介绍 脊髓刺激(SCS)是一种向脊髓传递电脉冲的神经调节技术 通过 放置在硬膜外腔的电极。 它已经实施了50多年,用于治疗慢性神经病和顽固性疼痛[ 1 ].
据估计,全球每年约有50000名慢性疼痛患者接受SCS治疗[ 2 ]. 基于感觉异常的SCS机制是基于Melzack和Wall提出的痛觉调制门控制理论。 据推测,电脉冲会激活大直径纤维(Aβ),阻止小直径纤维向大脑传输有害信息[ 三 , 4 ]. 根据门控制理论,大直径和小直径初级感觉纤维之间的相互作用可能发生在脊髓的同一节段,因此,传递疼痛信号的初级感觉神经元的SCS部位和背根神经节(DRG)应该位于同一节或更高的节段。 其他研究人员提供了证据,证明脊髓上区域也参与SCS诱导的镇痛[ 5 – 7 ]. 许多研究的结果表明,许多中枢神经系统(CNS)衍生的神经递质,如γ-氨基丁酸[ 8 ]、血清素[ 5 , 9 ],大麻素1受体[ 10 , 11 ]和阿片肽[ 12 – 14 ]参与SCS诱导的镇痛。
我们对针刺的神经化学机制、电针(EA)以及经皮穴位电刺激(TEAS)的相关技术感兴趣。 我们之前的研究结果表明,电针通过激活内源性阿片系统以频率依赖的方式诱导镇痛:2 Hz电针选择性增加脑内脑啡肽的基因表达和释放,脑啡肽与μ-阿片受体(MOR)和δ-阿片样受体(DOR)结合,激活下行抑制通路, 产生镇痛效果。 相反,100 Hz的电针选择性地增加脊髓中强啡肽的基因表达和释放,并激活κ阿片受体(KOR)[ 15 – 17 ]. 我们想知道SCS是否也通过激活内源性阿片系统来诱导镇痛,因为初级传入纤维可以被周围神经末梢的EA或背根和/或背角的SCS激活。 有几条证据支持这一假设。 例如,SCS患者的疼痛缓解和脑脊液(CSF)中β-内啡肽升高同时发生[ 14 ]. 动物实验表明,4Hz SCS产生的镇痛作用被MOR拮抗剂纳洛酮阻断,而60Hz SCS的镇痛作用则被DOR特异性拮抗剂纳曲酮阻断[ 13 ]. 在50 Hz的SCS后,脊髓中的痛阈和强啡肽A(1–13)水平均升高[ 12 ].
这些研究强烈表明内源性阿片肽参与SCS镇痛。 然而,在不同频率的SCS中参与SCS的阿片受体的类型尚未完全阐明。 EA和SCS是否具有相同或相似的神经化学机制有待研究。 此外,频率非常高的SCS(如10kHz)的镇痛效果受到患者的好评,但其机制尚不清楚。
本研究旨在确定(1)SCS是否在神经性疼痛的大鼠备用神经损伤(SNI)模型中产生镇痛作用,以及SCS频率特异性的镇痛特征,(2)内源性阿片肽及其相应受体是否参与其中, 尤其是在10 kHz的SCS中。
材料和方法 动物 138只雄性Sprague-Dawley大鼠体重160±10 g,给予啮齿动物食物和水 随意 每个笼子容纳1-4个。 室温保持在20–25°C,湿度约为50%,12:12小时的明暗循环(07:00时灯亮)。 所有动物实验程序均经北京大学动物护理和使用委员会批准(伦理批准ID:LA2020434),并符合美国国立卫生研究院《实验动物护理和使用指南》。
外科 备用神经损伤(SNI) 采用改良的SNI神经病理性疼痛模型。 用2%-3%异氟醚麻醉大鼠。 左后肢的胫神经和腓总神经用4–0丝轻轻结扎,腓肠神经保持完整,类似于Decosterd和Woolf描述的模型[ 18 ]. 缝合并闭合上覆肌肉和皮肤。
脊髓刺激系统导线的植入 SNI手术后三天(图。 1 A) ,对表现出机械性超敏反应的大鼠进行L1和T12水平的椎板切除术,这对应于L4–L6腰椎脊髓区域。 专门为大鼠设计的SCS导线(中国北京品思医疗有限公司)包含三个间隔2 mm的接触点(图。 2 A) ●●●●。 该导线沿嘴侧方向插入硬膜外,并用缝合线固定在肌肉和韧带上。 伤口用缝合线闭合,导线的远端从颈部底部的皮肤中穿出,并固定在头部,以便稍后连接到外部神经刺激器(T902,北京品思医疗有限公司,中国)。 以2 Hz的频率进行刺激,如果电极放置正确,则可以获得中下躯干、同侧后肢或尾巴的运动收缩。
图1。
实验设计。 A类 SNI模型的动物准备以及SCS电极和鞘内导管的植入。 B类 30分钟SCS程序。 C类 不同强度的SCS。 D类 不同频率的短期SCS和CSF采集。 电子 24小时不同频率的SCS和CSF采集。 脊髓刺激; PWT,爪子收回阈值; SNI,避免神经损伤; MT,电机阈值; 脑脊液。
图2。
导线植入说明和导线位置的X射线验证。 A类 导线尺寸和植入导线的位置。 B类 植入电极相对于椎骨和肋骨的位置通过X射线验证。 通过计算肋骨的数量来识别T13–L1椎骨:13 第个 肋骨吻侧附着在13只 第个 胸椎。
在研究之前,测试了所有动物的阻抗和运动阈值(MT)。 阻抗表示电极在脊髓-硬膜外间隙中的功能状态,0–1 kΩ表示两个触点之间存在短路,1–10 kΩ表示电极处于工作状态,1–10MΩ表示两触点之间存在开路状态。 在SCS之前测量,平均阻抗为3.99±0.11 kΩ(图。 三 A) 电极植入后3天。 11天后,它增加到~6 kΩ,并保持在这个水平。 通过缓慢增加SCS的振幅(以0.01-mA为增量),从零开始以2 Hz,200μs的脉冲宽度,直到观察到中下躯干或后肢的肌肉收缩,来测定MT。
图3。
