wnt(重量),刺猬,霍克斯,SOX公司,萌芽的,以及其他。17-22可以合理地假设,这些分子通路可以在出生后的生活中部分重演,因为复杂的肺结构可以在广泛损伤后精确地重建,正如在各种肺部疾病中观察到的那样。
在这项研究中,我们重点关注Wnt信号及其效应物β-catenin,原因有很多。首先,这种相关的信号通路参与哺乳动物的肺发育和器官发生。19-22其次,金属蛋白酶matrylisin/MMP-7是β-catenin反式激活的靶点,最近被发现是肺纤维化的关键调节因子。23-25第三,Wnt通路与一些人类纤维性疾病的发病机制有关。26,27最后,最近有直接证据表明β-catenin信号在上皮-间质转化(EMT)诱导中的作用,EMT是胚胎发育、肿瘤进展和损伤后纤维化组织修复的关键阶段发生的重要过程。28-32
人们普遍认为,可以证明Wnt通路的激活或影响β-连环蛋白反式激活功能的异常就地通过β-catenin积累的特异表达模式。特别是,如免疫组织化学所示,β-连环蛋白的细胞质/核积累是证明β-连环素翻译后稳定的可靠方法。33,34在本研究中,我们研究了β-catenin在一系列IPF/UIP样本中的表达模式,以及两个基因产物cyclin-D1和matrilysin/MMP-7,其表达受β-catening控制。23,24,35
材料和方法
研究人群
研究组包括20名先前未经治疗的患者,这些患者的临床、影像学(胸片和高分辨率计算机断层扫描(HRCT))、生理学和支气管肺泡灌洗检查结果均符合IPF的诊断。根据最近定义的标准,外科肺活检的组织学检查揭示了UIP的所有主要特征。1-4五份正常肺样本(肺癌大手术患者未受影响的组织碎片),两份胎儿肺样本(妊娠12周和15周),10份机化性肺炎(OP/BOOP)患者样本,4份非特异性间质性肺炎(NSIP)样本,2例急性间质性肺炎伴弥漫性肺泡损伤(AIP/DAD)和2例脱屑性间质性肺损伤(DIP),根据最新共识标准定义,4作为对照进行分析。所有样品按照标准方案固定在缓冲福尔马林和石蜡包埋中。
免疫组织化学染色和抗体
所有病例均用泛β-连环蛋白单克隆抗体(克隆15B8;西格玛化学公司,密苏里州圣路易斯)进行免疫染色。使用微波炉和0.01 mol/L柠檬酸缓冲液(pH 6.0)进行热诱导抗原回收30分钟。在自动细胞染色系统(GenoMx i6000,Biogenex)中使用敏感的抗生物素-链亲和素过氧化物酶技术(Biogenex,San Ramon,CA)处理所有样品。
为了更好地确定上皮和间充质病变的性质和分化水平,我们使用了识别低分子量细胞角蛋白8/18/19的抗体(克隆5D3,Biogenex);细支气管基底细胞中表达的细胞角蛋白-5(CK5,克隆XM26;Novocastra,Newcastle,UK);尿蛋白1,一种识别Clara细胞中CC10抗原的兔抗体(DAKO,Glostrup,丹麦);识别表面活性蛋白A的SP-A单克隆抗体(克隆PE-10,DAKO);识别α-平滑肌肌动蛋白的1A4单克隆抗体(DAKO)(无抗原回收);识别tenascin的TN2单克隆抗体(DAKO)。此外,选择了一组抗体来彻底研究表达核β-连环蛋白的损伤的分子网络特征:单克隆抗体4A4(加州圣克鲁斯圣克鲁斯生物技术公司),与人类p63的所有已知变种发生广泛反应;p40抗体(癌基因研究产品,马萨诸塞州波士顿),一种多克隆兔抗血清,专门识别缺失反式激活域的截短ΔN-p63亚型;36p53特异性单克隆抗体(克隆DO-1,DAKO);识别p21的单克隆抗体WAF1(加权平均1)(克隆SX118,DAKO)。关于这些分子在正常肺和IPF/UIP样本中的详细信息之前已经描述过。16,37最后,为了更好地评估IPF/UIP样本中Wnt/β-catenin途径激活的功能,我们在连续切片上用免疫组织化学方法研究了cyclin-D1(克隆DCS-6;Progen Biot.,Heidelberg,Germany)和matrylisin/MMP-7(克隆141-7B2;Chemicon,Temecula,CA)的表达。
免疫染色评价
正常肺组织结构(如存在)用作β-catenin免疫染色的内部对照,只有正常支气管/细支气管段显示明确的膜结合β-catentin表达(线性模式)的制剂才被认为适合解释。核染色定义为空核明显时为阴性,同时有明显的膜免疫染色,而核免疫反应时为阳性。在轻度苏木精复染制剂中,阳性细胞核的颜色从蓝色变为棕色,可以更好地评估细胞核模式。
当细胞表型包括CK5和ΔN-p63表达时,上皮病变被定义为细支气管或支气管-肺泡交界。16只有核染色对p63、ΔN-p63、p53、p21呈阳性WAF1(加权平均1).
