摘要
臂带型肌营养不良2A型(LGMD2A)是一种以进行性肌肉无力和消瘦为特征的常染色体隐性遗传疾病。LGMD2A由钙蛋白酶-3基因突变引起(第3章)对Ca进行编码2+-依赖性半胱氨酸蛋白酶主要表达于骨骼肌。潜在的病理机制尚未完全阐明。LGMD2A人类受试者的线粒体异常有不同的报道,并且在第3章-淘汰了老鼠模型。在我们的研究中,我们结合了组织化学、免疫组织化学、分子、生化和超微结构分析,以便更好地概述2例新发性LGMD2A患者的线粒体特征第3章-相关突变。两名患者均接受了详细的临床评估,随后进行了股四头肌肌肉活检。根据肌肉无力的典型临床定位,首次怀疑这两名患者中存在LGMD2A,并通过分子研究证实。发现两种新的纯合突变,c.2242C>G(p.Arg748Gly)和c.291C>A(p.Phe97Leu):c.2242C>G(p.Arg748-Gly)突变与第一位患者的线粒体质量显著减少和肌原纤维断裂有关,而c.291C>A(p.Phe97Leu)第二例患者的突变伴随着反应性线粒体增殖和碎裂红色纤维。我们的结果描绘了第3章LGMD2A线粒体功能障碍的突变特异性模式及其超微结构特征。
简介
肢体带肌营养不良型(LGMD2A)是一种常染色体隐性遗传病,其特征是肩胛和/或骨盆股骨带肌选择性缓慢进行性无力和消瘦(1). 行走障碍通常发生在诊断后的1或20年(2). 肌肉无力的平均发病时间是成年早期,范围很广,从儿童早期到生命的第五个十年(三,4). LGMD2A是由钙蛋白酶-3基因的功能缺失突变引起的(第3章),编码Ca2+,纳+-依赖性半胱氨酸蛋白酶(calpain-3)主要表达于骨骼肌(5,6). 据报道,LGMD2A是LGMD最常见的常染色体隐性遗传形式,平均占所有病例的30%-40%,显著的地理差异从美国白人的9.2%到孤立社区的80%不等(7,8). 钙蛋白酶-3主要定位于骨骼肌纤维的肌节,在那里它与巨大的蛋白质titin结合(9,10). 它也发现于肌浆网附近的三联体区域,在那里它与ryanodine受体-1(RYR-1)和钙螯合蛋白相互作用(11). 因此,钙蛋白酶-3似乎参与许多重要的生理功能。事实上,已经证明钙蛋白酶-3对控制肌节的形成、维持和重塑至关重要(12–14). 它对肌浆网摄取钙也很重要(15)细胞骨架的组织和重塑(16,17)和身体压力适应(18). 除了其蛋白水解活性外,钙蛋白酶-3还具有多种细胞过程所需的非蛋白水解结构功能,如钙的调节2+肌浆网释放(11,19). 这种二元性至少可以部分解释LGMD2A中观察到的广泛临床症状和病理特征。此外,钙蛋白酶-3可能具有大量底物,从而控制多个细胞过程,这使得进一步难以识别病理后果及其潜在的病理机制。在人类受试者中发现了线粒体功能障碍的孤立报告,因此需要进行更明确的研究,以确认这种可能的因素,从而观察到第3章突变。因此,对钙蛋白酶-3基因敲除(C3KO)小鼠进行的进一步对照研究表明,线粒体功能障碍可能是钙蛋白酶-3突变相关的常见发现(20,21). 研究表明,当C3KO小鼠反复注射心脏毒素时,再生细胞中的线粒体生物发生受损(22). C3KO小鼠肌肉中也显示出结构和功能异常,如线粒体肿胀、ATP生成减少和氧化应激增加,而线粒体生物生成增加(20). 同一组使用C3KO小鼠进行耐力运动的另一项研究表明,PGC1α水平降低,但未显示其对线粒体质量和氧化能力的影响(21). 在LGMD2A患者的报告研究中,即使使用了详细的生化、组织化学和电子显微镜研究,线粒体异常也更难确定(三,23)。
在本研究中,我们使用组织化学、免疫组织化学、生化和超微结构分析,以便更深入地了解LGMD2A患者反复观察到的线粒体异常。目前的研究是对从2名患者身上获得的肌肉活检进行的,这2名患者的肌肉活检被确定为第3章基因。我们的结果证实了这两种情况下线粒体缺陷的发生,更重要的是描绘了线粒体功能障碍的突变特异性模式及其超微结构特征。
材料和方法
患者
来自2个无亲属关系的近亲家庭(1个黎巴嫩人和1个叙利亚人)的两名年轻男性患者最近出现渐进性肌肉无力,步态和行走能力受损。两名患者均接受了详细的临床评估,随后进行了股四头肌肌肉活检。对人体材料的研究是按照1964年赫尔辛基宣言及其后来的修正案中规定的道德标准进行的。父母和患者提供了口头和书面同意书,以执行和报告诊断评估所需的所有测试。
组织化学和免疫组织化学分析
新接受的肌肉活检的方向和处理如下:将两个圆柱体在液氮冷却的2-甲基丁烷(M32631,Sigma-Aldrich,Darmstadt,Germany)中闪冻,用于横截面和纵截面的组织化学和免疫组化评估;将1个圆柱体固定在10%的福尔马林缓冲液中,进行石蜡渗透和切片;将另一个小圆柱体浸入戊二醛/多聚甲醛Karnovsky溶液中进行超微结构评估;如果需要的话,将残余组织在液氮中大量冷冻,用于生化和分子研究。