SCS在神经病理性疼痛大鼠模型中镇痛作用的表征。 A类 , B类 阻抗变化的时间进程 A类 和电机阈值 B类 SCS电极植入后。 C1类 在50 Hz脊髓刺激30分钟期间和之后,重度和中度机械性超敏大鼠的50%PWT。 指挥与控制 平均50%PWT由 C1类 .Pre-SCS,0 min时50%PWT in C1类 ; 在SCS期间,15分钟和30分钟的平均PWT C1类 ; SCS后,60 min时50%PWT C1类 用Dunnett的多重比较检验进行重复方差分析** P(P) < 0.01 与 预-SCS; ### P(P) < 0.001 与 Dur-SCS中度和重度机械过敏之间的50%PWT。
鞘内导管植入术 导线植入后,用23号针在L5或L6水平穿刺椎管。 将一根32号PE–10导管(Smiths Medical ASD,Inc.,USA)引入蛛网膜下腔,并向前推进3.5–4厘米,到达腰部扩大处。 导管固定在筋膜上,其远端皮下穿通,然后附着在肩胛骨之间的皮肤上,用于鞘内给药。 为了从生理上确定导管尖端的正确位置,大鼠在处死后被解剖。 99%的大鼠的导管末端位于蛛网膜下腔L4–L6水平。
机械敏感性评估 按照Dixon的描述,对大鼠的爪子撤退阈值(PWT)进行了测试[ 19 ]. 将大鼠放置在带有金属网底部的塑料笼中,使其完全接触到爪子,以适应约30分钟的环境,直到笼探索和主要梳理活动停止。 测试区域是腓肠神经分布中的后爪外侧表面。 使用校准的von Frey纤维测试PWT,弯曲力范围为0.4至15.0 g(8根纤维:0.4、0.6、1.0、2.0、4.0、6.0、8.0和15.0 g)。 根据Dixon的升降法,在系列中间开始使用2.0 g灯丝进行测试。 根据大鼠是否抬起爪子决定退出。 如果出现正撤回,则施加额外的减小力。 如果出现负撤回,则施加更大的力。 使用查普兰描述的50%PWT方法计算PWT 等 . [ 20 ].
30分钟SCS程序 SNI手术后7天,这些动物开始接受SCS(图。 1 A) ●●●●。 X射线显示的位于T13–L1椎体中的触点被选作阳极和阴极(图。 2 B) ●●●●。 选择三个电极中的任意两个覆盖包含T13–L1段的区域。 通常,中间触点被设置为阴极。 将大鼠放在金属丝网上约30分钟进行习服,然后测定基线50%PWT。 之后,实施了30分钟的SCS; 在SCS启动后的15分钟和30分钟,以及SCS终止后的15 min和30分钟再次获得50%的PWT(图。 1 B) ●●●●。
入选和排除标准 如果基线50%PWT<1 g或>6 g,则将SNI大鼠排除在主要研究之外。然而,将50%PWT<1 g的动物作为严重机械超敏的亚组进行分析。 电极未覆盖X光检查证实的T13–L1水平的动物也被排除在外。
实验设计 不同机械敏感性水平下的SCS 将动物分为以下组:第1组-严重(50%PWT值<1 g, n个 =4)和第2组-中度(50%PWT介于1和6 g之间, n个 = 4). 使用50Hz、脉冲宽度为200μs(恒流模式下的双相对称方波)和80%MT的刺激。
不同强度的SCS(20%、40%、60%和80%MT) 中度机械性超敏反应的动物被随机分为以下组:第1组-对照组(No-SCS)( n个 = 7); 第2组–SCS( n个 = 7). 第2组动物接受刺激,强度设置为20%、40%、60%和80%MT(图。 1 C) ●●●●。 采用50 Hz和200μs脉冲宽度的SCS(恒流模式下的双相对称方波)。
不同频率(2、15、50、100和2/100 Hz以及10 kHz)下的短期SCS 将动物分为以下组:第1组-对照组(No-SCS)( n个 =9); 第2组–SCS( n个 = 11). 在两次实验之间的24小时冲洗期内,对每只动物进行了多次频率测试。 为了避免偏差,测试频率的顺序安排为:2只大鼠接受SCS,顺序为2、15、50、100 Hz、10 kHz和2/100 Hz(密集分散波形,频率介于2和100 Hz之间,各持续3 s[ 17 ])(图。 1 D) ; 2只大鼠,频率为15、50、100 Hz、10 kHz、2/100和2 Hz; 2只大鼠,频率为50、100 Hz、10 kHz、2/100、2和15 Hz; 2只100Hz、10kHz、2/100、2、15和50Hz的大鼠; 2只10kHz、2/100、2、15、50和100 Hz的大鼠; 和1只2/100、2、15、50、100 Hz和10 kHz的大鼠。 频率研究中使用了在80%MT下刺激200μs(10 kHz下30μs)脉冲宽度(恒流模式下的双相对称方波)。
在不同频率(2、50、100 Hz和10 kHz)下连续24小时的SCS 将动物随机分为以下组:第1组-对照组(No-SCS)( n个 = 6); 第2组–SCS( n个 =6)。 在每只大鼠中测试多个频率,任何两个测试频率之间有24小时的洗脱期。 为了避免偏差,测试频率的顺序安排为2只大鼠按顺序2、50、100 Hz和10 kHz接受SCS(图。 1 E) ; 2只100Hz、10kHz、2和50Hz的大鼠; 和2只10kHz、2Hz、50Hz和100Hz的大鼠。
在研究习服前30分钟,将大鼠放置在带有金属丝网底部的试验笼中,然后测定基线50%PWT。 然后,在80%MT下24小时SCS,施加200μs(10 kHz下30μs)脉冲宽度(恒流模式下的双相对称方波),并在24小时连续SCS期间每2小时测量50%PWT。
