1.简介 骨质疏松症是一种全身性骨骼疾病,其特点是骨量减少,骨强度减弱,导致骨脆性,并在骨吸收超过新骨形成时发生。
1 大约30%的女性和12%的男性在一生中的某个时候受到影响,尤其是在老年人和绝经后女性中。
2 骨质疏松性骨折的后果可能很严重,包括残疾、依赖性降低甚至死亡。 骨质疏松症已成为世界范围内的一种主要流行疾病,极大地增加了医疗负担。
三 骨质疏松症治疗的目的是恢复骨内稳态并保持正常的骨量。尽管市场上有许多可用的骨质疏松症药物,但人们对长期使用这些药物的固有副作用感到担忧,促使人们探索新的治疗靶点。
4 然而,骨质疏松症的病理生理学是多因素的,包括成骨细胞和破骨细胞之间的失衡,微结构破坏,骨髓中脂肪生成增加,以及血管生成、氧化应激和遗传/表观遗传因素的变化。
5 最近的证据表明,骨髓间充质干细胞(BMSCs)的异常分化是骨质疏松症发生的主要因素之一,因此可能成为骨质疏松症治疗的潜在靶点。
6
长非编码RNA(lncRNA)是一类含有200个或更多核苷酸的转录物,其蛋白质编码能力较低或没有。
7 可以想象,lncRNAs在转录和转录后水平的基因表达中发挥着重要作用,并调节各种生物功能和病理意义,包括骨骼的功能和病理含义。
8 事实上,越来越多的证据表明,各种lncRNAs参与骨内稳态。 例如,Huang等人。
9 结果表明,过度表达的lncRNA H19促进了BMSCs的成骨细胞分化并增加了异位骨形成,而lncRNA H29的敲除则消除了这些影响。 已发现LncRNA LINC00311可诱导骨质疏松大鼠破骨细胞增殖并抑制破骨细胞凋亡。
10 最近的一项研究还发现,lncRNA SNHG1的表达可以抑制BMSCs的成骨分化。
11
非编码RNA序列,包括lncRNAs,通过与RNA结合蛋白(RBP)的相互作用,在许多细胞过程中发挥着不同的功能,其中RBP在转录后基因调控中发挥着多种作用。
12 RBP是通过影响mRNA代谢参与基因表达的共转录或转录后调节的转录因子。
11 聚嘧啶束结合蛋白1(PTBP1)是调节mRNA衰变稳定性和前mRNA剪接的RBP之一。
13 除此之外,PTBP1还参与BMSC诱导的低恶性癌细胞的侵袭
14 并与人类脂肪来源MSC向最终内胚层分化有关。
15 迄今为止,PTBP1在BMSCs分化中的潜在作用尚未报道。
DNA甲基转移酶(DNMT)1是表观遗传修饰的积极参与者,通过维持细胞分裂期间的DNA甲基化,负责基因组甲基化模式的被动传递。
16 骨保护素(Opg)是BMSC分化和脂肪生成过程中的重要调节因子。
17 作为DNMT1的潜在靶点和破骨细胞分化和作用的关键调节因子之一,Opg通过竞争性抑制Opg配体和RANK在破骨细胞及其前体上的相互作用,发挥核因子κ-B(RANK)信号通路受体激活剂的抑制剂的作用。
18 原发性骨质疏松症患者也发现Opg的高甲基化。
19 此外,以前的研究表明,lncRNA SNHG1可以结合并调节DNMT1在某些疾病(如癌症)中的表达。
20 ,
21 因此,我们假设lncRNA SNHG1可以调节DNMT1的表达,进而影响Opg甲基化状态,从而促进骨质疏松症的发生,但需要进一步确定。
本研究的目的是阐明lncRNA SNHG1在骨质疏松症发病机制中的功能作用以及潜在的机制途径。
2.材料和方法 2.1. 动物 所有动物均按照《实验动物护理和使用指南》进行饲养和处理。 所有动物研究均由宁波大学实验动物伦理委员会批准(2020‐106)。 如前所述,建立了雌激素缺乏性骨质疏松小鼠模型。
22 11–12周龄时体重为20–25 g的雌性C57BL/6小鼠在标准条件下进行12小时光/暗循环。 温度和湿度分别控制在20–25°C和40%–70%。 老鼠可以自由享用标准的食物和水。 在随机化之前,所有动物均已适应环境至少5天。 随后,小鼠随机接受双侧卵巢切除术(OVX)或假手术(sham)。 8周后,安乐死后收集小鼠股骨,用多聚甲醛固定48小时。 采集血样并采集骨组织。 然后立即将样品冷冻在液氮中,并将其储存在−80°C下,以备进一步实验。