分子分析:β-连环蛋白基因测序
三个具有代表性的IPF/UIP样品用于分子分析。使用DNeasy Tissue试剂盒(加利福尼亚州查茨沃斯市齐根)从10μm石蜡切片中提取DNA。对于序列分析,β的外显子3-连环蛋白使用应用生物系统合成器(加利福尼亚州福斯特市)合成的引物,通过聚合酶链反应扩增该基因。引物序列(ATTTGATGGTGGAGAGATGGC和CCAGCTGTTCTTGAG TGAAGG)如前所述。38在含有50 ng DNA、每个引物5 pmol、2μmol/l dNTPs、1.5 mmol/l MgCl的10μl标准反应混合物中进行聚合酶链反应2,0.5 U塔克DNA聚合酶(普罗米加,威斯康星州麦迪逊)。为了进行序列分析,使用QIAEX凝胶提取试剂盒(Qiagen)对50μl聚合酶链反应进行凝胶纯化。使用Applied Biosystem Dye Terminator Cycle-Sequencing kit(加州福斯特市Perkin Elmer)进行序列反应,并在Applied Biosystem 373型自动DNA测序仪(Perkin Elmer)上进行分析。四个毛瘤样本β-连环蛋白突变被纳入阳性对照。39
结果
正常成人和胎儿肺中β-儿茶素的细胞内表达模式
如前所述,在胎儿肺中,β-catenin核免疫染色显示在萌芽的肺泡结构中(图1a)▶.21在正常成人肺中,β-catenin的表达严格限制在所有内皮细胞和上皮细胞的细胞膜上,如清晰的线性免疫染色模式所示(图1b)▶在小比例的立方肺泡细胞中,细胞核明显聚集,根据形态学和免疫表型被识别为II型肺泡细胞,其特征是CK8/18/19和SP-a表达,CK5和ΔN-p63缺失(未显示)。
图1。
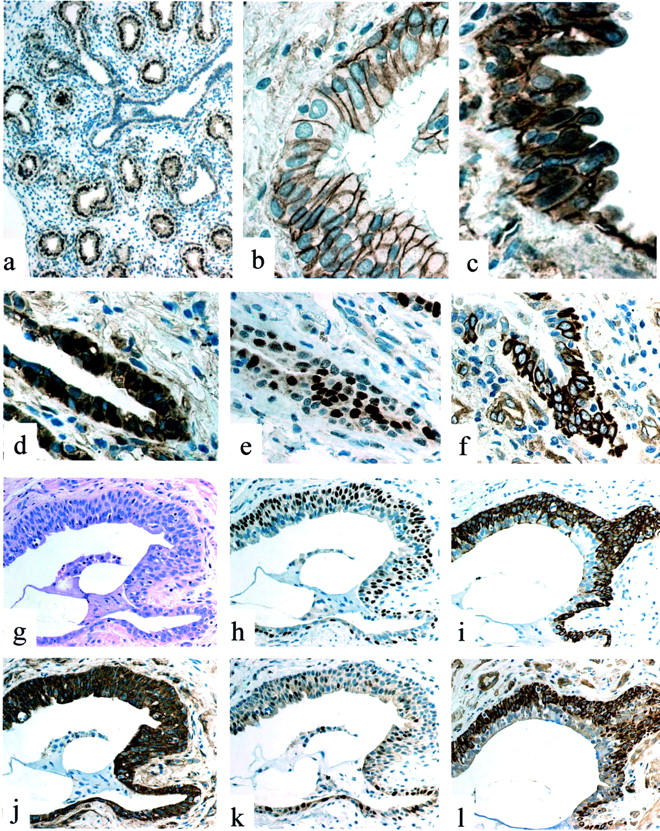
答:胎肺(12周):β-catenin的核表达在肺泡芽中明显,但在气道细胞中不明显。b:正常肺:细支气管基底细胞和纤毛细胞中β-catenin的离散膜免疫反应。抄送:IPF/UIP:增殖性细支气管病变中β-catenin的异常核聚积。日期:IPF/UIP:异常细支气管基底细胞中β-catenin的核表达。电子邮箱:IPF/UIP(串行段至d日):cyclin-D1表达细胞。f:IPF/UIP(串行段至d日):基质蛋白酶/MMP-7异常表达。克:IPF/UIP:H&E表现为小的蜂窝状细支气管病变。小时:IPF/UIP(串行段至克):ΔN-p63核表达证明基底细胞增生。i:IPF/UIP(串行段至克):CK5表达证明基底细胞增生。记者:IPF/UIP(串行段至克)基底和管腔上皮细胞中β-catenin的异常胞内表达。克:IPF/UIP(串行段至克):cyclin-D1表达增加。我:IPF/UIP(串行段至克):基质溶素/MMP-7在基底细胞中的异常表达。
IPF/UIP样本中β-连环蛋白的细胞内表达模式
在IPF/UIP患者中,表达核β-catenin的细胞数量显著增加,特别是在肺结构异常重塑明显的区域。
支气管病变
在大多数(20个样本中的18个)的增殖性支气管病变中,显示了大量表达β-catenin核聚积的上皮细胞(图1;c、d和j)▶核β-catenin积聚在这些异常结构中分布不均,主要发生在增生的基底细胞簇中。核β-连环蛋白的存在在表现出蜂窝状改变的细支气管中尤其明显(图1,g和l)▶和/或细支气管化(支气管细胞迁移到肺泡腔的过程)。有趣的是,在相同的位点,p53和p21的核过度表达waf1可以如前所述进行证明。16通过使用识别ΔN-p63和高分子量细胞角蛋白CK5的抗体,在连续切片上证实了所有这些病变的细支气管性质(图1,h和i)▶以及之前证明的表面活性剂A和CC10抗原的缺失。16