选择性阿片受体拮抗剂的实验设计 阿片受体拮抗剂D-Phe-Cys-Tyr-D-Trp-Orn-Thr-Pen-Thr-NH 2 (CTOP,ab120078,美国Abcam)和去甲萘酚胺(nor-BNI,P5296,美国Sigma)新鲜溶解在盐水中。 Naltrindole(NTI,B6429,APExBIO,USA)新鲜溶解在10%二甲基亚砜中。 给药前将所有药物溶液预热至37°C。 静脉注射药物后,用10μL预先加热的生理盐水冲洗导管。 在SNI手术后11、12、13、14、15和16天,在SCS前30分钟(2100 Hz和10kHz)注射拮抗剂,在测试的任何两个频率之间有24小时的冲洗期。 在80%MT电流和200μs脉冲宽度(10 kHz时为30μs)下获得50%的PWT。
各实验组的治疗总结见表 1 CTOP实验中的大鼠被分为三组:第1组-幼稚(no-SNI, n个 = 9); 第2组-对照组(SNI,no-SCS, n个 =9),和组3–SCS(SNI,SCS, n个 = 9). 为了避免偏差,测试频率的顺序安排为3只大鼠按2、100 Hz和10 kHz的顺序接受SCS(表 2 ); 3只100Hz、10kHz和2Hz的大鼠; 以及3只分别为10kHz、2 Hz和100Hz的大鼠。
表1。
实验组 SNI公司 SCS公司 敌手
天真无邪 – – –
无SCS + – –
SCS+车辆 + + –
SCS+CTOP 3μg + + +
SCS+CTOP 10μg + + +
SCS+CTOP 30μg + + +
SCS+NTI 3 mg/kg + + +
SCS+NTI 10 mg/kg + + +
SCS+Nor-BNI 5.74μg + + +
SCS+Nor-BNI 22.96μg + + +
表2。
第1阶段 第2阶段 第3阶段
敌手 SCS(赫兹) 时间 敌手 CTOP(微克) SCS公司 (赫兹) 时间 敌手 CTOP(微克) SCS(赫兹) 时间
载具(10μL) 2 第8天 三 2 第11天 10 2 第14天
100 第9天 10 100 第12天 30 100 第15天
10公里 第10天 10 10千 第13天 30 10公里 第16天
NTI实验中的大鼠被分为三组:第1组-幼稚(no-SNI, n个 = 9); 第2组-对照组(SNI,no-SCS, n个 =11)和第3组(SNI、SCS, n个 = 10). 为了避免偏差,测试频率的顺序安排为3只大鼠按2、100 Hz和10 kHz的顺序接受SCS(表 三 ); 4只100Hz、10kHz和2Hz的大鼠; 和3只10kHz、2Hz和100Hz的大鼠。
表3。
第1阶段 第2阶段 第3阶段
敌手 SCS(赫兹) 时间 敌手 NTI(mg/kg) SCS(赫兹) 时间 敌手 NTI(mg/kg) SCS(赫兹) 时间
载具(1 mL/kg) 2 第8天 三 2 第11天 10 2 第14天
100 第9天 三 100 第12天 10 100 第15天
10公里 第10天 三 10公里 第13天 10 10公里 第16天
将Nor-BNI实验中的大鼠分为三组:第1组-幼稚(no-SNI, n个 = 10); 第2组-对照组(SNI,no-SCS, n个 =11),组3–SCS(SNI,SCS, n个 = 10). 为了避免偏差,频率序列的安排方式是,3只大鼠接受SCS,顺序为2、100 Hz和10 kHz(表 4 ); 3只100Hz、10kHz和2Hz的大鼠; 4只大鼠,频率分别为10 kHz、2 Hz和100 Hz。
表4。
第1阶段 第2阶段 第3阶段
敌手 SCS(赫兹) 时间 敌手 Nor-BNI(μg) SCS(赫兹) 时间 敌手 Nor-BNI(μg) SCS(赫兹) 时间
车辆(10μL) 2 第8天 5.74 2 第11天 22.96 2 第14天
100 第9天 5.74 100 第12天 22.96 100 第15天
10公里 第10天 5.74 10公里 第13天 22.96 10公里 第16天
酶联免疫吸附试验(ELISA) 麻醉下SCS 30分钟后立即从延髓池收集脑脊液(CSF)。 蛋氨酸脑啡肽(Met-enk)的浓度是按照商业ELISA试剂盒(MBS756126,MyBiosource,Inc.,San Diego,USA)的说明进行测定的。
统计 使用IBM SPSS Statistics 17.0(美国伊利诺伊州芝加哥SPSS Inc.)和GraphPad Prism 7.0(美国加利福尼亚州圣地亚哥GraphPad-Software Inc.)进行统计分析和绘图。 为了进行比较,使用重复测量方差分析和Dunnett的多重比较。 数据的分布通过Shapiro-Wilk正态性检验进行检验。 非正态分布数据通过非参数检验(Kruskal-Wallis检验和Dunn的多重比较检验)进行分析。 对于所有数据,结果表示为平均值±SEM,以及 P(P) <0.05(双尾)被认为具有统计学意义。
结果 SCS运动阈值的确定 通过以2 Hz频率和200μs脉冲宽度从0 mA逐渐增加刺激强度来评估每只大鼠的MT。 实验条件下大鼠的平均MT在0.12至0.17 mA之间(图。 三 B) ●●●●。