2.2. 临床组织采集 在本研究中,从骨质疏松患者和非骨质疏松患者(对照组)中采集了六对骨组织,并在我院进行了常规治疗手术,并立即储存在−80°C下,以供后续分析。 实验方案由中国科学院大学华美医院伦理委员会(PJ‐NBEY‐KY‐2019‐081‐01)审查和批准,本研究中的所有方法均符合批准的指南。 所有参与研究的患者在进行任何研究程序之前均出具了书面知情同意书。
2.3. 细胞培养和治疗 小鼠骨髓间充质干细胞(MUBMX‐01001)从Cyagen Inc.获得,并在37°C、5%CO下培养 2 在含有10%胎牛血清(Gibco)、2 mmol/l l-谷氨酰胺、100 U/ml青霉素和100μg/ml链霉素的Alpha-MEM培养基中。 然后用成骨分化诱导培养基(通过向生长培养基中添加10 mMβ-甘油磷酸钠、0.1μM地塞米松和50 mg/ml抗坏血酸配制)处理BMSC 或脂肪分化诱导培养基(通过向高糖MEM培养基中添加1%双抗体、10 mM 3-异丁基-1-甲基黄嘌呤、10 mM-吲哚美辛、10 nM地塞米松配制)分别诱导成骨或成脂分化。
23 采用茜素红S(ARS)染色和油红O(ORO)染色检测相应诱导培养基培养后的成骨和成脂分化。 用生长培养基处理的骨髓间充质干细胞作为对照。 诱导后,收集细胞进行进一步分析。 对于5‐Aza‐2′-脱氧胞苷(5‐Aza)处理,用10µM 5‐Za(Sigma)处理细胞96小时,每24小时更换一次培养基。
2.4. 质粒构建和转染 通过PCR扩增全长lncRNA SNHG1序列,然后将其插入pcDNA 3.1载体(Life Technologies)中,以根据制造商的说明建立过度表达SNHG1的载体(pcDNA‐SNHF1)。 空载体被用作阴性对照(NC)。 使用与上述相同的方法,克隆了PTBP1和Opg,并分别命名为pcDNA-PTBP1与pcDNA-Opg。 针对lncRNA SNHG1、PTBP1、Opg和sh阴性对照(sh‐NC)的短发夹(sh)RNA来自GenePharma。 使用Lipofectamine 3000(Life Technologies)进行细胞转染。 简言之,在α-MEM培养基中生长至70%融合的骨髓间充质干细胞在成脂或成骨诱导培养基中培养之前,用Lipofectamine 3000试剂和pcDNA或sh‐RNA培养48小时。 对于体内转染,通过尾静脉将lent‐sh‐SNHG1和/或sh‐Opg注入循环。 连续注射3周后,小鼠最终被安乐死。
2.5. 血红素和曙红染色 将小鼠股骨固定在磷酸盐缓冲的10%多聚甲醛中24小时,然后在4°C下用8%甲酸在连续摇晃下脱钙。 然后将脱水组织包埋在石蜡中,并切成5µm的切片,用于苏木精和伊红(HE)染色。 按照常规方案进行血红素和曙红染色。 简言之,在脱蜡和再水化后,用苏木精溶液对5μm纵切面染色5分钟,然后在1%酸性乙醇(70%乙醇中的1%HCl)中浸泡5次,然后在蒸馏水中冲洗。 然后用曙红溶液对切片染色3分钟,然后用分级酒精脱水并在二甲苯中清除。 随机选择至少5个区域,并在光学显微镜(尼康)下进行拍照,以进行组织学评估。
2.6. ORO和ARS染色 如前所述,在BMSCs成骨和成脂分化后14天进行油红O染色和ARS染色。
23 丢弃细胞培养基,用磷酸盐缓冲盐水(PBS;Solarbio)冲洗细胞三次。 将细胞在室温下(RT)的4%多聚甲醛中固定20分钟,然后用ORO(Cyagen Biosciences)或ARS(Cyangen Bioscinces)染色30分钟,然后根据制造商的说明用去离子水清洗三次。 洗涤后,用相差显微镜对细胞的脂滴进行拍照。 为了定量矿化,ARS染色用100 mmol/l十六烷基氯化吡啶(Sigma)溶解。 培养1 h后,在570 nm处用分光光度法测量吸光度。