肺泡结构
在受损的肺泡结构中发现表达核β-catenin的细胞,通过连续切片的形态学和免疫表型(表面活性剂-A阳性和ΔN-p63阴性)识别为立方型II肺细胞。阳性立方形肺细胞的数量从正常到严重受累肺泡逐渐增加(图2,a和b)▶β-catenin核表达见于所有增大和/或不典型立方细胞。
图2。
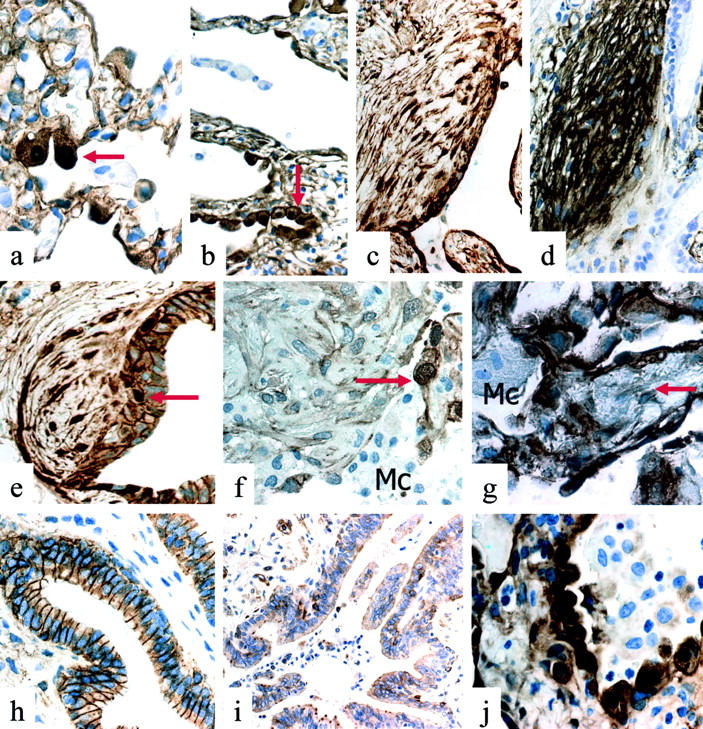
答:IPF/UIP:立方型II肺细胞中β-catenin的核蓄积(箭头)在几乎未受影响的牙槽骨结构中。b:IPF/UIP:立方型II肺细胞中β-catenin的核蓄积(箭头)严重受损的牙槽骨结构。抄送:IPF/UIP:上皮下成纤维细胞灶中多刺成纤维细胞中β-连环蛋白的核积聚。日期:IPF/UIP(串行段至c(c)):上皮下成纤维细胞病灶中纺锤形成纤维细胞中tenascin的强表达。电子邮箱:IPF/UIP:上皮下成纤维细胞病灶梭形细胞中β-catenin的核蓄积。注意β-catenin在支气管基底细胞中的核表达(箭头).传真:OP/BOOP:管腔内Masson体梭形细胞和巨噬细胞(Mc)中未观察到核β-catenin表达。注意增生的II型肺细胞中有强烈的核免疫染色(箭头).克:DAD:肺泡肺细胞中有明显的核β-catenin表达。间质成纤维细胞(箭头)巨噬细胞(Mc)缺乏明显的免疫反应性。小时:NSIP:支气管细胞膜β-catenin的表达。i:NSIP:基质蛋白酶/MMP-7在细支气管细胞中的罕见表达。记者:NSIP:核β-catenin在许多肺泡肺细胞中的表达。
成纤维细胞灶
在纺锤形细胞中观察到β-catenin的核表达,形成成纤维细胞病灶,在20个样本中有16个样本存在,这些病灶可以在连续切片上清楚识别和免疫组织化学分析(图2、c和e)▶这些病灶的特征是肌纤维母细胞,在连续切片上表现为强烈的α-平滑肌肌动蛋白和tenascin免疫反应(图2d)▶,通常位于壁内,位于异常细支气管段下方,形成严格相关的病变(图2e)▶这种模式与OP/BOOP中的肺泡内炎性息肉(Masson体)不同(图2f)▶AIP/DAD间质成纤维细胞(图2g)▶样本用作对照,其中只有少数(不到10%)梭形细胞表达核β-catenin。
细胞周期蛋白D1和基质溶素的表达
通过连续切片的免疫组织化学分析,IPF/UIP样本中可显示大量同时表达细胞周期素D1和基质溶素的细胞(图1;e、f、k和l)▶Matrilysin免疫反应在所有类型的增殖性细支气管病变中均明显,在增生性和非典型基底细胞中表现出特殊的强度(图1、f和l)▶这种模式与正常和病理对照样品明显不同,在正常和病理对照样品中,基质溶素的表达似乎不一致(图2i)▶.
细胞周期蛋白D1和基质溶素都清楚地位于与β-连环蛋白核过度表达相对应的位置,尽管这三种分子在增殖性上皮病变的序列切片上的分布并不相同。事实上,β-catenin核和/或细胞质过度表达在绝大多数细胞中都很明显(图1、d和j)▶,而基质溶素和细胞周期蛋白D1仅在一定比例的细胞中表达(图1;e、f、k和l)▶这一发现的原因尚不清楚,但可能归因于未定义的分化信号。与此观点一致的是,基质溶素和ΔN-p63在支气管病变较浅部位的细胞中停止表达(图1、h和l)▶.