SCS镇痛与机械过敏的严重程度 以前的研究结果表明,SCS的镇痛作用受模型机械过敏严重程度的影响[ 23 , 24 ]. 在重度和中度机械性超敏反应动物中检测SCS的镇痛作用:中度机械性过敏大鼠的50%PWT从3.42±0.72 g显著增加到9.79±3.56 g( P(P) <0.01),在15分钟SCS期间(图。 三 C1)。 50%的PWT几乎恢复到手术前水平。 同时,严重机械性超敏反应大鼠的50%PWT仅从前SCS水平0.55±0.10 g增加到1.89±1.10 g( P(P) >0.05)(图。 三 C1)。 为了比较SCS对重度和中度机械性超敏反应动物的镇痛作用,测量SCS期间50%的PWT(Dur-SCS); 这表示SCS启动后15 min(15 min-SCS)和30 min(30 min-SCS。 数据表明,中度机械过敏大鼠(8.85±2.77 g)的Dur-SCS 50%PWT比Pre-SCS水平(3.42±0.72 g)显著增加( P(P) <0.01,图。 三 C2),表明疼痛阈值增加。 中度机械性超敏大鼠的Dur-SCS也显著高于重度机械性超敏感大鼠(1.20±0.53 g, P(P) <0.01,图。 三 C2),表明SCS对中度机械性过敏大鼠具有更好的镇痛作用。
刺激强度-镇痛反应关系 采用20%MT至80%MT的不同刺激强度研究刺激强度与镇痛效果之间的关系。结果显示,除了40%MT外,刺激强度与止痛效果之间存在强度-反应关系(图。 4 A1、A2和表S1)。
图4。
不同强度和频率的SCS对短期SCS的镇痛作用,以及4种频率持续延长SCS的作用。 A1、A2 在50 Hz的SCS不同MTs下镇痛作用的时间过程。 中度机械性超敏大鼠的50%PWT呈强度依赖性和可逆性升高(另见表S1)。 看来60%MT或以上的SCS需要具有可靠的镇痛效果。 地下一层、地下二层 SCS在80%MT的不同频率下镇痛作用的时间过程。SCS的所有频率(2、15、50、100 Hz、10 kHz和2/100 Hz交替频率)在中度机械过敏大鼠中显示出相似程度的镇痛作用。 对于短期SCS,Pre-SCS表示0 min时的50%PWT 第1页 和 地下一层 ; 在SCS期间,15分钟和30分钟的平均PWT 地下一层 ; SCS后,60 min时50%PWT 第1页 和 地下一层 . C1、C2 在四种频率下延长80%MT SCS期间和之后50%PWT变化的时间过程。 在2、50、100 Hz和10 kHz下延长SCS达24小时,具有类似的持续镇痛效果。 除50Hz组外,SCS终止后1h镇痛效果消失。 SCS前,时间0 in时50%PWT C1类 ; 在SCS期间,从2小时到24小时,平均每2小时50%的PWT C1类 ; SCS后,25小时内50%PWT C1类 用Dunnett的多重比较检验进行重复方差分析** P(P) < 0.01, *** P(P) < 0.001 与 预-SCS; # P(P) < 0.05, ## P(P) < 0.01, ### P(P) < 0.001 与 No-SCS组中的Dur-SCS。
无刺激频率-镇痛反应关系 我们以前的研究表明,EA的镇痛作用是频率依赖性的[ 15 , 25 , 26 ]. 因此,我们检测了SCS在6种不同频率(2-、15-、50-、100-Hz SCS、10-kHz SCS和2/100-Hz交替SCS)下的镇痛效果。 与No-SCS组自身的Pre-SCS值或Dur-SCS值相比,SCS后各频率组的50%PWT都有类似的增加(图。 4 B1和B2)。 这些频率组之间的镇痛效果没有显著差异(表S2)。
持续SCS诱导的持续镇痛 为了在临床情况下模拟SCS的长期治疗,在连续24小时刺激期间评估SCS在四个选定频率(2、50、100 Hz和10 kHz)下的镇痛效果。 在整个24小时治疗期间,每隔一小时收集50%的PWT数据。 在24小时内,SCS在所有四种频率下都有类似的延长镇痛作用(图。 4 C1、C2; 2-Hz Pre-SCS:3.78±0.75 g,Dur–SCS:8.21±0.96 g; 50-Hz Pre-SCS:3.63±0.31克,Dur-SCS:9.21±0.79克; 100-Hz Pre-SCS:3.21±0.47克,Dur–SCS:8.56±0.85克; 10-kHz预-SCS:3.02±0.48 g,Dur-SCS:9.10±0.60 g)。 在任何频率组之间均未发现显著差异(表S3)。 在连续刺激的24小时内,镇痛效果持续,没有表现出任何耐受迹象。 测定了24小时SCS的后效。 SCS终止后60分钟,所有频率组的50%PWT值恢复到预刺激基线水平,50Hz组除外(图。 4 C1、C2和表S3)。 这与电针镇痛形成鲜明对比,电针镇痛在长时间刺激>6小时后出现耐受性,表明两种干预措施之间的潜在镇痛机制存在重大差异。
阿片受体参与SCS的镇痛作用 2-Hz SCS后50%PWT的增加被MOR特异性拮抗剂CTOP和KOR特异性拮抗剂Nor BNI阻断,但不被DOR特异性拮抗剂NTI阻断(图。 5 ,表S4和S5),表明MOR和KOR参与调节2-Hz SCS的镇痛作用。
图5。