2.7. 免疫荧光染色 免疫荧光(IF)染色如下。 细胞用PBS清洗,并在室温下用4%多聚甲醛固定20分钟。 然后,用PBS冲洗细胞三次,并在室温下在0.3%吐温-100(Biosharp)中渗透1h。 在用适当体积的可用山羊血清(Boster Biological Technology)封闭30分钟并清洗三次后,样品与初级抗-Pparγ抗体(ab59256,稀释1:200;Abcam)在4°C下孵育过夜。 样品用PBS洗涤三次,然后用荧光素标记的相应二级抗体在RT下孵育1小时。 用4,6-二氨基-2-苯基吲哚盐酸盐(DAPI)(D8417;Sigma)对细胞核进行复染。 用IX73显微镜(Olympus)拍摄照片,以证明脂肪分化。
2.8. 碱性磷酸酶(ALP)染色和活性 碱性磷酸酶染色在BMSCs成骨分化后第14天进行。
24 简而言之,用PBS清洗细胞,并用4%多聚甲醛固定10分钟,然后在碱性磷酸酶溶液(CWBIO)中培养,在RT避光染色20分钟。三次清洗后,用倒置光学显微镜获得钙矿化图像。 在与10 mM对硝基苯基磷酸酯(Sigma)孵育15分钟后,通过微孔板读数器在420 nm处计算ALP活性。
2.9. 荧光原位杂交(FISH)分析 生物素标记的正、反义SNHG1探针和相应的FISH分析对照寡聚体是从GenePharma获得的,FISH检测如前所述。
25 为了固定化,细胞在室温下用4%多聚甲醛处理20分钟。用PBS洗涤后,用杂交溶液预杂交细胞。 然后,将细胞与SNHG1探针在37°C下孵育过夜。 使用抗生物素Cy3抗体(C5585;Sigma)进行信号检测。 用DAPI对细胞核进行复染,以评估细胞核形态。 将样品与浓度在0.5µg/ml之间的稀释DAPI溶液在PBS中在黑暗中RT培养5分钟。使用IX73显微镜(Olympus)测量免疫荧光。
2.10条。 RNA下拉分析 如前所述,进行生物素化RNA下拉分析,以确定lncRNA SNHG1和PTBP1之间的相互作用。
26 试验中使用的生物素标记的RNA是通过生物素RNA标记混合物(罗氏)和T7 RNA聚合酶的体外转录合成的。 简而言之,细胞在免疫沉淀(IP)缓冲液(20 mM Tris‐HCl,pH 8.0,200 mM NaCl,1 mM EDTA,1 mM-EGTA,0.5%Triton X‐100,0.4 U/μl RNasin)中溶解,最终细胞浓度为5×10 6 细胞/ml。通过将全细胞裂解物与生物素化RNA在RT下孵育30分钟来进行检测。然后用链霉亲和素磁珠分离RNA蛋白复合物(Thermo Scientific)。 RNA-蛋白复合物与链霉亲和素磁珠在滚筒上RT培养30分钟。 磁珠在磁性支架中分离,样品在37°C的洗脱缓冲液中在摇床上培养30分钟后洗脱。 使用十二烷基硫酸聚丙烯酰胺凝胶电泳(SDS‐PAGE)分离沉淀组分,并通过western blot进行可视化。
2.11. RNA免疫沉淀分析 根据制造商的说明,使用商用EZ‐Magna RIP™试剂盒(Millipore)进行RNA免疫沉淀(RIP)分析。 简而言之,细胞(5×10 6 )在4°C下用RIP裂解缓冲液处理30 min,然后用RIP缓冲液培养全细胞裂解产物,该缓冲液含有与Ago2(Millipore)或抗PTBP1抗体(ab133734;Abcam)或抗SNRP70抗体(ab51266;Abcan)或免疫球蛋白(Ig)G(Abcam。 用Trizol试剂(Life Technologies)提取与PTBP1相关的共沉淀RNA,并通过定量实时PCR(qPCR)进行分析。 与SNRP70和物种匹配的正常IgG相关的富集分别作为RIP阳性和阴性对照。 总RNA被视为输入对照。