β-连环素在其他间质性肺炎中的细胞内表达模式
在所有OP/BOOP、AIP/DAD、NSIP和DIP分析的病例中,细支气管病变极为罕见,免疫组织化学分析未发现β-catenin异常表达(图2h)▶因此,在细支气管上皮中未观察到基质溶素过度表达(图2i)▶另一方面,在发生广泛肺泡损伤和修复的肺泡疾病患者(如AIP/DAD、OP/BOOP和NSIP)的样本中,表达核β-catenin的肺泡肺细胞比例显著(图2j)▶.
β-连环蛋白基因序列分析
扩增区测序后,在三个IPF/UIP样本中未发现突变迹象。在同一实验中,作为已知阳性对照分析的所有四个毛母细胞瘤样本在β-连环蛋白基因。
讨论
在这项工作中,我们提供了以前未被认识到的IPF/UIP中Wnt/β-catenin信号通路参与的证据,如β-catentin在不同参与位点的广泛核积累所证明的。我们使用免疫组织化学方法揭示了β-连环蛋白的亚细胞定位,从膜位置到细胞核。这种形态学方法能够高灵敏度地检测β-catenin的细胞内再分布和核积累,并被广泛用于证明Wnt通路在人类发育和病理中的激活状态。21,33,34,40-42为了进一步支持β-连环蛋白核免疫反应的功能意义,我们研究了就地Wnt/β-catenin途径的两个靶基因cyclin-D1和matrilysin的表达,表明这两个分子在同一参与位点显著过度表达核β-catening。
最后,我们搜索了影响β-catenin基因的突变,表明β-catentin基因的异常可能与观察到的Wnt通路的激活无关,尽管我们的技术可能遗漏了小部分细胞中发生的突变。需要进一步研究IPF/UIP样本中影响Wnt/β-catenin级联相关复杂分子阵列的其他遗传和/或表达异常,包括APC、Axin和GSK3β,以及Wnt配体和卷曲的受体。43事实上,根据我们的数据,不可能确定Wnt/β-catenin激活是引起疾病还是继发于疾病。然而,本研究中研究的所有其他间质性肺疾病均无类似特征,这强烈表明了IPF/UIP中Wnt/β-catenin异常激活的致病相关性。
在本研究中,在IPF/UIP样本中的三个不同受累部位可以显示核β-连环蛋白的积聚:1)细支气管增生性病变,2)肺泡结构受损,以及3)成纤维细胞病灶。支气管增生性病变,包括基底细胞增生、鳞状化生、蜂窝状和细支气管化,在IPF/UIP中很常见,并且是该病公认的特殊特征。1-4,16根据我们的研究,Wnt通路的异常激活在这些病变中特别明显并延伸,在大多数情况下涉及基底细胞室。这个观察很有趣,因为基底细胞被认为是支气管和细支气管结构的更新成分。此外,这些发现是独特的,因为在正常肺样本和除IPF/UIP以外的各种间质性疾病中,使用相同方法研究的细支气管仅显示膜性β-连环蛋白免疫反应。另一方面,在立方增生性肺泡细胞中观察到的β-catenin核表达可被视为对肺泡损伤的生理反应的一部分,因为II型肺泡细胞在肺泡增殖/再生发生的各种条件下表现出这种模式,包括胎儿发育,21以及各种肺部疾病,如AIP/DAD和OP/BOOP,如本研究所示。
核β-连环蛋白定位于形成IPF/UIP壁内成纤维细胞病灶(纤维增生斑块)的梭形细胞中,与AIP/DAD组织性肺炎和间质肌成纤维细胞的泡内Masson小体不同。这一发现是相关的,因为Wnt通路的异常激活实际上可以提供必要的自分泌生存信号,以诱导IPF/UIP壁内成纤维细胞病灶特有的对凋亡的抵抗。7,44-46在这方面,值得注意的是,由于基因突变导致的β-连环蛋白核聚积是侵袭性纤维瘤病(硬纤维瘤)发病机制的一个中心特征,硬纤维瘤是一类与IPF/UIP成纤维细胞病灶具有某些形态学和表型特征的间叶病变,包括Wnt通路的异常激活,如本研究所示。47、48最近,一项实验研究进一步支持了我们的假设,在该实验研究中,β-连环蛋白稳定能够失调间充质细胞的增殖、运动和侵袭性,导致转基因小鼠的增殖性成纤维细胞病变。49
如上所述,β-catenin在异常核聚积模式中的分布涉及相邻隔室,主要集中在支气管-肺泡连接处。我们认为,这种特殊的带状分布可以解释UIP的一些典型组织学特征,如时间和空间异质性以及蜂窝状结构错位。1我们认为,这些数据与一种致病模型相一致,在该模型中,斑驳的外观,从早期病变到以IPF/UIP病变为特征的广泛纤维化和重塑,是由于异常β-catenin激活对生理组织修复机制的逐渐干扰而产生的,从正在进行的损伤和修复过程的病灶开始,如Wnt信号浓度梯度。Wnt靶基因的放松调控表达可能会对不同的气道成分(即细支气管和肺泡)产生不同的影响,最终导致肺泡损失和细支气管增殖。因此,在不同的系统和细胞类型中,Wnt通路的激活能够通过调节细胞周期蛋白D1和c的可用性来触发或抑制生存和死亡-myc公司,这两种蛋白质在细胞增殖和凋亡中发挥作用。50,51在这个模型中,肺泡细胞可能特别容易受到Wnt信号紊乱的影响,因为不同的分化和死亡诱导信号,包括p53、p21waf1型p63的反式激活亚型在损伤后修复肺泡中同时表达。16,52-55另一方面,通过截短的显性负性ΔN-p63亚型的组成性表达,可以保护支气管基底细胞免受凋亡的影响,从而产生强大的抗凋亡信号。16,56
Wnt通路激活在IPF/UIP中的致病作用进一步得到了就地β-catenin/LEF-1信号传导的另一个重要下游靶基因金属蛋白酶基质蛋白酶(MMP-7)异常表达的证明。最近的微阵列基因表达分析也证实了IPF/UIP中基质溶素表达的异常增加。25实验研究也提供了进一步的支持,因为苦参素基因敲除小鼠可以防止博莱霉素诱导的肺纤维化。25基质蛋白酶在细支气管增生性病变和肺重塑中的作用可归因于这种金属蛋白酶的特殊多功能作用,包括诱导上皮细胞迁移、凋亡和化生转化。57,58