在6μm厚的切片上进行常规染色和其他特殊染色(24,25),包括苏木精和伊红(H&E)以及石蜡切片上的Masson三色染色;H&E、改良Gomori三色染色、碘酸Schiff(±淀粉酶)、油红O、pH 9.4和4.3的ATP酶、NADH、琥珀酸脱氢酶(SDH)、细胞色素c氧化酶(COX)和冷冻切片上的COX-SDH。在冰冻切片上也进行了免疫组织化学(IHC)研究,包括ATP合成酶(ab5432;1:1000;Abcam,Numelab,Beirut,Lebanon)、主要组织相容性复合体(MHC)Ⅰ类(ab52922;1:750;Abcam,Numelab)、C5b9(ab55811;1:250;Abcam-Numelab.)、,以及在年轻患者中经常观察到的一组营养不良性肌病抗体。该小组包括肌营养不良蛋白N-和C-末端和杆状结构域(NCL Dys-3,2,1;1:20;Leica Biosystems,法国巴黎)、utrophin(NCL-DRP2;1:10;Leica Biosystes)、肌多糖(NCL A、B、D、G-SARC;1:100;Leica-Biosystems)、dysferin(NCL-Hamlet 1;1:40;Leica-Piosystems,caveolin-3(610421;1:300;BD Transduction Laboratories,Madrid,Spain)、,钙蛋白酶-3(Calp2C4,1:100;徕卡生物系统公司)、RYR-1(ab59225;1:50;Abcam,Numelab)、SEPN1(ab105943;1:100;Abcam,Numelab)、FKRP(ab65243;1:50,Abcam、Numellab)、dystroglycan A(ab64568;1:50、Abcam和Numelap)、dysroglycan B(NCL-B-DG;1:五十;徕卡生态系统公司)以及层粘连蛋白α2和4(NCL-merosin;1:100,徕卡生物系统公司)。
Epon包埋切片的超微结构评价
将固定在卡诺夫斯基溶液(多聚甲醛/戊二醛)中的肌肉组织块用锇染色,然后进一步加工并嵌入塑料(Epon)中。用甲苯胺蓝对Epon块的1毫米厚切片进行染色,并在光学显微镜下进行评估,以选择具有诊断病变的特征区域。从横向和纵向的嵌入Epon的组织块上切下的选定超薄切片,用柠檬酸铅和醋酸铀酰进行进一步染色,并用透射电子显微镜进行评估。对照片进行检查,以帮助阐明两种肌肉活检中的损伤模式。
蛋白质提取和免疫印迹分析
从接受骨科护理的患者和年龄匹配的对照个体的部分肌肉活检中获取总蛋白提取物。简单地说,50 mg肌肉组织在含有Tris-HCl的PEBt3缓冲液中在pH 6.8(125 mM)、十二烷基硫酸钠(5%v/v)、乙二胺四乙酸(5 mM)、甘油(15%v/v)、DTT(100 mM)和尿素(4 M) ●●●●。然后将均质组织置于冰上10 分钟,在95°C下加热6 分钟,以最大速度离心10分钟 分钟。收集上清液,并使用Bradford试验估计蛋白质浓度(26). 蛋白质(75 μg)用来自Bio-Rad(法国Marnes-La-Coquette)的mini-PROTEAN电泳系统在4%–7%和5%–11%变性聚丙烯酰胺凝胶上分离,并印在Amersham Hybond PVDF膜上(GE Healthcare)。抗肌营养不良蛋白(Dys2 C-terminus,1:200;徕卡生物系统)和dysferlin(Hamlet,1:300;徕卡生态系统)或钙蛋白酶-3(Calp2C4,1:100;徕卡生物系统(Leica Biosystems))、小窝蛋白-3(ab2912,1:400;Abcam,Numelab)、α-肌多糖(ab189254,1:1000;Abcam,Numelab)和β-肌营养不良聚糖(NCL-b-DG,1:400,徕卡生物系统)抗体鸡尾酒分别用于探测4%–7%和5%–11%的膜。GAPDH(ab9484,1:1500;Abcam,Numelab)用作装载控制。使用了过氧化物酶结合的抗兔或抗鼠二级抗体(分别为ab6721和ab6789,1:1000;Abcam,Numelab)。使用基于化学发光的方法检测信号(Clarity Western ECL;Bio-Rad)。
线粒体活性的生化评估
使用基于酶的分光光度法评估两名患者的线粒体呼吸链复合物活性(复合物I-V)和基质相关酶(例如柠檬酸合酶)。对液氮速冻肌肉活检部分制备的匀浆进行生化评估。如前所述,使用Cary 50紫外可见分光光度计(安捷伦科技公司;法国莱斯乌利斯瓦里安公司)进行分光光度测定(27)。
分子研究
从冷冻肌肉样品中提取基因组DNA(20 mg)按照制造商的说明使用QIAamp DNA Mini Kit(51304;德国希尔顿市齐根市)。第3章使用覆盖24个外显子和50个外显基因的面板,通过下一代测序分析基因序列 侧翼内含子区的bp。测序基于Illumina测序技术,所有显著变异均通过Sanger测序进行验证。
结果
临床和神经系统结果