2.12. qPCR检测 动物骨组织取自OVX和假小鼠。 从被诊断为骨质疏松症的患者和对照患者身上采集人体骨骼样本。 使用Trizol(生命技术)方法从骨髓间充质干细胞和骨组织中提取总RNA。 细胞或组织在Trizol中均质并用氯仿(0.2 ml/ml Trizol)萃取。 使用异丙醇(0.5 ml/ml Trizol)从其水相中沉淀RNA。 在7500℃下通过离心法造粒RNA 克 和4°C保持5分钟,在75%乙醇中洗涤,再次离心,风干10分钟,并溶解在200μl HPLC水中。 使用NanoDrop分光光度计(Thermo Scientific)测量RNA浓度,使用安捷伦生物分析仪RNA 6000 Nano试剂盒(安捷伦科技)测定RNA质量。 然后使用iScript™cDNA合成试剂盒(Bio-Rad)将提取的RNA反向转录为第一链cDNA。 qPCR分析是使用UltraSYBR混合物试剂盒(Thermo Scientific)在Applied Biosystems 7500 Fast Dx Real‐Time PCR系统(Life Technologies)上进行的。 qPCR检测中使用的引物如下:人SNHG1(正向:CCACCTCTGTTCCCGTCAT,反向:GACAGCAGTCAAGGGA); 鼠标SNHG1(正向:TCCTTGTTCGGGGTTTGAGG,反向:ACAGCACCTGACTAAGC); Opg(前进:GAGTGTGAGGAGGAAGCGGTTA,后退:GTGCTGCAGTTCGTGTT); Runt相关转录因子2(Runx2)(正向:AGATGGACTGTGGTTACCG,反向:GGACCTCTGCACTT); ALP(正向:CATTCCCATGTCTTCACCTTTG,反向:TCCTCTTGTTCACATG); 骨形态发生蛋白4(Bmp4)(正向:CATTCCCATGTTCTTCACCTTG,反向:TCCTCTTGTTCCGTTCACATG); 骨γ-羧基谷氨酸蛋白(Bglap)(正向:CATTCCCATGTTCTTCACCTTTG,反向:TCCTCTTTCCGTTCACATG); 过氧化物酶体增殖物激活受体γ(Ppar‐γ)(正向:AAGAGCTGACCCATGGTTG,反向:ACCCTTCACAAG); 脂肪酸结合蛋白4(Fabp4)(正向:CGTAAATGGGGATTGGTCA,反向:TCGACTTTCCACTCTC); CCAAT增强子结合蛋白α(Cebpa)(正向:GTGGACAGAGAGCAACGA,反向:CCTTGACCAAGAGCTCTCA); PTBP1(正向:CAGAGGACGACCTCAAGAGGC,反向:GGTGGACTTGGAAAGACA); DMNT1(正向:GATCACATTTTGGCTGGTGTCT,反向:CATGGCATTCCTTGTCCT); 胶原蛋白1型α1(Col1a1)(正向:TGACTGGAGAGCGGAGT,反向:GTTCGGGCTGATGTACCAGT); 人类GAPDH(正向:CCAGGTGTCTCCTCTGA,反向:GCTGTAGCCAAATCGT); 小鼠GAPDH(正向:AGCCCAAGATGCCCTCAGT,反向:CCGTGTTCCTACCTCATG)。 GAPDH作为内部参考。 在内部验证的实验设置期间,与未转染细胞相比,用空pcDNA 3.1载体或sh‐NC转染的细胞中上述靶基因的表达没有改变。 因此,分析基因表达时,空的pcDNA 3.1载体和sh‐NC将作为后续转染研究的对照。 通过应用相对定量2计算目标基因的相对表达值 -∆∆Ct 测量折叠变化的方法。 所有测量重复三次。
2.13. 定量甲基化特异性PCR 定量甲基化特异性PCR(MSP)旨在特异性扩增亚硫酸氢盐转化的DNA靶点。
27 通过靶向Opg区域内富含CpG的区域,Opg的引物可以特异识别亚硫酸氢盐处理的甲基化和非甲基化DNA。 因此,我们使用MSP来测量Opg的甲基化状态。甲基化特异性Opg引物为:正向:TTCGGATTGGTGCATC; 背面:GCAAACTCTAAATTTCGCG。 基因组DNA在以下条件下扩增:95°C 5 min,然后在95°C 40个循环中扩增30 s,57°C扩增30 s和72°C扩增30s,最后在72°C下扩增5 min。 MSP的产物最终在2%溴化乙锭染色的琼脂糖凝胶上显现。 DNA甲基化程度估计为每个扩增子的非甲基化引物循环阈值减去甲基化引子循环阈值。