在此背景下要讨论的最后一个相关主题是EMT可能参与IPF/UIP的发病机制。这些有趣的现象发生在胚胎发生和癌症进展中,使细胞能够从其来源的上皮组织中分离并自由迁移。29此外,有明确的实验证据表明,成纤维细胞可以通过上皮细胞向间充质细胞的转化直接从组织纤维化中的上皮细胞衍生而来。30,32有趣的是,如实验EMT所示,β-连环蛋白信号在诱导上皮细胞的间充质表型中发挥相关作用,28,31因此有可能认为,在IPF/UIP的细支气管病变中观察到的β-catenin异常核化可能参与EMT相关过程,增加基底细胞运动(例如,通过改变金属蛋白酶的表达),从而促进细支气管化和组织重塑。该模型的另一个有趣的可能性是,IPF/UIP中的部分异常成纤维细胞可能直接来源于正在进行的损伤/修复过程中的上皮基底细胞前体,从而形成纤维母细胞边缘和异常细支气管段严格相关的特殊病变(如图2、c和e所示▶). 有趣的是,EMT可以由细胞因子(转化生长因子-β1、成纤维细胞-生长因子-2、胰岛素样生长因子、白细胞介素-8)在实验上诱导,这些细胞因子的表达可以通过β-catenin信号的复杂调节环调节,并可能参与肺纤维化的发病机制。28,59-64
总之,尽管本研究中无法确定导致IPF/UIP中Wnt通路异常激活的确切分子机制,但我们的发现有助于破译该疾病发病的分子机制,也可能有助于寻找新的药理策略来对抗不可逆的肺重塑。事实上,目前的研究主要集中在对Wnt途径发挥调节和抑制作用的分子上。65-67这些分子可能是Wnt通路紊乱的癌症以及IPF/UIP的潜在候选新治疗策略,IPF/UI是一种传统治疗方法无法令人满意的致命疾病。
脚注
向Marco Chilosi医学博士、Patologia医学院、维罗纳大学、Policilinico G.B.Rossi、P.le L.Scuro提出转载请求,意大利维罗纳,37134。电子邮件:marco.chilosi@univr.it。
部分由意大利协会(米兰)和意大利维罗纳Cassa di Risparmio di Verona基金会提供支持。
工具书类
-
1Katzenstein AL,Myers JL:特发性肺纤维化:病理分类的临床相关性。Am J Respir Crit Care Med 1998,157:1301-1315[内政部] [公共医学] [谷歌学者]
-
2美国胸科学会:特发性肺纤维化:诊断和治疗。国际共识声明。美国胸科学会(ATS)和欧洲呼吸学会(ERS)。美国Respir Crit Care Med杂志2000,161:646-664[内政部] [公共医学] [谷歌学者]
-
三。Poletti V,Kitaichi M:特发性间质性肺炎分类的事实和争议。结节病血管性弥漫性肺病2001,17:229-238[公共医学] [谷歌学者]
-
4:美国胸科学会/欧洲呼吸学会:特发性间质性肺炎的国际多学科共识分类。美国呼吸急救医学杂志2002,165:277-304[内政部] [公共医学] [谷歌学者]
-
5Ward PA,Hunninghake GW:肺部炎症和纤维化。Am J Respir Crit Care Med 1998年,157:S123-S129[内政部] [公共医学] [谷歌学者]
-
6Kolb M、Margetts PJ、Anthony DC、Pitossi F、Gauldie J:IL-1β的瞬时表达可导致急性肺损伤和慢性修复,从而导致肺纤维化。临床研究杂志2001年,107:1529-1536[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
7Selman M,King TE,Jr,Pardo A:特发性肺纤维化:关于其发病机制和治疗意义的流行和发展假说。《内科实习年鉴》2001,134:136-151[内政部] [公共医学] [谷歌学者]
-
8Gross TJ,Hunninghake GW:特发性肺纤维化。《新英格兰医学杂志》2001,345:517-525[内政部] [公共医学] [谷歌学者]
-
9Sheppard D:肺纤维化:是细胞过度反应还是通讯失败?临床研究杂志2001年,107:1501-1502[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
10Gauldie J,Kolb M,Sime PJ:特发性肺纤维化发病机制的新方向?Resir Res 2002,3:1-3[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
11Crystal RG、Bitterman PB、Mossman B、Schwarz MI、Sheppard D、Almasy L、Chapman HA、Friedman SL、King TE,Jr、Leinwand LA、Liotta L、Martin GR、Schwartz DA、Schultz GS、Wagner CR、Musson RA:特发性肺纤维化的未来研究方向:国家心脏、肺和血液研究所工作组总结。Am J Respir Crit Care Med 2002,166:236-246[内政部] [公共医学] [谷歌学者]
-
12Gauldie J:赞成:炎症机制是特发性肺纤维化发病机制的一个次要组成部分。Am J Respir Crit Care Med 2002年,165:1205-1206[内政部] [公共医学] [谷歌学者]
-
13Strieter RM:结论:炎症机制不是特发性肺纤维化发病机制的次要组成部分。Am J Respir Crit Care Med 2002年,165:1206-1207[内政部] [公共医学] [谷歌学者]
-
14Ziesche R、Hofbauer E、Wittmann K、Petkov V、Block LH:干扰素γ-1b和小剂量泼尼松龙长期治疗特发性肺纤维化患者的初步研究。《新英格兰医学杂志》1999,341:1264-1269[内政部] [公共医学] [谷歌学者]
-
15Selman M:从抗炎药到抗纤维化药物再到肺移植:特发性肺纤维化治疗的长期研究、临床尝试和失败。胸部2002122:759-761[内政部] [公共医学] [谷歌学者]
-
