")
")
主要功能和细节
检测类型:三明治 检测方法:比色法 平台:缩微板阅读器 分析时间:3小时30分钟 样品类型:核提取物,纯化蛋白质 灵敏度:1 ng
概述
-
产品名称 -
检测方法 比色法 -
样品类型 核提取物,纯化蛋白质 -
分析类型 三明治 -
敏感 >1纳克 -
范围 500纳克- 20000纳克 -
检测时间 3小时 3000万 -
物种反应性 与以下物质反应: 人类、植物、哺乳动物、真菌、其他物种 -
产品概述 通用SIRT活性测定试剂盒(比色法)是一整套优化的缓冲液和试剂,用于使用来自哺乳动物、植物、真菌和细菌等多种物种的核提取物或纯化的SIRT亚型(SIRT 1-7)来测量总SIRT酶的活性/抑制,其形式多种多样, 但不限于培养细胞和新鲜及冷冻组织。 检测SIRT的抑制或激活对于阐明基因激活和沉默的表观遗传调控机制非常重要,可能有助于癌症或神经疾病的诊断和治疗。 通用SIRT活性检测试剂盒(比色法)提供了成功实现这一目标的组件。 -
站台 缩微板阅读器
属性
-
储存说明 请参阅协议。 -
组件 48次测试 96次测试 10X洗涤缓冲液 1 x 14毫升 1 x 28毫升 8孔试纸条(带框架) 1 x 6个单元 1 x 12件 胶粘覆盖膜 1台 1台 捕获抗体,1000X 1 x 5微升 1 x 10微升 检测抗体,2000X 1 x 6微升 1 x 12微升 开发人员解决方案 1 x 5毫升 1 x 10毫升 HDAC抑制剂,50µM 1 x 50微升 1 x 100微升 SIRT分析缓冲液 1 x 4毫升 1 x 8毫升 SIRT分析标准,50µg/mL 1 x 10微升 1 x 20微升 SIRT系数,50倍 1 x 50微升 1 x 100微升 SIRT抑制剂,50 mM 1 x 40微升 1 x 80微升 SIRT基板,50X 1 x 60微升 1 x 120微升 停止解决方案 1 x 5毫升 1 x 10毫升 -
研究领域 -
功能 NAD依赖性蛋白脱乙酰酶,将转录调控直接与细胞内能量学联系起来,并参与多种分离细胞功能的协调,如细胞周期、对DNA损伤的反应、代谢、凋亡和自噬。 可以通过组蛋白的去乙酰化调节染色质功能,并可以促进组蛋白和DNA甲基化的改变,导致转录抑制。 脱乙酰化广泛的转录因子和协同调节因子,从而对目标基因的表达进行正调控和负调控。 作为NAD(+)/NADH细胞溶质比率的传感器,该比率因葡萄糖缺乏和与热量限制相关的代谢变化而改变。 对骨骼肌细胞分化至关重要,对低营养素的反应介导对骨骼肌成肌细胞分化的抑制作用,这也涉及5'-AMP活化蛋白激酶(AMPK)和烟酰胺磷酸核糖转移酶(NAMPT)。 eNoSC(能量依赖性核仁沉默)复合物的成分,该复合物可调节rDNA沉默以响应细胞内能量状态,并通过招募组蛋白修饰酶发挥作用。 eNoSC复合物能够感知细胞的能量状态:当葡萄糖饥饿时,NAD(+)/NADP(+)比率升高会激活SIRT1,导致组蛋白H3脱乙酰化,然后由SUV39H1在“Lys-9”(H3K9me2)将H3二甲基化,并在rDNA位点形成沉默染色质。 将SUV39H1的“Lys-266”去乙酰化,导致其活化。 通过脱乙酰PCAF和MYOD1抑制骨骼肌分化。 HIST1H1E的去乙酰化H2A和'Lys-26'。 组蛋白H4的去乙酰化酶“Lys-16”(体外)。 通过染色质重塑参与NR0B2/SHP共表达功能:通过NR0B2/SHP招募到LRH1靶基因启动子,从而刺激组蛋白H3和H4脱乙酰化,导致转录抑制。 建议通过积极调节端粒长度来促进基因组完整性; 然而,关于端粒周围异染色质定位的报道是相互矛盾的。 建议通过调节核SUV39H1的可用池,在组成异染色质(CH)的形成和/或维护中发挥作用。 氧化/代谢应激通过MDM2抑制SUV39H1多泛素化来减少SUV39H2降解。 SUV39H1水平的增加增加了CH中SUV39H2的周转,这反过来似乎加快了异染色质的更新,异染色素在应激反应期间与更大的基因组完整性相关。 去乙酰化p53/TP53的“Lys-382”并损害其诱导转录依赖性促凋亡程序和调节细胞衰老的能力。 