阿片受体拮抗剂对2-Hz SCS镇痛作用的影响。 A1、A2 MOR特异性拮抗剂CTOP(3和10μg)显著减弱镇痛作用。 地下一层、地下二层 NTI(3和10 mg/kg)是一种DOR特异性拮抗剂,不影响镇痛效果。 C1、C2 KOR特异性拮抗剂Nor–BNI(5.74和22.96μg)显著降低镇痛效果。 SCS前,0分钟时50%PWT 第1页 , 地下一层 、和 C1类 ; 在SCS期间,15分钟和30分钟的平均PWT 第1页 , 地下一层 、和 C1类 ; SCS后,60 min时50%PWT 第1页 , 地下一层 、和 C1类 用Dunnett的多重比较检验进行重复方差分析* P(P) < 0.05, ** P(P) <0.01, *** P(P) < 0.001 与 预-SCS; # P(P) < 0.05, ## P(P) < 0.01, ### P(P) < 0.001 与 SCS+Veh组中的Dur-SCS。 CTOP,D-Phe-Cys-Tyr-D-Trp-On-Thr-Pen-Thr-NH 2 ; NTI,纳曲多尔; 去甲肾上腺素、去甲肾上腺素。
KOR特异性拮抗剂Nor-BNI阻止了100Hz SCS下50%PWT的增加,但CTOP或NTI没有阻止(图。 6 ,表S6和S7)。 这些结果表明,KOR参与了100 Hz SCS镇痛效应的介导。
图6。
阿片受体拮抗剂对100Hz SCS镇痛作用的影响。 A1、A2 CTOP(10和30μg)是一种MOR特异性拮抗剂,不影响镇痛效果。 地下一层、地下二层 NTI(3和10 mg/kg)是一种DOR特异性拮抗剂,不影响镇痛效果。 C1、C2 KOR特异性拮抗剂Nor-BNI(5.74和22.96μg)显著降低镇痛效果。 预-SCS,0 min时50%PWT 第1页 , 地下一层 、和 C1类 ; 在SCS期间,15分钟和30分钟的平均PWT 第1页 , 地下一层 、和 C1类 ; SCS后,60 min时50%PWT 第1页 , 地下一层 、和 C1类 用Dunnett的多重比较检验进行重复方差分析** P(P) <0.01*** P(P) < 0.001 与 预-SCS; # P(P) < 0.05, ### P(P) < 0.001 与 SCS+Veh组中的Dur-SCS。
10-kHz SCS的镇痛作用被测试的三种受体拮抗剂Nor-BNI、CTOP和NTI中的任何一种阻断(图。 7 ,表S8和S9),表明MOR、DOR和KOR均参与调节10-kHz SCS的镇痛作用。
图7。
阿片受体拮抗剂对10kHz SCS镇痛作用的影响。 A1、A2 MOR特异性拮抗剂CTOP(10和30μg)显著降低镇痛效果。 地下一层、地下二层 NTI(3和10 mg/kg)是一种DOR特异性拮抗剂,可显著抵消超敏反应抑制作用。 C1、C2 KOR特异性拮抗剂Nor-BNI(5.74和22.96μg)显著降低镇痛效果。 预-SCS,0 min时50%PWT 第1页 , 地下一层 、和 C1类 ; 在SCS期间,15分钟和30分钟的平均PWT 第1页 , 地下一层 、和 C1类 ; SCS后,60 min时50%PWT 第1页 , 地下一层 、和 C1类 使用Dunnett多重比较检验的重复方差分析** P(P) < 0.01, *** P(P) < 0.001 与 预-SCS; # P(P) < 0.05, ## P(P) < 0.01, ### P(P) < 0.001 与 SCS+Veh组中的Dur-SCS。
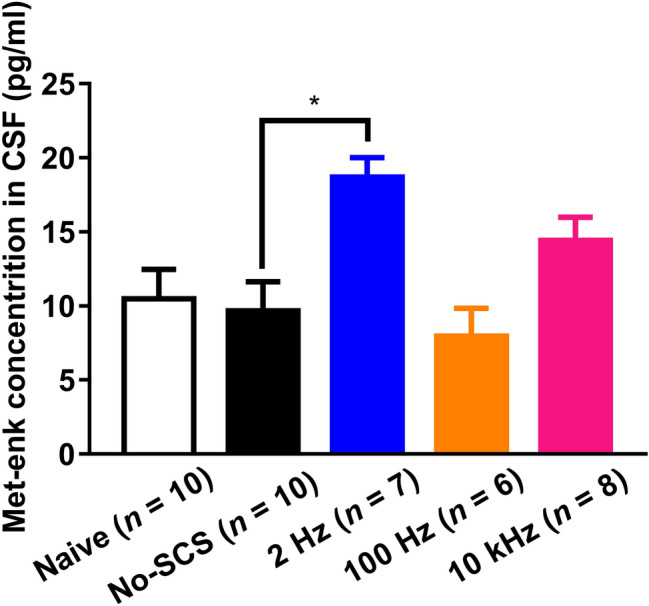
CSF中的Met-enk含量 除了阿片肽和阿片受体参与的药理证据外,SCS后CSF中Met-enk含量的变化也被测定。 结果表明,2Hz SCS后,脑脊液中Met-enk的浓度从No-SCS组的9.68±1.96 pg/mL显著增加到18.70±1.30 pg/mL。 10-kHz SCS后Met-enk含量有增加的趋势(14.43±4.41 pg/mL),但与No-SCS组相比,未达到统计显著水平。 100Hz SCS后Met-enk的含量保持不变(7.95±1.89 pg/mL,图。 8 ).
图8。
三种频率SCS后大鼠脑脊液中的Met-enk含量(Kruskal-Wallis和Dunn的多重比较试验* P(P) < 0.05). 蛋氨酸脑蛋白; 脑脊液。
讨论 SCS作为一种重要的神经调节技术,近年来在慢性疼痛治疗中得到了越来越广泛的应用。 镇痛作用的潜在机制尚未完全了解,目前正在努力提高镇痛效果。 利用中度机械性超敏SNI大鼠模型,我们表征了SCS的镇痛作用,并研究了其作用机制。 特别是,我们对是否涉及内啡肽和阿片受体以及脉冲频率的选择是否对治疗效果至关重要感兴趣。
SCS的镇痛作用和刺激参数 SCS的镇痛效果可能受到许多因素的影响,例如受刺激的脊髓节段、神经病理性疼痛的严重程度以及波形、脉宽、强度和频率等刺激参数。
据报道,SCS用于含有受损脊神经或以上的脊髓节段时,其镇痛效果更为有效[ 23 , 24 ]这与SCS的镇痛作用是基于门控理论的假设是一致的。 由于SNI大鼠有来自L4、L5和L6的脊髓神经损伤,因此我们选择对应于脊髓L4、M5和M6的T13–L1椎体节段作为刺激部位。 SCS诱导镇痛的门控假说也得到了以下事实的支持:持续24小时的SCS刺激后没有产生耐受性。据报道,持续6小时电针大鼠的镇痛效果显著降低[ 17 ]. 人们认为中枢内啡肽在介导电针的镇痛作用中起着重要作用[ 15 ].