16Chilosi M、Poletti V、Murer B、Lestani M、Cancellieri A、Montagna L、Piccoli P、Cangi G、Semenzato G、Doglioni C:特发性肺纤维化中的异常再上皮化和肺重塑:ΔN-p63的作用。实验室投资2002,82:1335-1345[内政部] [公共医学] [谷歌学者]
-
17Warburton D、Tefft D、Mailleux A、Bellusci S、Thiery JP、Zhao J、Buckley S、Shi W、Driscoll B:肺重塑、修复和再生是否再现了呼吸个体发育?《美国呼吸急救医学杂志》2001164:S59-S62[内政部] [公共医学] [谷歌学者]
-
18Takash W、Canizares J、Bonneaud N、Poulat F、Mattei MG、Jay P、Berta P:SOX7转录因子:序列、染色体定位、表达、反式激活和对Wnt信号传导的干扰。《核酸研究》2001,29:4274-4283[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
19Weidenfeld J,Shu W,Zhang L,Millar SE,Morrisey EE:WNT7b启动子受肺上皮中TTF-1、GATA6和Foxa2的调节。生物化学杂志2002,277:21061-21070[内政部] [公共医学] [谷歌学者]
-
20Tebar M,Destree O,de Vree WJ,Ten Have-Opbroek AA:Tcf/Lef和sFrp的表达以及β-catenin在发育中小鼠肺中的定位。机械开发2001,109:437-440[内政部] [公共医学] [谷歌学者]
-
21Eberhart CG,Argani P:Wnt信号在人类发育中的作用:胎儿肺、肾、胎盘、毛细血管、肾上腺和软骨中的β-catenin核移位。《儿科疾病与病理学》2001,4:351-357[内政部] [公共医学] [谷歌学者]
-
22Lako M、Strachan T、Bullen P、Wilson DI、Robson SC、Lindsay S:WNT11的分离、表征和胚胎表达,WNT11是一种映射到11q13.5的基因,在骨骼、肾脏和肺的发育中可能发挥作用。基因1998,219:101-110[内政部] [公共医学] [谷歌学者]
-
23Brabletz T,Jung A,Dag S,Hlubek F,Kirchner T:Beta-catenin调节人类结直肠癌中基质金属蛋白酶-7的表达。《美国病理学杂志》1999,155:1033-1038[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
24Crawford HC、Fingleton BM、Rudolph-Owen LA、Goss KJ、Rubinfeld B、Polakis P、Matrisian LM:金属蛋白酶-基质溶素是肠道肿瘤中β-catenin反式激活的靶点。癌基因1999,18:2883-2891[内政部] [公共医学] [谷歌学者]
-
25Zuo F、Kaminski N、Eugui E、Allard J、Yakhini Z、Ben-Dor A、Lollini L、Morris D、Kim Y、DeLustro B、Sheppard D、Pardo A、Selman M、Heller RA:基因表达分析显示,基质溶素是小鼠和人类肺纤维化的关键调节因子。美国国家科学院院刊2002,99:6292-6297[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
26Surendran K,McCaul SP,Simon TC:Wnt-4在肾纤维化中的作用。美国生理杂志2002,282:F431-F441[内政部] [公共医学] [谷歌学者]
-
27Shackel NA、McGuinness PH、Abbott CA、Gorrell MD、McCaughan GW:丙型肝炎病毒相关性肝硬化的病理生物学研究:肝内差异基因表达分析。《美国病理杂志》2002,160:641-654[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
28Morali OG、Delmas V、Moore R、Jeanney C、Thiery JP、Larue L:IGF-II在上皮细胞向间质转化期间诱导β-catenin快速迁移至细胞核。癌基因2001,20:4942-4950[内政部] [公共医学] [谷歌学者]
-
29Thiery杂志:肿瘤进展中的上皮-间充质转变。Nat Rev Cancer 2002年2月442-454日[内政部] [公共医学] [谷歌学者]
-
30杨杰,刘毅:肾小管上皮细胞向肌成纤维细胞转变的关键事件剖析及其在肾间质纤维化中的意义。《美国病理杂志》2001,159:1465-1475[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
31Kim K,Lu Z,Hay ED:β-catenin/LEF-1信号通路在EMT诱导中作用的直接证据。细胞生物学国际2002,26:463-476[内政部] [公共医学] [谷歌学者]
-
32Iwano M、Plieth D、Danoff TM、Xue C、Okada H、Neilson EG:组织纤维化期间成纤维细胞来源于上皮的证据。临床研究杂志2002,110:341-350[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