患者1为9岁 岁时,他第一次评估身体逐渐虚弱和越来越不稳定(图1A). 血液分析显示肌酸激酶(CK)水平升高3966 IU/L,每月随访逐渐增加,达到4977 IU/L,然后6750 IU/L.当CK水平达到7000时,安排第一次肌肉活检 随访第三个月的IU/L。最初是在一家小医院进行的,没有特殊程序的规定。没有对冰冻切片进行评估,石蜡切片被诊断为“炎症性肌病与多发性肌炎相一致”。父母拒绝接受类固醇治疗,并带孩子去咨询儿科神经科医生,后者发现骨盆带肌无力比通常的近端肌无力表现得更为严重,没有典型的高尔动作标志。这名儿童的步态也有障碍,行走困难。他的“步态不稳”很可能反映了额外的远端运动障碍。神经科医生指出,他的哥哥,12岁 现年9岁的受试者在进行临床评估时,表现出类似但更为严重的症状和9岁时开始的运动障碍 岁。为哥哥做的诊断工作被拒绝了。一个弟弟,然后4岁 岁,临床表现正常。父母是表亲,但近亲属中没有类似的运动功能障碍报告。从左侧股外侧肌进行肌肉活检,并在新鲜状态下迅速送到我们的神经肌肉诊断实验室进行评估。
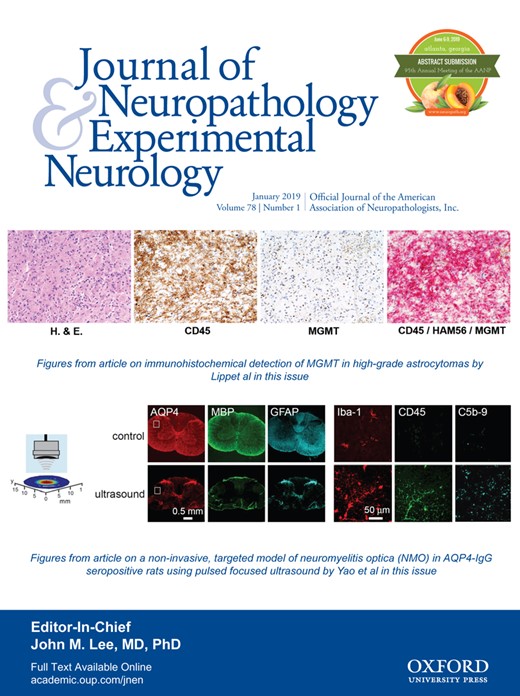
图1。
基于LGMD2A特征的2个独立家族的系谱。(A)黎巴嫩一家,有三个兄弟姐妹,来自近亲父母(表亲)。(B)叙利亚一家有4个兄弟姐妹,来自近亲父母(一级堂兄弟姐妹)。探针用箭头表示。ID号下方的阿拉伯数字代表诊断年龄。填充符号和空符号分别表示临床受影响(LGMD2A阳性)和未受影响的个体。双线表示近亲婚姻(一级表亲)。
患者2 25岁 岁时,他因左腿开始的渐进性虚弱病史来到神经科诊所7 几年前(图1B). 临床检查显示颅神经功能正常,但弥漫性广泛性肌肉萎缩保留斜方肌。上肢轴向和近端无力的特征为3+/5,翼状突出。深部肌腱反射消失。他步态蹒跚,从蹲姿站立时明显虚弱,轻度感觉异常和麻木,无临床肌强直。他的CK水平是1955 首次评估时的IU/L和1734 一个月后随访IU/L。所有其他实验室评估,包括ESR、ANA和TSH水平,均在正常范围内。肌电图(左三角肌、左趾总伸肌、右股内侧肌和右胫前肌)显示严重的全身性肌病,左三角体和左趾伸肌的复视增加,低振幅和短持续时间的运动单位,右胫前肌多发性痉挛增加,伴有一次强直性放电。神经传导研究没有显示任何神经病变的证据。对左股四头肌进行了肌肉活检,结果表明存在如下所述的营养不良过程。进行了重复的临床评估2 几年后,包括右肱二头肌、右股内侧肌、右胫前肌和右腓肠肌的重复肌电图,均表明严重的进行性全身肌病影响到所有肢体的近端和远端肌肉,与不断发展的营养不良过程相一致。他的家族史与此相关,因为他的父母都是一级堂兄弟,但他的三个弟弟都没有临床证据显示出类似的营养不良过程。
肌肉活检:组织化学和免疫组织化学特征
作为诊断检查的一部分,对两名患者进行了左股四头肌(股外侧肌)的肌肉活检。病理评估显示两名患者都有营养不良特征,线粒体特征各不相同。患者1肌肉活检显示肌纤维大小和形状有明显变化,有散在的大肌营养不良纤维、显著的肌内膜纤维化和成簇的小再生肌纤维(图2). 纤维分裂和肌原纤维断裂频繁。末端补体复合物(C5b9)沉积突出显示多灶性小面积肌纤维坏死。炎症浸润,主要是CD8+T淋巴细胞和CD68+巨噬细胞,CD20较少+B淋巴细胞散在坏死肌纤维附近的肌内膜中,也可见于小动脉周围的肌周。沉积MHC I(MHC-1)涂层再生肌纤维。使用一组与儿童期发病的肌营养不良相关的蛋白抗体进行免疫组织化学评估,结果表明,除了钙蛋白酶-3和RYR-1外,在远离坏死灶的完整和再生肌纤维上的蛋白表达正常,而钙蛋白酶-3与RYR-1则明显减少。线粒体呼吸链酶活性(包括NADH-TR、SDH和COX)的组织化学评估显示,线粒体活性显著降低,多灶性活性严重丧失使大多数肌纤维呈现“蛾样”外观(图2). 免疫组化染色显示ATP合成酶也显著降低。
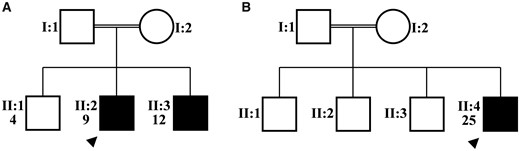
图2。
肌肉活检组织学(患者1)。苏木精-伊红(H&E)和改良Gomori三色(GT)染色显示营养不良特征,包括纤维大小变化、纤维分裂和肌内膜纤维化。还观察到肌纤维坏死和肌噬(黑色箭头)以及异常再生纤维(白色箭头)。COX和SDH组织酶反应表现出斑块状,“蛾食”状,反映线粒体损失和肌原纤维网络破坏。上面板和下面板分别表示截面放大100倍和200倍。比例尺:50 微米。
患者2肌肉活检显示广泛的营养不良特征,肌纤维大小和形状有明显变化,大量分裂肌纤维,再生肌纤维,大量内化细胞核和肌内膜纤维化(图3). 还观察到与患者1相似的多灶性小面积肌纤维坏死和小再生肌纤维簇。免疫组织化学染色显示,肌肉营养不良症中最常见的蛋白表达谱正常,但钙蛋白酶-3除外,钙蛋白酶-3完全缺失。然而,一些病理特征是患者2特有的。线粒体酶组织化学染色显示,许多大型营养不良性肌纤维的中央区域苍白,而在一些肌纤维内线粒体活性轻度增加的外周肌瓣下缘内,线粒体密度和大小增加。少量I型肌纤维内COX活性完全丧失。还发现散在的碎红纤维和许多分叶状纤维(图3)。
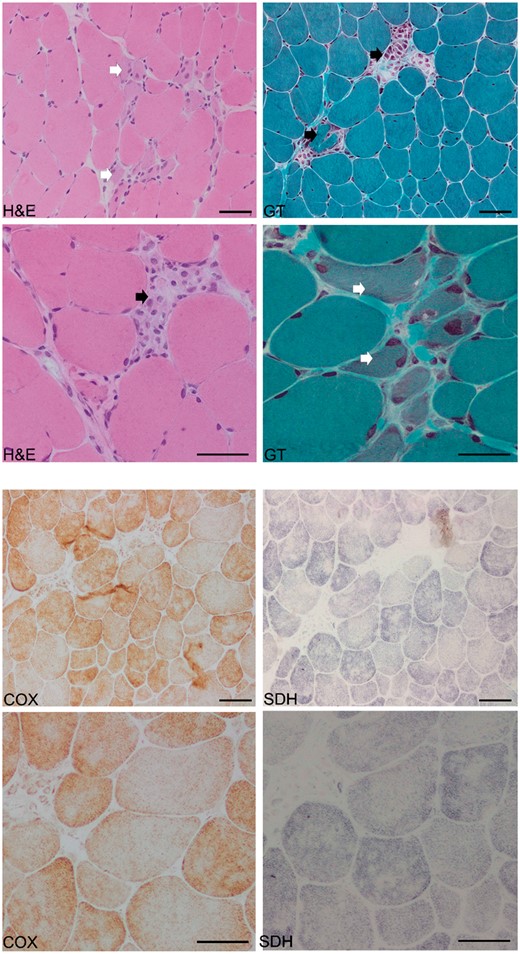
图3。
肌肉活检组织学(患者2)。苏木精-伊红(H&E)和改良Gomori三色(GT)染色显示肌内膜纤维化、纤维分裂、肌纤维大小和形状的显著变化以及碎裂的红色纤维簇(白色星号)。组织酶学染色(COX、SDH和COX/SDH)显示,许多大型营养不良性肌纤维的中央区域苍白,线粒体(黑色星号)的肌膜下堆积,以及2%肌纤维(黑色箭头)内COX活性偶尔丧失。上下面板分别表示截面放大100倍和200倍。比例尺:50 微米。