脱乙酰化TAF1B,从而通过RNA聚合酶I抑制rDNA转录。脱乙酰化MYC,促进MYC与MAX的结合,降低MYC的稳定性,导致转化能力受损。 去乙酰化FOXO3以应对氧化应激,从而提高其诱导细胞周期阻滞和抗氧化应激抵抗的能力,但抑制FOXO3-介导的凋亡转录活性诱导; 也导致FOXO3泛素化和蛋白体降解。 在调节转录活性和凋亡方面,似乎对MLLT7/FOXO4有类似的影响。 去乙酰化物DNMT1; 从而损害DNMT1甲基转移酶诱导的依赖性转录抑制因子活性,调节DNMT1细胞周期调节功能和DNMT1介导的基因沉默。 脱乙酰化酶RELA/NF-kappa-B p65从而抑制其反式激活潜能并增强对TNF-α的反应中的凋亡。 脱乙酰HIF1A、KAT5/TIP60、RB1和HIC1。 去乙酰化FOXO1导致其核保留并增强其转录活性,从而增加肝脏中的糖异生。 抑制E2F1转录活性和凋亡功能,可能通过去乙酰化。 参与HES1和HEY2介导的转录抑制。 与MYCN合作似乎参与了DUSP6/MAPK3的转录抑制,从而通过“Ser-62”磷酸化使MYCNs稳定。 脱乙酰MEF2D。 对可能与组蛋白H3的局部脱乙酰化有关的AR依赖基因的拮抗剂介导的转录抑制所必需。 抑制HNF1A介导的转录。 CREBZF用于抑制ESRRG。 调节AP-1转录因子活性。 脱乙酰化物NR1H3和NR1H2以及‘Lys-434’处的NR1H_3脱乙酰化正调控NR1H3:RXR靶基因的转录,促进NR1H3-蛋白体降解并导致胆固醇外流; 提出了转录到达轮后启动子清除机制。 参与脂质代谢。 通过抑制PPARG(可能与NCOR1和SMRT/NCOR2相关)调节白色脂肪细胞的脂肪生成和脂肪动员。 去乙酰化ACSS2导致其活化,以及HMGCS1。 参与肝脏和肌肉代谢。 通过去乙酰化和激活PPARGC1A,在低血糖条件下激活骨骼肌中的脂肪酸氧化,并参与葡萄糖稳态。 参与肝脏PPARA和脂肪酸β氧化的调节。 参与葡萄糖对胰岛β细胞胰岛素分泌的正向调节; 该功能似乎意味着UCP2的转录抑制。 建议去乙酰化IRS2,从而促进其胰岛素诱导的酪氨酸磷酸化。 去乙酰化SREBF1亚型SREBP-1C,从而降低其在脂肪生成基因表达中的稳定性和反式激活。 通过抑制参与DNA修复的基因(如XPC和TP73)参与DNA损伤反应,脱乙酰化XRCC6/Ku70,促进补充额外因子到受损DNA位点,如SIRT1-脱乙酰化NBN可以招募ATM来启动DNA修复,SIRT1-去乙酰化XPA与RPA2相互作用。 还通过同源重组和特异性单链退火(独立于XRCC6/Ku70和NBN)参与DNA双链断裂的DNA修复。XPC的转录抑制可能涉及E2F4:RBL2抑制复合物和蛋白激酶B(AKT)信号。 TP73的转录抑制可能涉及E2F4和PCAF。 脱乙酰化WRN,从而调节其解旋酶和核酸外切酶活性,并调节WRN核转位以应对DNA损伤。 在“Lys-6”和“Lys-7”处脱乙酰化APEX1,并通过促进APEX1与XRCC1的结合来刺激细胞AP内切酶活性。 通过阻断细胞质p53/TP53的核转位并可能将其重定向至线粒体,增加p53/TP33介导的转录依赖性凋亡。在“Lys-539”和“Lys-542”处脱乙酰化XRCC6/Ku70,使其将BAX与线粒体隔离,从而抑制应激诱导的凋亡。 可能通过去乙酰化ATG5、ATG7和MAP1LC3B/ATG8参与自噬。 去乙酰化AKT1,导致AKT1和PDK1与PIP3的结合增强,并促进其活化。 建议在调节与细胞衰老相关的STK11/LBK1依赖性AMPK信号通路中发挥作用,这似乎涉及调节STK11/LBK1的乙酰化状态。 能去乙酰化STK11/LBK1,从而增加其活性、细胞质定位和与STRAD的结合; 然而,这种活性在正常细胞中的相关性尚不清楚。 在内皮细胞中显示出抑制STK11/LBK1活性并促进其降解。 在“Lys-64”和“Lys-70”处脱乙酰SMAD7,从而促进其降解。 去乙酰化CIITA并增强其MHC II类转录激活,并有助于其稳定性。 终止MECOM/EVI1。 在“Lys-487”处脱乙酰化PML,这种脱乙酰化促进PML对PER2核定位的控制。 