2.14. 亚细胞分馏 根据制造商的说明,使用NE‐PER核和细胞质萃取试剂(Thermo Scientific)分离细胞核和细胞溶质部分。 简单地说,收集细胞并在冰上与冷CER I孵育10分钟。 然后,添加冰冷CER II,然后在冰上再培养5分钟。 在4000左右离心后 克 将上清液转移到新的试管中5min。 这是细胞质部分。 用冰凉NER悬浮颗粒,并在最高设置下每10分钟旋转15秒,共40分钟。 在4000左右离心后 克 5min后,收集上清液作为核组分。 从细胞质和细胞核提取物中分离出总RNA。 qPCR用于评估细胞核和细胞质部分的相对比例。 U6和GAPDH分别作为核标记和细胞质标记。
2.15条。 蛋白质印迹 通过使用补充有蛋白酶抑制剂(罗氏)的放射免疫沉淀分析(RIPA)缓冲液在4°C下制备全细胞和组织裂解物。 使用直接检测光谱仪(Merck)测量蛋白质浓度。 在还原条件下,用10%SDS‐PAGE钠分离20毫克蛋白质,然后通过电印迹将其转移到聚偏二氟乙烯(PVDF)膜上。 在用5%的非脂肪乳封闭后,将膜与来自抗PTBP1抗体Abcam的稀释原代兔抗体(ab133734;稀释1:2000)Opg(ab183910,稀释1:1000)、Ppar‐γ(ab59256,稀释1:100)、Fabp4(ab92501,稀释1:2000, 和β-actin(ab179467,稀释1:5000)在4°C下过夜。 然后在RT条件下将膜与物种匹配的二级抗体孵育2小时。使用增强化学发光底物检测免疫反应带,并通过BioRad-Chemidoc MP系统(Bio-Rad)捕获。 靶蛋白的折叠变化被标准化为看家蛋白β-actin的水平。
2.16. 统计分析 在所有体外研究中,实验至少进行了三次重复,每次至少有三个独立样本。 在体内研究中,所有动物研究中,每组有六只小鼠。 数据表示为平均值±标准偏差(SD)。 使用Student的 t吨 通过事后分析进行测试或单向方差分析。 使用IBM SPSS Statistics for Windows 23.0版(IBM)进行统计分析。 A类 第页 值<0.05被认为具有统计学意义。
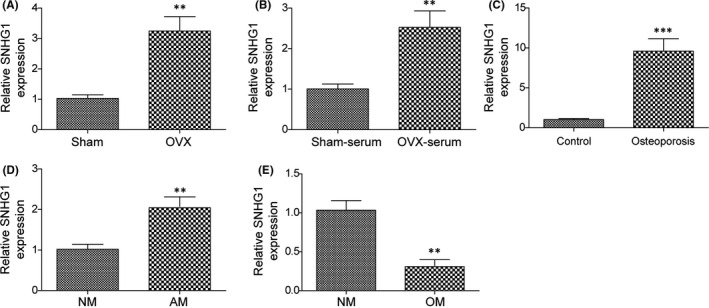
3.结果 3.1. LncRNA SNHG1与骨质疏松症患者骨髓基质干细胞分化相关 为了阐明lncRNA SNHG1在骨质疏松症中的作用及其与BMSC分化的关系,我们首先研究了lncRNA SNHG1的在体内小鼠模型和骨质疏松症患者中的表达。 qPCR显示,与假对照小鼠相比,OVX小鼠的骨组织和血清中lncRNA SNHG1的表达显著上调(图 1A、B ). 同样,我们观察到骨质疏松症患者组织中lncRNA SNHG1的表达高于对照组(图 1个 ). 我们进一步研究了成骨诱导剂或脂肪诱导剂刺激的BMSCs中lncRNA SNHG1的表达。 在诱导的第14天,脂肪分化的诱导显著增强了BMSCs中lncRNA SNHG1的表达,而经成骨诱导培养基处理的细胞中lncRNA SNHG1的表达显著下调(图 1D、E ). 这些结果表明,lncRNA SNHG1的表达与骨质疏松症中BMSC的分化有关。
图1。
LncRNA SNHG1与骨质疏松症中BMSC的分化有关。 用qPCR检测以下样品的LncRNA SNHG1表达:(A,B)OVX和羞耻小鼠的骨组织和血清; (C) 骨质疏松症患者的骨组织与对照组; (D) 脂肪诱导剂和生长介质处理的骨髓间充质干细胞; (E) 经成骨诱导剂和生长介质处理的骨髓间充质干细胞。 AM,脂肪诱导培养基; NM,正常生长培养基; OM,成骨诱导培养基。 使用Student’s t吨 ‐测试** 第页 < 0.01; *** 第页 < 0.001
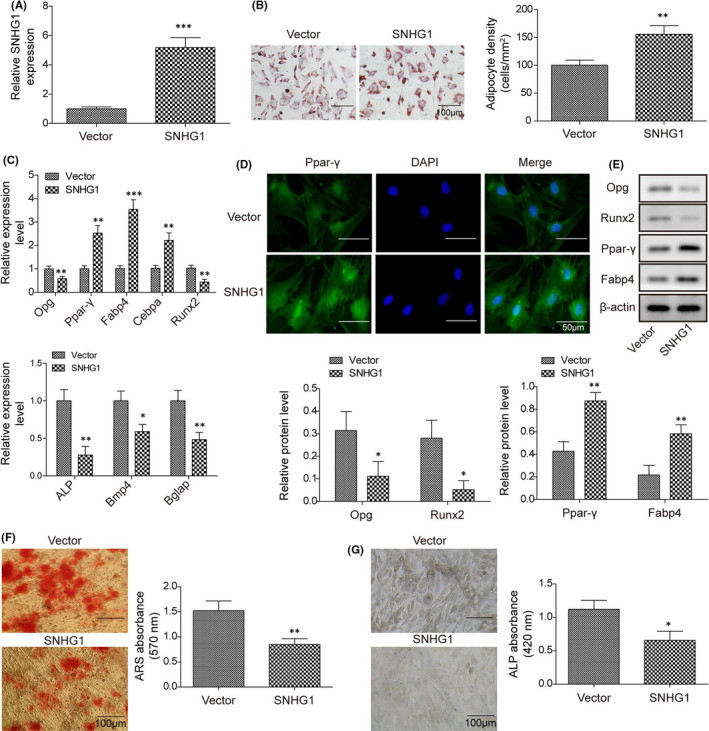