在SNI模型中比较了SCS在不同程度机械性超敏反应(中度)时的镇痛作用 与 严重)。 我们的数据表明,SCS对中度机械性过敏大鼠的镇痛更有效。 这与之前报告的结果一致[ 23 , 24 ].
我们研究了SCS在不同强度(对应20%、40%、60%和80%MT)下的镇痛效果。 可以预测,在运动阈值内,刺激强度与脊髓刺激的镇痛作用呈正相关(图。 4 ),与以前的报告一致[ 27 – 29 ]. 选择80%MT对应的刺激强度来研究SCS的机制,并在神经病理性疼痛大鼠模型中产生显著的镇痛作用。
SCS的频率可以作为向中枢神经系统传递神经调制信息的独立信号。 已经证明,电针或TEAS刺激周围神经以频率依赖的方式缓解疼痛[ 15 – 17 ]. 例如,2-Hz TEAS在减少神经病理性疼痛方面比100Hz更有效[ 30 ]. 相比之下,100-Hz TEAS对由上部神经元损伤引起的肌肉痉挛引起的疼痛更有效[ 31 ]. 我们假设SCS诱导的镇痛也具有频率特异性,因为用于外周神经刺激和SCS的脉冲几乎相同,除了这两种方式中使用的刺激强度不同。 此外,初级传入神经在介导电针镇痛中起着关键作用。 脊髓刺激也可以激活这些神经,电极位于脊髓背根和背角附近。 然而,与之前的一些发现类似[ 28 , 31 ]我们的数据显示,不同频率(2、15、50、100、2/100 Hz和10 kHz)的SCS具有相似的镇痛程度。 这一发现与一些临床报告不一致,其中10-kHz SCS比传统SCS更有效[ 32 ]. 一种可能性是患者可能无法耐受动物研究中使用的高刺激强度(80%MT)。 较低频率和较高强度的SCS会产生不适感。 相比之下,10-kHz刺激每单位时间提供更多能量,但不会产生不舒服的感觉异常。 因此,患者在较高强度下更容易耐受。 据报道,一种新的刺激模式,即差分目标复合编程SCS,比传统SCS更有效[ 33 – 36 ]. 不幸的是,这些刺激模式的神经化学机制尚未完全理解,也未纳入本研究。
本研究结果表明,不同频率的SCS在长时间刺激后具有相似的抗伤害效果和相似的持续效果。 值得注意的是,本研究仅使用了一种神经病理性疼痛的SNI模型。 在得出SCS在缓解疼痛方面的频率特异性的任何结论之前,还应该研究各种疼痛的其他模型。
阿片受体与SCS 镇痛机制:SCS与EA 我们以前的研究表明内源性阿片肽及其受体在电针镇痛中起着关键作用[ 15 – 17 ]. 2 Hz电针可增加中枢神经系统中脑啡肽的释放,并与MOR和DOR结合以诱导镇痛作用。 另一方面,100 Hz的电针增加强啡肽的释放并激活KOR[ 15 – 17 ].
在本研究中,2 Hz的SCS增加了大鼠脑脊液中Met-enk的浓度,并且MOR特异性拮抗剂阻断了其镇痛作用。 这些数据表明,与EA类似,SCS在2Hz时的镇痛作用是由Met-enk和MOR介导的。
KOR选择性拮抗剂也显著减弱了2-Hz SCS的镇痛作用,表明KOR可能也参与了2-Hz CS诱导的镇痛作用。
100Hz SCS的镇痛作用被KOR拮抗剂完全阻止。 此外,在脑脊液中未发现Met-enk浓度的变化,并且MOR或DOR拮抗剂未阻断镇痛作用,表明100-Hz SCS由KOR介导。 100Hz脊髓刺激系统可能具有类似于100Hz电针的神经化学机制:它选择性地刺激脊髓释放强啡肽,激活KOR以产生镇痛作用。 值得注意的是,在本研究中,脑脊液中Met-enk浓度的差异很大,在未来的研究中,更多的动物将能够增加统计分析的能力。
佐藤及其同事发现60Hz SCS的镇痛作用是由DOR介导的[ 13 ]. 然而,在我们的研究中,在DOR拮抗剂存在的情况下,100-Hz SCS的镇痛效果稍低,但没有达到统计学意义。 差异可能是因为他们的研究中使用了渗透微型泵持续给药,而我们的研究中则使用了静脉注射。
结果表明,SCS和EA的镇痛机制既有相似之处,也有差异。 产生差异的主要原因可能是EA信号仅由原发性影响神经纤维介导,而SCS不仅激活感觉神经元,还激活脊髓中的中间神经元。 后者可能会有多种反应。
值得注意的是,我们的结果与之前报道的结果一致:低频(2或4 Hz)SCS激活MOR,高频(50、60或100 Hz)SCS-激活KOR[ 13 ],我们的结果表明低频SCS也激活KOR。
10千赫SCS SCS在10 kHz时的镇痛作用被三种阿片受体拮抗剂(MOR、DOR和KOR)阻断。 我们最近在大鼠身上使用单单位记录技术的研究表明,EA-like外周电刺激的信号通过aβ和aδ初级传入感觉纤维传递到中枢神经系统。 初级感觉神经元记录的最高脉冲频率为~250 Hz,即使刺激频率高达1000 Hz。 这可能是由于神经元的不应期[ 37 ]. 因此,10kHz SCS的频率太高,无法传输到中枢神经系统神经元。 本研究的数据表明,100 Hz和10 kHz的SCS诱导了类似的伤害性反应减少。 可以合理假设脊髓神经元在10 kHz时SCS的实际或有效刺激频率可能不大于250 Hz。 另一种可能性是,10-kHz SCS可能通过使无髓C纤维或其他神经元保持不易反应,从而阻断伤害性信号的传输。
与长期电针刺激后出现的耐受性发展不同,在持续SCS 24小时内未发现镇痛效果下降的迹象。 这表明除内啡肽外的其他机制在SCS镇痛中发挥重要作用。
门控理论也可以用来解释SCS的镇痛作用。 强啡肽能神经元是抑制性中间神经元,在门控制机制中起关键作用。 它们接收来自初级传入Aβ纤维的信号并关闭门。 因此,疼痛信号无法进入CNS[ 38 ]. 2、100-Hz、10-kHz SCS和100-Hz EA的镇痛作用均被KOR拮抗剂阻断,表明这些频率的SCS激活抑制性强啡肽能中间神经元产生镇痛作用。
总之,在中度神经病理性疼痛的SNI模型中,SCS具有有效的抗伤害作用,且后效最小。 在我们的实验条件下,SCS的抗伤害作用具有强度依赖性和持续性,并且没有产生耐受性。 许多内啡肽及其受体可能参与调节这种效应。 SCS和EA的机制既有相似之处,也有不同之处。
确认 我们感谢中国北京品思医疗有限公司的技术援助。 特别感谢傅旭(北京品思医疗有限公司)及其同事为动物研究修改SCS设备。 这项工作得到了科技部国家重点研发计划(2016YFC01055012019YFD1712000)的支持。
参与者信息 张荣,电子邮箱: zhangrong@bjmu.edu.cn。
Ji-Sheng Han,电子邮件: hanjisheng@bjmu.edu.cn。
工具书类
1 Shealy CN、Mortimer JT、Reswick JB。 刺激背柱电抑制疼痛:初步临床报告。 Anesth分析。 1967; 46:489–491。 [ 公共医学 ] [ 谷歌学者 ]
2 Sdrulla AD,Guan Y,Raja SN。脊髓刺激:临床疗效和潜在机制。 疼痛练习。 2018; 18:1048–1067. doi:10.1111/papr.12692。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
三。 Melzack R,Wall PD。疼痛机制:一种新理论。 痛苦论坛。 1996; 5:3–11. doi:10.1016/S1082-3174(96)80062-6。 [ 内政部 ] [ 谷歌学者 ]