透射电子显微镜研究
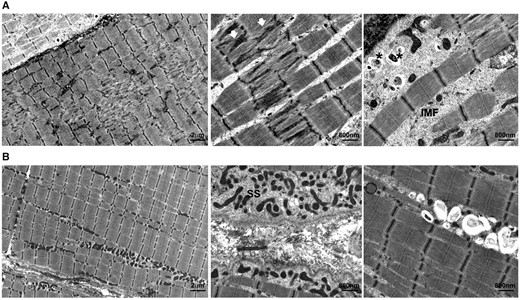
对患者1的活检进行的超微结构研究显示,肌原纤维断裂的多灶性小区域,让人联想起小孔,线粒体缺失,肌丝在流动加厚的Z带中失去方向。在许多肌原纤维间隙和肌膜下“泡状物”内也观察到成簇的小而收缩的、通常是固缩的线粒体,其中一些包含在吞噬细胞空泡内,两者都因游离的、非溶酶体结合的糖原颗粒堆积而膨胀(图4A)。
图4。
先证者肌肉活检的超微结构分析。(A)在患者1中,左面板(×2900)和中面板(×9300)切片显示斑片状肌原纤维断裂Z轴带状流(白色箭头)。肌原纤维间(IMF)和肌下膜(SS)间隙中发现异常的常固缩线粒体(右幅,×9300)(一些包含在自噬空泡中)(星号)。两名患者扩张的IMF间隙和SS泡内的糖原颗粒密度均增加,患者1更明显。(B)在患者2中,左面板(×2900)和中面板上部(×9300)切片显示小而细长的线粒体局部SS和IMF增生,内部轮廓模糊,其中一些包含自噬空泡(右面板;×9300。未发现肌原纤维断裂。
在2号患者中,超微结构研究显示肌下膜和肌原纤维间小的、卵形或细长的线粒体增殖增加。一些线粒体偶尔出现固缩和塌陷,一些线粒体在自噬细胞空泡中部分退化为膜状(图4B). 与患者1相比,未发现肌原纤维紊乱。
多重免疫印迹分析
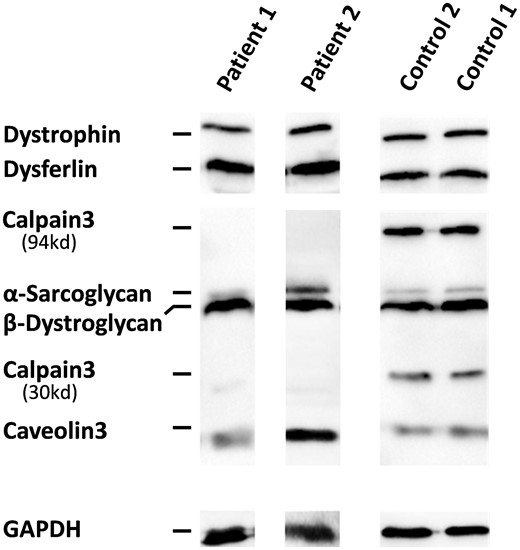
为了进一步验证和扩展免疫组织化学发现,我们随后通过多重免疫印迹评估了钙蛋白酶-3的稳态水平以及几种常见于各种肌肉营养不良的骨骼肌蛋白。如所示图5蛋白质分析表明,与对照组相比,这两名患者的钙蛋白酶-3蛋白完全缺失。其他评估蛋白,肌营养不良蛋白、dysferrin、α-肌多糖、β-肌营养不良聚糖和小窝蛋白-3(18kDa)正常表达。
图5。
多重免疫印迹分析。与对照组相比,多重免疫印迹法的蛋白质分析显示两名患者中完全没有钙蛋白酶-3。Dystrophin(423 kDa)、dysferrin(200 kD)、α-肌多糖(50 kDa。GAPDH被用作负荷控制。
线粒体活性的生化评估
通过酶分光光度法对两名患者的冷冻肌肉部分进行线粒体呼吸链复合体活性的定量评估。患者1的所有呼吸链复合体的活性显著降低。如参考酶柠檬酸合成酶的活性所示,活性降低是线粒体大量消耗的次要原因(表)。患者2的所有活动均在正常范围内,线粒体含量保持在正常范围,如该患者柠檬酸合成酶的活动所示。
分子分析
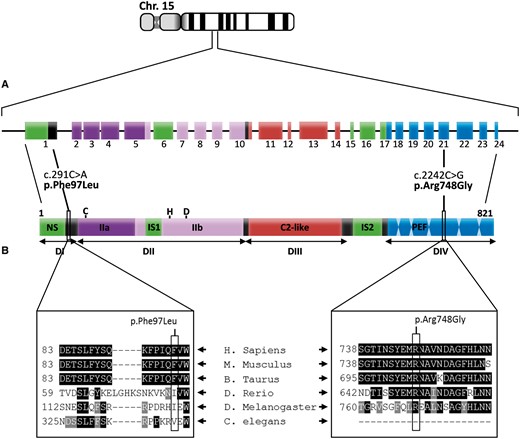
临床和组织学评估以及免疫印迹分析表明,潜在的营养不良过程提示钙蛋白酶-3这两名患者均存在肢体束带肌的缺陷和临床表现。因此,我们试图通过对钙蛋白酶病的全长进行测序来确认其与钙蛋白酶病之间的因果关系第3章两名患者的基因。通过高通量测序,我们在患者1和患者2中分别鉴定了2个新的纯合错义突变c.2242C>G(p.Arg748Gly)和c.291C>A(p.Phe97Leu)(图6). 两种致病性纯合子变体均经Sanger测序证实。此外,对携带致病性变体的患者1的另一个家庭成员(兄妹)的鉴定证实了其与杂合父母的分离。无法对我们完全失去联系的叙利亚难民家庭进行进一步的分子分析。这两种变异体在CADD等工具中都有很强的致病性预测(http://cadd.gs.washington.edu)、SIFT(http://sift.jcvi.org)和PolyPhen-2(http://genetics.bwh.harvard.edu). 多重序列比对分析表明,第一个突变,c.2242C>G,导致PEF(Penta E-F hand)结构域中已知结合Ca的甘氨酸取代高度保守的氨基酸精氨酸(p.Arg748)2+并参与蛋白质二聚体。第二个突变导致N-末端结构域中的亮氨酸残基取代了不太保守的苯丙氨酸氨基酸,该结构域是催化核心的一部分(图6)。
图6。
DNA测序第3章基因和多序列比对。(A)Calpain-3基因定位于第15号染色体(15q15.1)。它由24个外显子组成,编码821个氨基酸多域蛋白酶。对患者1和患者2的calpain-3基因进行高通量测序,分别鉴定出2个新的纯合突变c.2242C>G(p.Arg748Gly)和c.291C>A(p.Phe97Leu)。突变c.2242C>G定位于外显子21,涉及一个氨基酸残基,该氨基酸残基是与Ca有关的PEF(Penta E-F hand)结构域的一部分2+结合和蛋白质二聚体。突变c.291C>A定位于外显子1,涉及一个氨基酸残基,该氨基酸残基是包含核定位信号的N末端(NS)结构域的一部分。(B)不同生物体氨基酸的多重序列比对表明,p.Arg748残基在物种间具有进化保守性,而p.Phe97残基仅在哺乳动物中具有保守性。使用带有默认参数的在线软件Clustal Omega(EMBL-EBI)实现对齐。