3.2. LncRNA SNHG1增强BMSC成脂分化但抑制BMSC成骨分化 我们探讨了lncRNA SNHG1在转染BMSCs的BMSC分化中的作用。 与用空载体转染的细胞相比,用pcDNA‐SNHG1转染的细胞中lncRNA SNHG1的表达显著上调,证实了pcDNA‐SNHG1的成功转染(图 2安培 ). ORO染色结果表明,转染pcDNA-SNHG1的BMSCs经成脂诱导培养基处理后,其成脂分化能力增强,导致脂滴形成和脂肪细胞密度增加(图 2B型 ). 然而,ARS染色显示,经pcDNA‐SNHG1转染的BMSCs经成骨诱导介质处理后,其成骨分化能力低于用空载体转染的,并且形成的矿化结节较少(图 2楼 ). 我们进一步检测了相应组中成骨相关基因(Opg、Runx2、ALP、Bmp4、Bglap)和脂肪生成相关基因(Ppar‐γ、Fabp4、Cebpa)的表达,以证实我们的观察结果。 与染色结果一致,SNHG1过度表达的BMSCs中脂肪生成相关基因的基因和蛋白表达增加,而成骨相关基因的表达减少(图 2C、E ). 此外,通过免疫荧光染色(IF),lncRNA SNHG1的过度表达增加了BMSCs中的Ppar‐γ荧光信号(图 二维 )但ALP染色后,染色面积较小且不太强烈,抑制了ALP活性(图 2G(2G) ). 综上所述,我们的观察结果表明,lncRNA SNHG1通过增强脂肪分化但抑制成骨分化来调节BMSC分化。
图2。
LncRNA SNHG1增强BMSC成脂分化,但抑制BMSC成骨分化。 用pcDNA‐SNHG1或空载体转染BMSCs。 (A) 转染pcDNA‐SNHG1或空载体的BMSCs的LncRNA SNHG1表达; (B) 用代表性图像计算ORO染色后的脂肪细胞密度; (C) 通过qPCR检测Opg、Runx2、Ppar‐γ、Fabp4、ALP、Bmp4、Bglap和Cebpa的mRNA表达; (D) Ppar‐γ的免疫荧光染色; (E) western blot检测Opg、Runx2、Ppar‐γ和Fabp4蛋白水平; (F) 用代表图像进行ARS染色后的ARS吸光度; (G) 用代表图像进行ALP染色后的ALP吸光度。 使用Student’s t吨 ‐测试* 第页 < 0.05; ** 第页 < 0.01; *** 第页 < 0.001
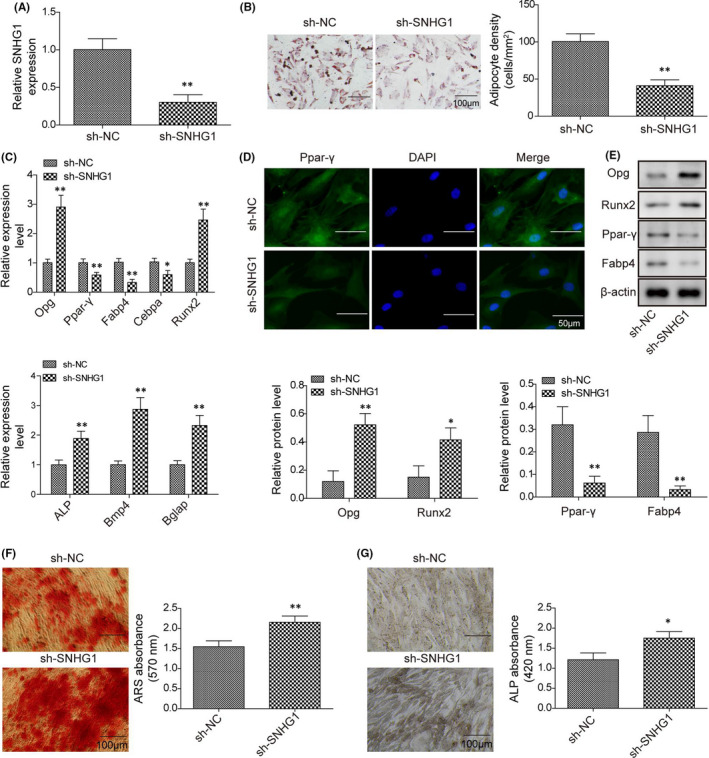
3.3. 沉默lncRNA SNHG1抑制BMSC成脂分化但促进BMSC成骨分化 我们进一步探讨了lncRNA SNHG1沉默对转染sh-SNHG1的BMSC分化的影响。 在重复实验中,与sh-NC转染相比,sh-SNHG1转染成功抑制了骨髓间充质干细胞中lncRNA SNHG1的表达(图 3A级 ). 依次进行BMSC的成骨或成脂分化诱导以及一系列染色。 还检测了成骨和脂肪生成相关基因的基因和蛋白表达。 正如预期的那样,lncRNA SNHG1沉默显著抑制了BMSCs的成脂分化能力,表现为脂滴形成减少,但增强了成骨分化能力,当用成脂和成骨诱导介质处理时,表现为矿化结节形成和ALP活性增加, 分别(图 3B、F ,G)。 同时,在sh-SNHG1转染的BMSCs中,脂肪生成相关基因的基因和蛋白表达降低,而成骨相关基因的表达增加(图 3C、D ,E)。 这些发现支持lncRNA SNHG1在调节BMSC分化中的作用。
图3。
沉默lncRNA SNHG1抑制BMSC脂肪分化,但促进BMSC成骨分化。 骨髓间充质干细胞转染sh-SNHG1或sh-NC。 (A) 转染sh-SNHG1或sh-NC的BMSCs的LncRNA SNHG1表达; (B) 用代表性图像计算ORO染色后的脂肪细胞密度; (C) qPCR检测Opg、Runx2、Ppar‐γ、Fabp4、ALP、Bmp4、Bglap和Cebpa的mRNA表达; (D) Ppar‐γ的免疫荧光染色; (E) western blot检测Opg、Runx2、Ppar‐γ和Fabp4蛋白水平; (F) 用代表图像进行ARS染色后的ARS吸光度; (G) 用代表图像进行ALP染色后的ALP吸光度。 使用Student’s t吨 ‐测试* 第页 < 0.05; ** 第页 < 0.01
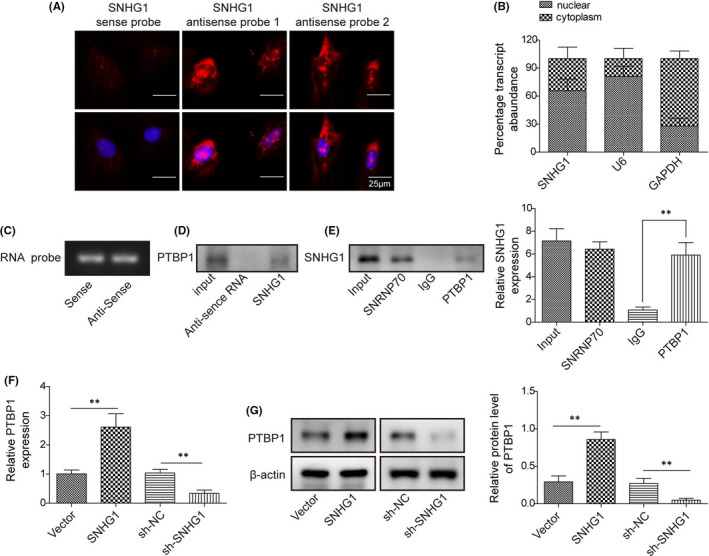
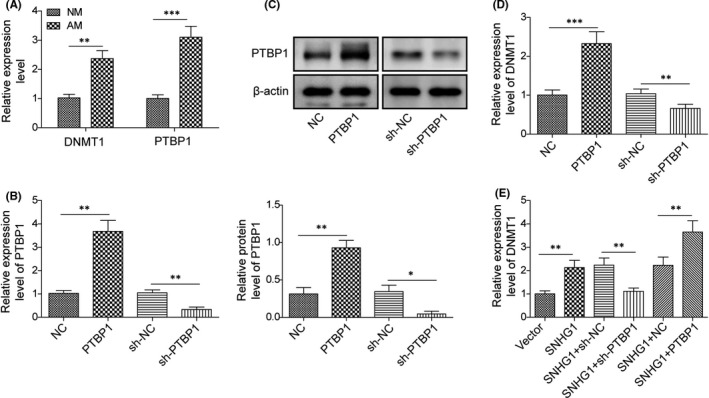