4 Guan Y.脊髓刺激:神经生理学和神经化学作用机制。 Curr Pain头痛报告2012; 16:217–225. doi:10.1007/s11916-012-0260-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
5 Tazawa T、Kamiya Y、Kobayashi A、Saeki K、Takiguchi M、Nakahashi Y等。脊髓刺激调节单侧脊髓神经损伤大鼠下行抗伤害系统的脊髓上中枢。 Mol疼痛。 2015; 11:36. doi:10.1186/s12990-015-0039-9。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
6 SaadéNE、Barchini J、Tchachaghian S、Chamaa F、Jabbur SJ、Song Z等。脊髓刺激缓解疼痛效果中背外侧索的作用:对神经性疼痛大鼠模型的研究。 2015年实验脑研究; 233:1041–1052. doi:10.1007/s00221-014-4180-x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
7 Song Z、Ansah OB、Meyerson BA、Pertovara A、Linderoth B。在神经病理性疼痛的啮齿动物模型中,头内侧髓质参与脊髓刺激的作用。 神经科学。 2013; 247:134–144. doi:10.1016/j.neuroscience.2013.05.027。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
8 Ultenius C,Song ZY,Lin PY,Meyerson BA,Linderath B.神经病理性疼痛啮齿类动物脊髓刺激效应中的脊髓GABA能机制:GABA合成是否参与? 神经调节。 2013; 16:114–120. doi:10.1111/ner.12007。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
9 Song ZY、Meyerson BA、Linderath B.脊髓5-HT受体对神经病变大鼠模型中脊髓刺激的镇痛作用有贡献。 疼痛。 2011; 152:1666–1673. doi:10.1016/j.pain.2011.03.012。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
10 Yang F,Xu Q,Shu B,Tiwari V,He SQ,Vera-Portcarrero LP,等。大麻素CB1受体的激活有助于通过Aβ纤维刺激抑制脊髓伤害性传递和机械性超敏反应。 疼痛。 2016; 157:2582–2593. doi:10.1097/j.pain.0000000000000680。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
11 Sun L、Tai L、Qiu Q、Mitchell R、Fleetwood-Walker S、Joosten EA等。CB1受体的内源性大麻素激活有助于通过重复脊髓刺激长期逆转神经病理性疼痛。 欧洲疼痛杂志。 2017; 21:804–814. doi:10.1002/ejp.983。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
12 Ding XH,Hua F,Sutherly K,Ardell JL,Williams CA。C2脊髓刺激诱导大鼠T4脊髓释放强啡肽:心肌缺血敏感神经元的电位调节。 美国生理学杂志Regul Integr Comp Physiol。 2008; 295:R1519–R1528。 doi:10.1152/ajpregu.00899.2007。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
13 佐藤·KL、国王·EW、约翰·LM、斯拉卡·KA。 脊髓刺激通过以频率依赖的方式激活阿片受体来降低超敏反应。 欧洲疼痛杂志。 2013; 17:551–561. doi:10.1002/j.1532-2149.2012.0022.x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
14 Tonelli L、Setti T、Falasca A、Martignoni E、Torcia E、Calcaterra FM等。与脊髓刺激相关的脑脊液阿片类物质和神经递质的研究。 应用神经生理学。 1988; 51:324–332. doi:10.1159/000099977。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
15 韩JS。 针刺:通过不同频率的电刺激产生的神经肽释放。 《神经科学趋势》。 2003; 26:17–22. doi:10.1016/S0166-2236(02)00006-1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
16 Ulett GA、Han S、Han JS。 电针:机制和临床应用。 生物精神病学。 1998; 44:129–138. doi:10.1016/S0006-3223(97)00394-6。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
17 韩JS,Terenius L.针刺镇痛的神经化学基础。 《药物毒理学年鉴》。 1982; 22:193–220. doi:10.1146/annurev.pa.22.040182.001205。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
18 装饰I,Woolf CJ。 备用神经损伤:持续性周围神经病理性疼痛的动物模型。 疼痛。 2000; 87:149–158. doi:10.1016/S0304-3959(00)00276-1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
19 狄克逊·WJ。 实验观察的有效分析。 《药物毒理学年鉴》。 1980; 20:441–462. doi:10.1146/annurev.pa.20.040180.002301。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
20 Chaplan SR、Bach FW、Pogrel JW、Chung JM、Yaksh TL。大鼠爪子触觉超敏的定量评估(Von Frey hairs)神经科学方法杂志。 1994; 53:55–63. doi:10.1016/0165-0270(94)90144-9。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
21 Ju J,Shin DJ,Na YC,Yoon MH.脊髓阿片受体在神经病变大鼠鞘内痛觉素抗超敏作用中的作用。 神经科学快报。 2013; 542:118–122。 doi:10.1016/j.neulet.2013.03.026。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