讨论
隐性突变第3章据报道,LGMD2A是LGMD最常见的形式之一。据报告,不同地理区域的发病率增加,其临床和病理变化显著。在同一家族内报告的病例中,观察到的发病率、进展、临床和病理特征也存在差异。根据莱顿肌营养不良突变数据库,到目前为止,已鉴定出约500种不同的致病突变,大多数是错义突变(50%-60%)。
据我们所知,这是首次研究黎巴嫩LGMD2A的分子、生化、结构和超微结构基础。我们发现了两种新的纯合子错义突变第3章两个无亲属关系的家庭。第一个突变,c.2242C>G(p.Arg748Gly),在一个黎巴嫩近亲(表亲)家庭的一名9岁儿童中发现,而第二个突变c.291C>a(p.Phe97Leu),在另一个叙利亚近亲(一级表亲)家庭成员中发现。
这两名患者的临床表现各不相同,可能受发病年龄和疾病进展率的影响,也可能与基因突变的部位和类型有关第3章每个患者的基因。然而,两名患者的典型肌肉无力定位相似,主要累及骨盆带的轴向肌肉,并逐渐发展至肩带和腿部远端肌肉,从而损害步态和行走能力。肌肉活检组织学研究显示,肌营养不良模式主要表现为肌纤维大小变化增加、小再生和萎缩肌纤维的局灶簇、偶尔变性和坏死的肌纤维、肌内膜纤维化增加以及少量炎症浸润。此外,在这两种情况下,常发现同时表达utrophin和dystrophin的肌纤维,可能表明受累肌纤维的成熟延迟或中断。
通过IHC和多重免疫印迹分析,两名患者均显示钙蛋白酶-3蛋白缺失。然而,在2名患者中的每一名患者的诱导线粒体异常中都观察到了差异。突变c.2242C>G的特征是出现明显的“蛾形”外观,出现片状肌原纤维断裂,线粒体氧化酶活性在生化评估中显著降低。这种线粒体活性降低最可能是由于(1)线粒体质量显著下降,反映为柠檬酸合成酶活性显著下降,以及(2)超微结构研究中观察到的线粒体明显固缩,反映出残余线粒体明显缺乏功能。突变c.291C>A在肌纤维间和肌肉瘤下间隙诱导了显著的线粒体增殖,组织化学染色显示为“粗糙的红色”和“分叶的”肌纤维。这种线粒体增殖通常在肌肉活检中观察到,以证实疑似线粒体功能障碍,并被广泛用作潜在线粒体功能障碍的诊断组织学特征。散在的肌纤维中COX活性的丧失也被注意到。值得注意的是,虽然组织化学染色和超微结构分析表明,在c.291C>A突变的情况下,线粒体分布受到破坏,但线粒体酶活性在生物化学研究中保持在正常范围内(表)。如我们在超微结构研究中观察到的,这些结果很可能反映了线粒体的这种反应性增殖可能作为早期防御机制触发,以补偿正在进行的崩解和损失(图4). 这些发现强调了在评估线粒体活性时结合组织化学和生物化学研究的重要性,因为可能更好地描述细微的改变。在c.2242C>G突变的情况下,未观察到线粒体的肌膜下堆积和“分叶状纤维”。
| . | 活动(nmol/分钟 毫克)
. | . |
|---|
| . | 患者1(c.2242C>G). | 患者2(c.291C>A). | 参考范围(nmol/min 毫克). |
|---|
| CI公司 | 5.10* | 24.01 | [10.42–47.30] |
| CII公司 | 2.44* | 31.33 | [18.73–47.70] |
| CIII公司 | 25.80* | 124.99 | [81.28–210.86] |
| CIV公司 | 17.98* | 187.92 | [82.01–237.59] |
| 个人简历 | 20.09* | 100.72 | [62.21–130.72] |
| CII+III级 | 8.50* | 32.38 | [16.21–33.24] |
| 柠檬酸合成酶 | 50.18* | 142.25 | [110.86–288.65] |
| 乳酸脱氢酶 | 4382 | 4979 | [2084–7317] |
| . | 活动(nmol/分钟 毫克)
. | . |
|---|
| . | 患者1(c.2242C>G). | 患者2(c.291C>A). | 参考范围(nmol/min 毫克). |
|---|
| CI公司 | 5.10* | 24.01 | [10.42–47.30] |
| CII公司 | 2.44* | 31.33 | [18.73–47.70] |
| CIII公司 | 25.80* | 124.99 | [81.28–210.86] |
| CIV公司 | 17.98* | 187.92 | [82.01–237.59] |
| 个人简历 | 20.09* | 100.72 | [62.21–130.72] |
| CII+III级 | 8.50* | 32.38 | [16.21–33.24] |
| 柠檬酸合成酶 | 50.18* | 142.25 | [110.86–288.65] |
| 乳酸脱氢酶 | 4382 | 4979 | [2084–7317] |
| . | 活动(nmol/分钟 毫克)
. | . |
|---|
| . | 患者1(c.2242C>G). | 患者2(c.291C>A). | 参考范围(nmol/min 毫克). |
|---|