33Abraham SC、Klimstra DS、Wilentz RE、Yeo CJ、Conlon K、Brennan M、Cameron JL、Wu TT、Hruban RH:胰腺实性假乳头状肿瘤与胰腺导管腺癌在基因上不同,几乎总是存在β-catenin突变。《美国病理杂志》2002,160:1361-1369[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
34Brabletz T,Herrmann K,Jung A,Faller G,Kirchner T:核β-连环蛋白和c-myc的表达与肿瘤大小相关,但与结直肠腺瘤的增殖活性无关。《美国病理学杂志》2000,156:865-870[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
35Shtutman M、Zhurinsky J、Simcha I、Albanese C、D'Amico M、Pestell R、Ben-Ze'ev A:细胞周期蛋白D1基因是β-catenin/LEF-1途径的靶点。美国国家科学院院刊1999,96:5522-5527[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
36Hibi K、Trink B、Patturajan M、Westra WH、Caballero OL、Hill DE、Ratovitski EA、Jen J、Sidransky D:AIS是一种在鳞状细胞癌中扩增的癌基因。美国国家科学院院刊2000,97:5462-5467[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
37Chilosi M,Doglioni C:气道基底细胞中的组成性p63表达。弥漫性肺部疾病的分子靶点。结节病血管性弥漫性肺病2001,18:23-26[公共医学] [谷歌学者]
-
38Kitaeva MN、Grogan L、Williams JP、Dimond E、Nakahara K、Hausner P、DeNobile JW、Soballe PW、Kirsch IR:β-catenin突变在偶尔出现复制错误阳性肿瘤的结直肠癌中并不常见。癌症研究1997,57:4478-4481[公共医学] [谷歌学者]
-
39Chan EF,Gat U,McNiff JM,Fuchs E:一种常见的人类皮肤肿瘤是由激活β-catenin突变引起的。《自然遗传学》1999年,21:410-413[内政部] [公共医学] [谷歌学者]
-
40Polakis P:Wnt信号与癌症。基因开发2000,14:1837-1851[公共医学] [谷歌学者]
-
41Wang HL,Wang J,Xiao SY,Haydon R,Stoiber D,He TC,Bissonnette M,Hart J:在过度表达β-catenin的人结直肠癌中,cyclin D1和Fra-1的蛋白表达升高,但c-Myc的表达降低。国际癌症杂志2002,101:301-310[内政部] [公共医学] [谷歌学者]
-
42Kirchner T,Brabletz T:结肠腺癌序列中的模式化和核β-catenin表达。与胚胎原肠胚形成类似。《美国病理杂志》2000,157:1113-1121[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
43Wodarz A,Nusse R:Wnt信号在发育中的机制。《细胞开发生物年鉴》1998年,14:59-88[内政部] [公共医学] [谷歌学者]
-
44Lappi-Blanco E,Soini Y,Paakko P:闭塞性细支气管炎组织肺炎中新形成的纤维粘液样结缔组织的凋亡活性增加。《肺》1999年,177:367-376[内政部] [公共医学] [谷歌学者]
-
45Ueda Y、Hijikata M、Takagi S、Takada R、TakadaS、Chiba T、Shimotohno K:Wnt/beta-catenin信号抑制低血清培养基中的细胞凋亡并诱导啮齿类成纤维细胞的形态学改变。国际癌症杂志2002,99:681-688[内政部] [公共医学] [谷歌学者]
-
46King TE,Jr,Tooze JA,Schwarz MI,Brown KR,Cherniack RM:预测特发性肺纤维化患者的生存率:评分系统和生存模型。Am J Respir Crit Care Med 2001年,164:1171-1181[内政部] [公共医学] [谷歌学者]
-
47Alman BA,Li C,Pajerski ME,Diaz-Cano S,Wolfe HJ:散发性侵袭性纤维瘤病(硬纤维瘤)中β-catenin蛋白和体细胞APC突变增加。《美国病理杂志》1997,151:329-334[PMC免费文章] [公共医学] [谷歌学者]
-
48Saito T、Oda Y、Tanaka K、Matsuda S、Tamiya S、Iwamoto Y、Tsuneyoshi M:Beta-catenin核表达与散发性硬纤维瘤中cyclin D1过度表达相关。《病理学杂志》2001年,195:222-228[内政部] [公共医学] [谷歌学者]
-
49Cheon SS、Cheah AY、Turley S、Nadesan P、Poon R、Clevers H、Alman BA:Beta-catenin稳定失调间充质细胞增殖、运动和侵袭性,并导致侵袭性纤维瘤病和皮肤增生性伤口。美国国家科学院院刊2002,99:6973-6978[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
50Packham G,Cleveland JL:c-Myc和凋亡。《生物化学与生物物理学学报》1995,1242:11-28[内政部] [公共医学] [谷歌学者]
-