22 陈XH,韩建生。 脊髓中的三种阿片受体对2/15Hz电针镇痛都很重要。 欧洲药理学杂志。 1992年; 211:203–210. doi:10.1016/0014-2999(92)90530-H。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
23 Smits H、Ultenius C、Deumens R、Koopmans GC、Honig WMM、van Kleef M等。神经性疼痛动物模型中脊髓刺激的效果与触觉“超敏”程度有关。 神经科学。 2006; 143:541–546. doi:10.1016/j.neuroscience.2006.08.007。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
24 Smits H、van Kleef M、Joosten EA。 神经病理性疼痛大鼠模型中脊髓刺激背柱:节段性脊髓疼痛缓解机制的证据。 疼痛。 2012; 153:177–183. doi:10.1016/j.pain.2011.10.015。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
25 邢国光,刘凤英,瞿XX,韩建生,万毅。神经病理性疼痛大鼠脊髓背角的长期突触可塑性及其电针的调节。 实验神经学。 2007; 208:323–332。 doi:10.1016/j.expneuro.2007.09.004。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
26 姜毅,王毅,刘焕。 频率为2 Hz和100 Hz的经皮神经电刺激对大鼠慢性炎性疼痛的治疗效果比较。 中国中西医结合杂志。 2001; 21:923–925. [ 公共医学 ] [ 谷歌学者 ]
27 Shechter R、Yang F、Xu Q、Cheong YK、He SQ、Sdrulla A等。在神经性疼痛大鼠模型中,常规和千赫兹脊髓刺激可产生强度和频率依赖性的机械超敏反应抑制。 麻醉学。 2013; 119:422–432. doi:10.1097/ALN.0b013e31829bd9e2。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
28 Sato KL、Johanek LM、Sanada LS、Sluka KA。 脊髓刺激可减少神经病理性疼痛动物的机械性痛觉过敏和胶质细胞激活。 Anesth分析。 2014; 118:464–472. doi:10.1213/ANE.00000000000047。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
29 Meuwissen KPV、Gu JW、Zhang TC、Joosten EAJ。 在慢性神经病理性疼痛大鼠模型中,常规-SCS与突发-SCS以及对机械性超敏反应的行为影响:振幅的影响。 神经调节。 2018; 21:19–30. doi:10.1111/ner.12731。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
30 韩JS。 针灸镇痛:共识和争议领域。 疼痛。 2011; 152页:第41页至第48页。 doi:10.1016/j.pain.2010.10.012。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
31 赵伟林,王C,李振中,陈雷,李建军,崔维德,等。经皮穴位电刺激治疗脑损伤后肌肉痉挛的疗效和安全性:一项双盲、多中心、随机对照试验。 公共科学图书馆一号。 2015; 10:e0116976。 doi:10.1371/journal.pone.0116976。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
32 Song ZY、Viisanen H、Meyerson BA、Pertovara A、Linderath B。千赫频率和常规脊髓刺激对不同疼痛状态大鼠模型的疗效。 神经调节技术神经接口。 2014; 17:226–235. doi:10.1111/ner.12161。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
33 Kapural L,Yu C,Doust MW,Gliner BE,Vallejo R,Sitzman BT等。10-kHz高频和传统低频脊髓刺激治疗慢性腰腿痛的比较:一项多中心、随机、对照关键试验的24个月结果。 神经外科。 2016; 79:667–677. doi:10.1227/NEU.0000000001418。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
34 Vallejo R、Kelley CA、Gupta A、Smith WJ、Vallejo A、Cedeño DL。 在神经病理性疼痛动物模型中使用差异靶点复合脊髓刺激调节神经胶质相互作用。 Mol疼痛。 2020; 16:1744806920918057. doi:10.1177/1744806920918057。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
35 Fishman M、Cordner H、Justiz R、Provenzano D、Merrell C、Shah B等。对慢性顽固性背痛和腿痛受试者进行差异靶点多功能脊髓刺激和传统脊髓刺激的多中心、开放标签、随机对照临床试验的12个月结果。 疼痛练习。 2021; 21:912–923. doi:10.1111/papr.13066。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
36 Gong WY、Johanek LM、Sluka KA。 在神经病理性疼痛动物模型中,比较脊髓爆裂和张力刺激对痛觉过敏和体力活动的影响。 Anesth分析。 2016; 122:1178–1185。 doi:10.1213/ANE.000000000001161。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
37 霍润,韩世平,刘凤英,寿XJ,刘丽丽,宋天杰,等。初级传入纤维对不同频率针刺样外周刺激的反应:大鼠单单位记录的表征。 神经科学公牛。 2020; 36:907–918. doi:10.1007/s12264-020-00509-3。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
38 段斌,程丽珍,马奇芬。 脊髓回路传递机械性疼痛和瘙痒。 神经科学公牛。 2018; 34:186–193. doi:10.1007/s12264-017-0136-z。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
关联数据 本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。