| CI公司 | 5.10* | 24.01 | [10.42–47.30] |
| CII公司 | 2.44* | 31.33 | [18.73–47.70] |
| CIII公司 | 25.80* | 124.99 | [81.28–210.86] |
| CIV公司 | 17.98* | 187.92 | [82.01–237.59] |
| 个人简历 | 20.09* | 100.72 | [62.21–130.72] |
| CII+III级 | 8.50* | 32.38 | [16.21–33.24] |
| 柠檬酸合成酶 | 50.18* | 142.25 | [110.86–288.65] |
| 乳酸脱氢酶 | 4382 | 4979 | [2084–7317] |
| . | 活动(nmol/分钟 毫克)
. | . |
|---|
| . | 患者1(c.2242C>G). | 患者2(c.291C>A). | 参考范围(nmol/min 毫克). |
|---|
| CI公司 | 5.10* | 24.01 | [10.42–47.30] |
| CII公司 | 2.44* | 31.33 | [18.73–47.70] |
| CIII公司 | 25.80* | 124.99 | [81.28–210.86] |
| CIV公司 | 17.98* | 187.92 | [82.01–237.59] |
| 个人简历 | 20.09* | 100.72 | [62.21–130.72] |
| CII+III级 | 8.50* | 32.38 | [16.21–33.24] |
| 柠檬酸合成酶 | 50.18* | 142.25 | [110.86–288.65] |
| 乳酸脱氢酶 | 4382 | 4979 | [2084–7317] |
超微结构研究使我们能够查明两种突变之间的另一种不同表型。在c.2242C>G突变(患者1)的情况下,肌原纤维断裂和糖原积聚斑块内线粒体数量减少。这种肌原纤维的断裂让人想起SEPN1和RYR-1突变时经常观察到的核心和小核。有趣的是,患者1的RYR-1表达显著减少,患者2的RYR-1表达正常(补充数据图S1). 这种变化可能是由控制RYR-1表达的钙蛋白酶-3功能的特定缺失触发的。在c.291C>A突变的情况下,肌原纤维的结构和组织得到了较好的保留,线粒体的增殖能力得到了保留,尽管线粒体存活似乎有所缩短,导致自噬空泡中出现了固缩线粒体。
LGMD2A的特点是在地理背景之间,甚至在具有相同突变的兄弟姐妹之间具有显著的临床和病理变异性(28). 导致这种差异的确切原因,主要是与线粒体异常有关的原因,仍不清楚。过去几年的几项研究试图通过描述钙蛋白酶-3复杂的相互作用网络,从而揭示其潜在的生物学作用,从而揭示钙蛋白酶-3的这些有趣特性。研究表明,钙蛋白酶-3通过其催化活性在肌肉再生能力中发挥重要作用,主要是在生理应激条件下。当肌节伸展时,钙蛋白酶-3将其位置从Titin的M线区域转移到N2A区域,在那里它进行蛋白质水解并激活与调节信号转导有关的几个蛋白质,以应对外部应激(18,29,30). 因此,钙蛋白酶-3活性的丧失可以特别解释我们两位患者的退行性过程。
一些观察也表明钙蛋白酶-3在肌肉成熟和分化中起着重要作用(31). 我们的研究结果支持这一作用,因为这两名患者的肌纤维都表现出延迟成熟。Kramerova等人进一步证明第3章被击倒(第3章-KO)小鼠模型,Ca2+/钙调素依赖性蛋白激酶II(Ca-CaMKII)损伤导致PGC-1α的高效上调(21). 后者是一种转录辅激活子,调节线粒体生物学的广泛程序,其中包括其呼吸能力(32). 此外,心脏毒物诱导的坏死模型表明,AMP-activated protein kinase(AMPK)在第3章-KO再生肌纤维(32). 众所周知,ATP缺乏的细胞传感器AMPK可以刺激慢纤维型基因表达和氧化代谢,同时通过直接磷酸化抑制mTORC1活性。在骨骼肌中,mTORC1增强肌原纤维蛋白合成和线粒体生物生成,引导细胞生长和增殖(33,34). 因此,AMPK对mTORC1的抑制与再生过程中PGC-1α转录物和线粒体含量未能增加有关第3章-KO肌肉与WT相比(32)。
在LGMD2A患者中第3章-在分析的病例中,线粒体生理和结构的相关突变变化很大,甚至与小鼠中获得的突变不同。Nilsson等人使用酶分析和蛋白质印迹技术,在LGMD2A患者的肌肉活检中显示出保留的呼吸链功能(35). 然而,应该谨慎对待他们的数据,因为只有一个子集(14个中的5个)的患者进行了酶分析,并且只报告了这些获得的值的平均值,而不是单个值,这些值的平均值可能与正常病例中记录的值显着不同。此外,未进行线粒体组织化学研究。因此,与线粒体的假定局灶性丢失或多灶性增殖的可能相关性,如在线粒体功能障碍的病例中经常注意到的,并未概述。在怀疑存在细微改变的情况下,例如局部增殖和/或变性,这种相关性非常重要。在Pyle等人的另一项研究中,发现一名LGMD2A患者的骨骼肌出现呼吸链功能障碍,出现COX-阴性纤维,复合IV活性轻度降低(36)。