3.4。 LncRNA SNHG1与PTBP1相互作用并促进PTBP1的表达 为了更好地了解lncRNA SNHG1的功能和机制,我们通过FISH和qPCR检测了lncRNA SNHG1在正常培养基中培养的细胞中的特异亚细胞定位和细胞分馏。 FISH显示lncRNA SNHG1广泛定位于细胞核(图 4A级 ). qPCR显示lncRNA SNHG1主要表现为核分布(>60%)(图 4B类 ). 为了确定lncRNA SNHG1与PTBP1的潜在相互作用,我们使用体外合成的生物素化正反义SNHG1RNAs对骨髓基质细胞进行了RNA下拉分析(图 4摄氏度 ). 与BMSC提取物孵育后,western blot显示PTBP1被鉴定为lncRNA SNHG1的结合伙伴(图 4D(四维) ). 或者,使用PTBP1抗体进行RIP分析,以检测PTBP1沉淀物中的lncRNA SNHG1。 我们发现,与抗IgG对照组相比,PTBP1抗体显著沉淀lncRNA SNHG1(图 第四版 ),进一步证实了lncRNA SNHG1和PTBP1之间的相互作用。 为了确定lncRNA SNHG1和PTBP1相互作用的生物学效应,我们检测了lncRNA SNHG1过度表达或沉默的BMSCs中PTBP1的表达。 lncRNA SNHG1的过度表达增加了PTBP1的基因和蛋白表达,而lncRNA的沉默则降低了PTBPl的表达(图 4F、G ). 这些结果表明,lncRNA SNHG1与PTBP1相互作用并促进PTBP1的表达。
图4。
LncRNA SNHG1与PTBP1相互作用并促进PTBP1的表达。 (A) 骨髓基质干细胞中SNHG1的FISH(红色)。 DAPI染色图像为蓝色; (B) 通过qPCR分离MSCs中的SNHG1。 U6和GAPDH分别作为核和细胞质对照; (C) 合成的生物素化的正义和反义SNHG1探针; (D) RNA下拉,然后进行蛋白质印迹。 BMSC裂解物与正反义生物素标记的SNHG1探针孵育。 下拉后,用western blot检测探针招募的PTBP1; (E) RIP分析使用抗PTBP1抗体检测PTBP1和SNHG1之间的相互作用。 SNRP70或正常IgG分别作为阳性和阴性对照。 (F) 通过qPCR检测转染pcDNA-SNHG1/空载体或sh-SNHG1/sh-NC的BMSCs中PTBP1的mRNA表达; (G) 通过western blot检测转染pcDNA-PTBP1/empty载体或sh‐PTBP1/sh‐NC的BMSCs中PTBP1的蛋白水平。 使用Student’s t吨 测试或单向方差分析,适当时进行事后分析** 第页 < 0.01
3.5. LncRNA SNHG1与PTBP1相互作用并促进DNMT1的表达 先前的研究表明,RNA结合蛋白可以影响DNMT的稳定性并调节DNA甲基化。
28 ,
29 我们对DNMT1特别感兴趣,它是维持DNA甲基化的关键酶之一。
30 为了阐明PTBP1和DNMT1在BMSC分化中的潜在作用,我们检测了经脂肪分化诱导培养基处理14天的BMSC中PTBP1与DNMT1的基因表达。 有趣的是,用成脂诱导剂治疗14天后,BMSC中PTBP1和DNMT1的表达显著上调(图 5A级 ). 然后,我们在用pcDNA-PTBP1、sh-PTBP1或其相应的阴性对照转染的BMSCs中测试了PTBP1对DNMT1的调节作用。 qPCR和western blot均显示PTBP1正向调节DNMT1的表达(图 5B–D级 ). 值得注意的是,转染pcDNA-SNHG1上调了DNMT1的表达,当与pcDNA-SNHG1和pcDNA-PTBP1联合转染时,这一点更为显著,而沉默PTBP1部分逆转了lncRNA-SNHJ1对DNMT1作用(图 第五版 )表明lncRNA SNHG1通过与PTBP1相互作用促进DNMT1的表达。
图5。