在神经原性转换期间,通过抑制选择性NOTCH1靶基因 异构体2:异构体2显示为p53/TP53的去乙酰化'Lys-382',但活性低于异构体1。 结合起来,这两种亚型发挥相加作用。 异构体2调节p53/TP53的表达和细胞应激反应,并被p53/TP3抑制,呈现SIRT1异构体依赖的自身调节环。 (微生物感染)如果HIV-1感染,与病毒Tat蛋白相互作用并使其脱乙酰。 病毒Tat蛋白抑制SIRT1对RELA/NF-kappa-B p65的脱乙酰活性,从而增强其转录活性,SIRT1被认为有助于感染期间T细胞的过度激活。 SirtT1 75kDa片段:催化失活的75SirT1可能参与细胞凋亡的调控。 可能通过与细胞色素C结合和干扰凋亡小体组装,参与保护软骨细胞免受凋亡死亡。 -
组织特异性 广泛表达。 -
序列相似性 属于sirtuin家族。 I类亚科。 包含1个脱乙酰酶sirtuin型结构域。 -
翻译后 修改 多个赖氨酸残基甲基化; DNA损伤后甲基化增强,对p53/TP53的脱乙酰酶活性来说是必不可少的。 磷酸化。 STK4/MST1磷酸化,导致SIRT1介导的p53/TP53脱乙酰化受到抑制。 丝氨酸-27、丝氨酸-47和丝氨酸-530的MAPK8/JNK1磷酸化导致核定位和酶活性增加。 DYRK1A和DYRK3在Thr-530处的磷酸化激活脱乙酰酶活性并促进细胞存活。 雷帕霉素复合物1(mTORC1)在Ser-47的哺乳动物靶点磷酸化抑制脱乙酰活性。 被CaMK2磷酸化,导致p53/TP53和NF-kappa-B p65/RELA脱乙酰活性增加(通过相似性)。 涉及MAPK9的Ser-27磷酸化与蛋白质稳定性有关。 一些磷酸化位点存在一些模糊性:Ser-159/Ser-162和Thr-544/Ser-545。 经TNF-α处理后,组织蛋白酶B进行蛋白质水解,生成催化活性但稳定的SirtT1 75kDa片段(75SirT1)。 GAPDH对S-亚硝基化,导致抑制NAD依赖性蛋白脱乙酰酶活性。 -
细胞定位 细胞质。 线粒体和细胞核,PML体。 细胞质。 核心。 通过与PML的互动招募到核机构(PubMed:12006491)。 在细胞核中用APEX1染色(PubMed:19934257)。 可在核仁、核常染色质、异染色质和内膜中发现(PubMed:15469825)。 在细胞核和细胞质之间穿梭(通过相似性)。 利用XBP1亚型2在细胞核中进行结肠酸化(PubMed:20955178)。 -
UniProt提供的信息 -
替代名称 75信号T1 hSIR2型 hSIRT1型
查看所有内容 -
数据库链接
相关产品
-
相关产品
图像
-

通用SIRT活性检测试剂盒(ab156915) 用SIRT分析标准生成的图示标准曲线。 -
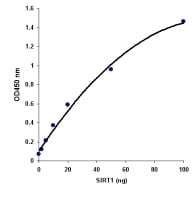
通用SIRT活性检测试剂盒(ab156915) 用重组SIRT1和ab156915证明SIRT活性测定的高灵敏度。
协议
数据表和文件
-
SDS下载 -
数据表下载
工具书类 (16)
龚H 等。 miR-146a通过NAMPT抑制和NAD+/SIRT失活阻碍AMPK的抗衰老作用。 信号传输目标热 7:66 (2022). 公共医学:35241643 太阳S 等。 微小RNA-138-5p通过抑制sirtuin 1信号传导来驱动心力衰竭的进展。 分子医学代表 23:不适用(2021年)。 公共医学:33576462 Lee IS公司 等。 4-己基间苯二酚给药对Saos-2细胞NAD+水平和SIRT活性的影响。 Maxillofac整形外科 43:39 (2021). 公共医学:34719767 严平(Yan P) 等。 LARP7通过变构增强SIRT1脱乙酰酶活性来改善细胞衰老。 单元格代表 37:110038 (2021). 公共医学:34818543 张X 等。 CD38在缺氧/缺血条件下通过转录抑制途径引起自噬通量抑制和心脏功能障碍。 前电池开发生物 8:191 (2020). 公共医学:32363189