我们的数据表明,一些致病性变体,例如c.2242C>G,模拟第3章-线粒体异常中KO相关表型,而其他,例如c.291C>A,诱导不同表型。因此,除了第3章-KO小鼠、其他因素和途径也可能与线粒体功能障碍的发生有关CAPN(大写字母)3种突变。似乎突变的位置、改变的氨基酸的性质以及对钙蛋白酶-3蛋白的结构、稳定性和各种活性的诱导结果都深刻地影响了疾病的进展并决定了疾病的结局。一些第3章研究表明,所描述的突变可以加速钙蛋白酶-3的自溶活性,甚至在其完成各种活性之前就导致其丢失(37,38)而其他突变则以改变底物识别和/或翻译后调控位点的方式影响钙蛋白酶-3构象(19,39). 进一步的突变可能只影响钙蛋白酶-3的蛋白水解活性,而不会改变其非蛋白水解功能(11)。
我们发现的第一个错义突变c.2242C>G定位于外显子21,涉及进化保守氨基酸p.Arg748,它是已知结合Ca的钙蛋白酶-3 PEF结构域的一部分2+并参与蛋白质二聚化以及底物识别。第二个错义突变c.291C>A定位于外显子1,涉及催化核心的N末端结构域的氨基酸p.Phe97。与Arg748氨基酸相反,Phe97在哺乳动物中主要保守,而在其他物种中则有所不同。无论是通过免疫印迹还是通过IHC,我们都观察到钙蛋白酶-3蛋白完全丢失。在可能不同的临床和病理结果的起源上,确定导致线粒体异常的机制仍然是假设的。需要新的见解来进一步了解多种致病机制及其观察到的变异性。
我们的数据表明第3章-相关突变可能导致各种信号通路的缺陷激活和快速改变,最终导致潜在的突变特异性线粒体缺陷。基因修饰物的存在也可能在临床结果的变异性中发挥作用,即使在同一家族成员中也是如此。继发性缺陷可能会显著促进特定临床表型的发展。这些发现应该提高人们对新型靶向治疗的认识。阐明与肌营养不良发病和进展相关的功能性病理生理过程可能有助于改进现有治疗方法的临床应用,而不依赖于疾病的主要病因。
这项工作得到了黎巴嫩国家科学研究中心(L-CNRS)的支持,奖项编号:103508。
作者没有双重性或利益冲突需要声明。
补充数据可以在以下位置找到academic.oup.com/jnen。
参考文献
1法宁
M(M)
,安吉利尼
C、。
2A型肢带型肌营养不良症的蛋白质和基因诊断:产量和陷阱
。肌肉神经
2015
;52
:163
–73
2布什比
公里。
肌肉营养不良
。Bailliers Clin Neurol公司
1994
;三
:407
–30
三蔡
J型
,米纳米
N个
,金
Y(Y)
,等。
Calpain 3基因突变:边缘肌营养不良症的遗传和临床病理研究
。神经肌肉疾病
2001
;11
:54755
4斯特劳布
V(V)
,贝尔托利
M。
我们在常染色体隐性肢带型肌营养不良症的试验准备中处于什么位置?
神经肌肉疾病
2016
;26
:111
–25
5戈尔
判定元件
,汤普森
心室颤动
,锂
H(H)
,等。
钙蛋白酶系统
。生理学评论
2003
;83
:731
–801
6小野
Y(Y)
,小岛
K(K)
,托里
F类
,等。
骨骼肌特异性钙蛋白酶是一种细胞内Na+依赖性蛋白酶
。生物化学杂志
2010
;285
:22986
–98
7乌尔塔松
M(M)
,萨恩斯
A类
,鲁多
C类
,等。
吉普斯科(西班牙巴斯克国家)的肢体带状肌营养不良
。大脑J神经
1998
;121
:1735
–47
8周
佛罗里达州
,安吉利尼
C类
,丹特尔
D类
,等。
异质性边缘肌营养不良人群Calpain III突变分析
。神经病学
1999
;52
:1015
–20
9返町
H(H)
,金巴拉
K(K)
,木村
S公司
,等。
肌肉特异性钙蛋白酶,p94,与2A型肢带肌营养不良有关,通过IS2(p94特异性序列)与连接素相关
。生物化学杂志
1995
;270
:31158
–62
10凯拉
Y(Y)
,野口勇
S公司
,米纳米
N个
,等。
钙蛋白酶3在人骨骼肌中的定位及其在肢带肌营养不良2A肌中的改变
。生物化学杂志(东京)
2003
;133
:659
–64
11小岛
K(K)
,小野
Y(Y)
,奥滕海姆
C类
,等。
钙蛋白酶-3在骨骼肌肌浆网中的非营利功能
。分子生物学杂志
2011
;407
:439
–49
12克拉默洛娃
我
,库德里亚肖娃
E类
,文卡特拉曼
G公司
,等。
钙蛋白酶3通过作用于泛素蛋白酶体途径上游参与肌节重塑
。人类分子遗传学
2005
;14
:2125
–34
13贝克曼
JS公司
,斯宾塞
M。
Calpain 3,正确肌节组装、翻转和维护的“看门人”
。神经肌肉疾病
2008
;18
:913
–21
14巴格达人
S公司
,马丁
M(M)
,理查德
我
,等。
Calpain 3缺乏与2A型缘肌营养不良症的肌核细胞凋亡和IkappaBα/NF-kappaA通路的深度扰动有关
。自然·医学
1999
;5
:503
–11
15米歇尔
LYM公司
,霍恩德洛普
JGJ公司
,宾德尔斯
RJM公司。
钙蛋白酶-3介导的钠调节+-钙2+交换器亚型3
。Pflugers拱门
2016