51Kim K,Pang KM,Evans M,Hay ED:β-catenin的过度表达诱导凋亡,与LEF-1的反式激活功能或主要G1细胞周期调控因子的参与无关。分子生物学细胞2000,11:3509-3523[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
52You Z,Saims D,Chen S,Zhang Z,Guttridge DC,Guan KL,MacDougald OA,Brown AM,Evan G,Kitajewski J,Wang CY:Wnt信号通过抑制c-Myc诱导的细胞凋亡来促进致癌转化。细胞生物学杂志2002,157:429-440[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
53Staversky RJ、Watkins RH、Wright TW、Hernady E、LoMonaco MB、D'Angio CT、Williams JP、Maniscalco WM、O'Reilly MA:氧损伤肺的正常重塑需要细胞周期蛋白依赖性激酶抑制剂p21(Cip1/WAF1/Sdi1)。《美国病理杂志》2002,161:1383-1393[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
54Adamson A、Perkins S、Brambilla E、Tripp S、Holden J、Travis W、Guinee D,Jr:弥漫性肺泡损伤中的增殖、c-myc和cyclin D1表达:在发病机制中的潜在作用和对预后的影响。《人类病理学》1999,30:1050-1057[内政部] [公共医学] [谷歌学者]
-
55Guinee D,Jr,Fleming M,Hayashi T,Woodward M,Zhang J,Walls J,Koss M,Ferrans V,Travis W:弥漫性肺泡损伤中p53和WAF1表达与细胞凋亡的关系。《美国病理杂志》1996,149:531-538[PMC免费文章] [公共医学] [谷歌学者]
-
56Yang A,McKeon F:p63和p73:p53模仿、威胁等。《Nat Rev Mol Cell Biol 2000》,1:199-207[内政部] [公共医学] [谷歌学者]
-
57Seiki M:细胞表面:基质金属蛋白酶调节迁移的阶段。《Curr Opin细胞生物学》2002,14:624-632[内政部] [公共医学] [谷歌学者]
-
58Crawford HC,Scoggins CR,Washington MK,Matrisian LM,Leach SD:基质金属蛋白酶-7由胰腺癌前体表达,并调节外分泌胰腺腺泡至导管化生。临床研究杂志2002,109:1437-1444[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
59Levy L、Neuveut C、Renard CA、Charneau P、Branchereau S、Gauthier F、Tran Van Nhieu J、Cherqui D、Petit-Bertron AF、Mathieu D、Buendia MA:白细胞介素-8的转录激活{beta}-catenin-TCF4。生物化学杂志2002,277:42386-42393[内政部] [公共医学] [谷歌学者]
-
60Lynch JP,III,Standiford TJ,Rolfe MW,Kunkel SL,Strieter RM:特发性肺纤维化中的中性粒细胞性肺泡炎。白细胞介素-8的作用。Am Rev Respir Dis 1992年,145:1433-1439[内政部] [公共医学] [谷歌学者]
-
61Liu JY,Sime PJ,Wu T,Warshamana GS,Pociask D,Tsai SY,Brody AR:转化生长因子-beta(1)肿瘤坏死因子-α受体敲除小鼠过度表达诱导纤维增生性肺疾病。Am J Respir细胞分子生物学2001,25:3-7[内政部] [公共医学] [谷歌学者]
-
62Khalil N,O'Connor RN,Flanders KC,Unruh H:TGF-β1,但不是TGF-α2或TGF-γ3,在晚期肺纤维化上皮细胞中存在差异:一项免疫组织化学研究。美国呼吸细胞分子生物学杂志1996,14:131-138[内政部] [公共医学] [谷歌学者]
-
63Strutz F、Zeisberg M、Ziyadeh FN、Yang CQ、Kalluri R、Muller GA、Neilson EG:碱性成纤维细胞生长因子-2在上皮-间充质转化中的作用。《肾脏国际》2002,61:1714-1728[内政部] [公共医学] [谷歌学者]
-
64El-Hariry I、Pignatelli M、Lemoine NR:FGF-1和FGF-2调节胰腺癌中E-cadherin和catenins的表达。国际癌症杂志2001,94:652-661[内政部] [公共医学] [谷歌学者]
-
65Bordonaro M,Lazarova DL,Augenlicht LH,Sartorelli AC:丁酸钠对Wnt信号通路的细胞类型和启动子依赖性调节。国际癌症杂志2002,97:42-51[内政部] [公共医学] [谷歌学者]
-
66Joe AK,Liu H,Suzui M,Vural ME,Xiao D,Weinstein IB:白藜芦醇在几种人类癌细胞系中诱导生长抑制、S期阻滞、凋亡和生物标志物表达的变化。《临床癌症研究》2002,8:893-903[公共医学] [谷歌学者]
-
67Hanai J、Gloy J、Karumanchi SA、Kale S、Tang J、Hu G、Chan B、Ramchandran R、Jha V、Sukhatme VP、Sokol S:内皮抑制素是Wnt信号的潜在抑制剂。细胞生物学杂志2002,158:529-539[内政部] [PMC免费文章] [公共医学] [谷歌学者]