;468
:243
–55
16盖恩
年少者
,库德里亚肖娃
E类
,波茨
A类
,等。
钙蛋白酶3裂解丝蛋白C并调节其与γ-和δ-淀粉聚糖相互作用的能力
。肌肉神经
2003
;28
:472
–83
17克拉默洛娃
我
,贝克曼
JS公司
,斯宾塞
乔丹。
钙蛋白酶病的分子和细胞基础(2A型肢体带状肌营养不良症)
。Biochim生物物理学报
2007
;1772
:128
–44
18小岛
K(K)
,川端康成
Y(Y)
,Nakao公司
H(H)
,等。
肌肉特异性钙蛋白酶在小鼠体内的动态分布在生理应激适应中起着关键作用,并在肌营养不良中受损
。临床研究杂志
2010
;120
:2672
–83
19小野
Y(Y)
,小岛
K(K)
,Ouchi新开
F类
,等。
一种偏心钙蛋白酶,CAPN3/p94/钙蛋白酶-3
。生物化学
2016
;122
:169
–87
20克拉默洛娃
我
,库德里亚肖娃
E类
,吴
B类
,等。
线粒体异常、能量缺乏和氧化应激是骨骼肌钙蛋白酶3缺乏的特征
。人类分子遗传学
2009
;18
:3194
–205
21克拉默洛娃
我
,埃尔莫洛娃
N个
,埃斯金
A类
,等。
未能上调肌肉适应所需基因的转录是肢带型肌营养不良2A(钙蛋白酶病)的基础
。人类分子遗传学
2016
;25
:2194
–207
22亚尔瓦奇
我
,阿莫伦维特
J型
,布拉甘扎
C类
,等。
钙蛋白酶-3缺失肌肉再生受损与mTORC1信号传导紊乱和线粒体生物发生缺陷有关
。骨骼肌
2017
;7
:27
23卡瓦伊
H(H)
,Akaike公司
M(M)
,库尼西格
M(M)
,等。
来自三个日本家庭的七名患者中新calpain 3基因突变的2A型边缘肌营养不良症的临床、病理和遗传特征
。肌肉神经
1998
;21
:1493
–501
24拉夫林
M。
肌肉活检:实验室调查
牛津:巴特沃斯·海尼曼;1993 26布拉德福德
MM。
利用蛋白质-眼睛结合原理快速灵敏地定量微克蛋白质
。Ana Biochem公司
1976
;72
:248
–54
27鲁斯汀
P(P)
,克里特岛
D类
,布尔热龙
T型
,等。
呼吸链缺陷的生化和分子研究
。临床化学学报
1994
;228
:35
–51
28古列里
M(M)
,马格里
F类
,达安吉洛
MG公司
,等。
意大利肢体束带型肌营养不良症患者大样本的临床、分子和蛋白质相关性
。Hum变种
2008
;29
:258
–66
29小岛
K(K)
,小野
Y(Y)
,Doi公司
N个
,等。
肌发生期、肌节长度和蛋白酶活性调节肌肉特异性钙蛋白酶的定位
。生物化学杂志
2007
;282
:14493
–504
30林下(Hayashi)
C类
,小野
Y(Y)
,Doi公司
N个
,等。
多分子相互作用表明连接蛋白/肌动蛋白N2A区是骨骼肌p94/calpain 3活性的调节支架
。生物化学杂志
2008
;283
:14801
–14
31斯宾塞
美赞臣
,盖恩
年少者
,返町
H(H)
,等。
体内肌肉转基因钙蛋白酶3的稳定表达:转基因小鼠的未成熟肌肉表明钙蛋白酶3在肌肉成熟中的作用
。《美国科学院院刊》
2002
;99
:8874
–9
32罗伊
GC公司
,Patten公司
是
,兹森盖勒
ZK公司
,等。
PGC-1缺乏骨骼肌线粒体含量与呼吸链容量的断开
。单元格代表
2013
;三
:1449
–56
33阿瑟顿
PJ公司
,巴布拉杰
J型
,史密斯
K(K)
,等。
AMPK-PGC-1-alpha或PKB-TSC2-mTOR信号的选择性激活可以解释耐力或阻力训练类肌肉电刺激的特定适应性反应
。美国财务会计准则委员会J
2005
;19
:786
–8
34巴尔
英国。
耐力和力量训练:细胞信号的教训
。锻炼与运动医学与科学
2006
;38
:1939
–44
35尼尔森
医疗保险
,麦克尼尔
LG公司
,北冈
Y(Y)
,等。
LGMD2A患者骨骼肌氧化还原状态和线粒体呼吸链功能
。公共科学图书馆一号
2014
;9
:电子102549
36派尔
A类
,南丁格尔
HJ公司
,格里芬
H(H)
,等。
非线粒体疾病的呼吸链缺陷
。神经遗传学
2015
;1
:e6(电子6)
37小野
Y(Y)
,岛田
H(H)
,返町
H(H)
,等。
2A型臂架肌营养不良相关突变引起的肌肉特异性钙蛋白酶p94的功能缺陷
。生物化学杂志
1998
;273
:17073
–8
38迪亚兹
BG公司
,莫尔多瓦努
T型
,柯伊伯
美赞臣
,等。
肌肉特异性钙蛋白酶p94的插入序列1作为一种内部前肽
。生物化学杂志
2004
;279
:27656
–66
39凯拉
Y(Y)
,野口勇
S公司
,Kurokawa村
R(右)
,等。
基因表达谱对2A型肢带型肌营养不良症分叶状纤维的表征
。神经科学研究
2007
;57
:513
–21
©2018美国神经病理学家协会。保留所有权利。






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}