LncRNA SNHG1与PTBP1相互作用并促进DNMT1的表达。 (A) 应用qPCR检测脂肪分化诱导剂处理14天的骨髓间充质干细胞PTBP1和DNMT1的mRNA表达; (B,C)qPCR检测PTBP1的mRNA表达; (C) western blot检测PTBP1蛋白水平; (D) 用qPCR检测DNMT1的mRNA表达; (E) 通过qPCR检测DNMT1在不同转染方案的不同组中的mRNA表达。 NM,正常生长培养基; AM,脂肪诱导培养基。 使用Student’s t吨 测试或单向方差分析,适当时进行事后分析* 第页 < 0.05; ** 第页 < 0.01; *** 第页 < 0.001
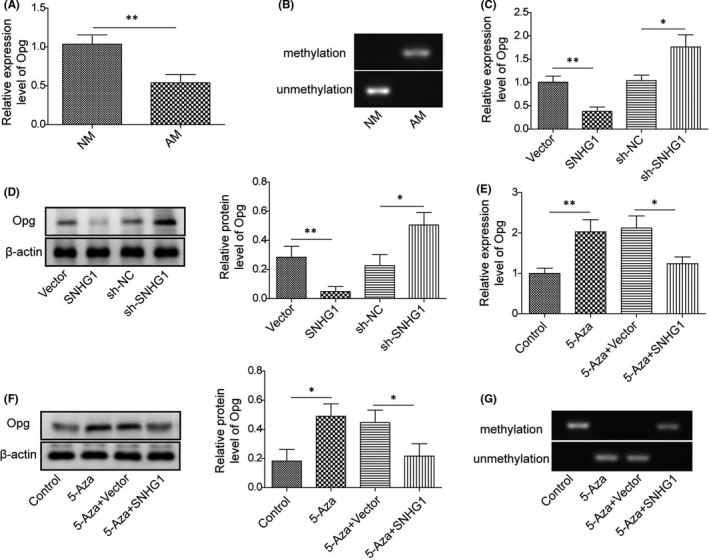
3.6. LncRNA SNHG1增强Opg甲基化以抑制Opg的表达 我们进一步研究了lncRNA SNHG1在BMSC成脂分化中的作用是否与经成脂诱导介质处理的BMSC中Opg甲基化的调节有关。 qPCR和MSP显示,用脂肪诱导剂治疗14天可抑制BMSCs中Opg的表达,但可促进Opg甲基化(图 6A、B ). 至于lncRNA SNHG1的作用,SNHG1-过表达对Opg的基因和蛋白表达具有显著的抑制作用,而SNHG1沉默增强了Opg的表达(图 6C、D ). 此外,DNA去甲基化剂5‐Aza处理增加了Opg的表达水平,而SNHG1过度表达消除了这些影响(图 6E、F ). MSP的DNA甲基化分析证实,5‐Aza显著抑制了Opg甲基化,而SNHG1的过度表达促进了Opg DNA甲基化(图 6克 ). 我们观察到的甲基化状态与基因表达之间的反向关系表明,Opg甲基化明显抑制Opg表达。 然而,lncRNA SNHG1可能增强Opg甲基化,从而下调Opg的表达。
图6。
LncRNA SNHG1增强Opg甲基化以抑制Opg的表达。(A)经脂肪分化诱导剂处理14天的骨髓间充质干细胞中Opg mRNA的qPCR表达; (B) MSP分析以检测Opg的甲基化状态; (C) qPCR检测转染pcDNA-SNHG1/空载体或sh-SNHG1/sh-NC的BMSCs中Opg的mRNA表达; (D) 通过western blot检测转染pcDNA-SNHG1/空载体或sh‐SNHG1/sh‐NC的BMSCs中Opg的蛋白水平; (E,F)5-Aza治疗96 h后,pcDNA-SNHG1或空载体转染BMSCs中Opg的mRNA表达和蛋白水平; (G) MSP分析检测5-Aza治疗96小时后pcDNA-SNHG1或空载体转染MSCs中Opg的甲基化状态。 NM,正常生长培养基; AM,脂肪诱导培养基。 使用Student’s t吨 测试或单向方差分析,适当时进行事后分析* 第页 < 0.05; ** 第页 < 0.01
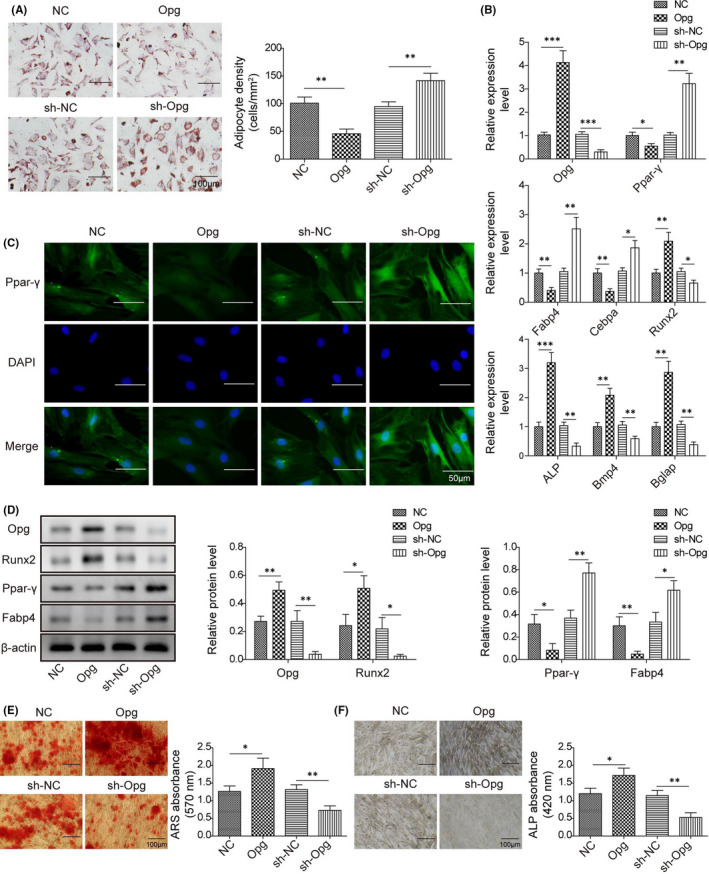
3.7. Opg调节BMSCs的细胞分化 用成骨或脂肪诱导培养基处理14天的骨髓间充质干细胞,检测Opg在其分化中的作用。 一贯地,Opg过度表达显著降低了脂滴形成和脂肪生成相关基因(Ppar‐γ、Fabp4和Cebpa)的基因和蛋白表达,但增加了矿化结节形成、ALP活性和骨生成相关基因的表达(Opg、Runx2、ALP、Bmp4和Bglap)(图 7A至F ). 当Opg被击倒时,所有这些影响都被逆转了。 这些结果表明,Opg以抑制脂肪生成但促进骨生成的方式调节BMSC分化。
图7。
Opg调节BMSCs的细胞分化。 用pcDNA-Opg/empty载体或sh‐Opg/sh‐NC转染BMSCs。 (A) 用代表性图像计算ORO染色后的脂肪细胞密度; (B) 通过qPCR检测Opg、Runx2、Ppar‐γ、Fabp4、ALP、Bmp4、Bglap和Cebpa的mRNA表达; (C) Ppar‐γ的免疫荧光染色; (D) western blot检测Opg、Runx2、Ppar‐γ和Fabp4蛋白水平; (E) 用代表图像进行ARS染色后的ARS吸光度; (F) 用代表图像进行ALP染色后的ALP吸光度。 使用单因素方差分析和事后分析进行比较* 第页 < 0.05; ** 第页 < 0.01; *** 第页 < 0.001
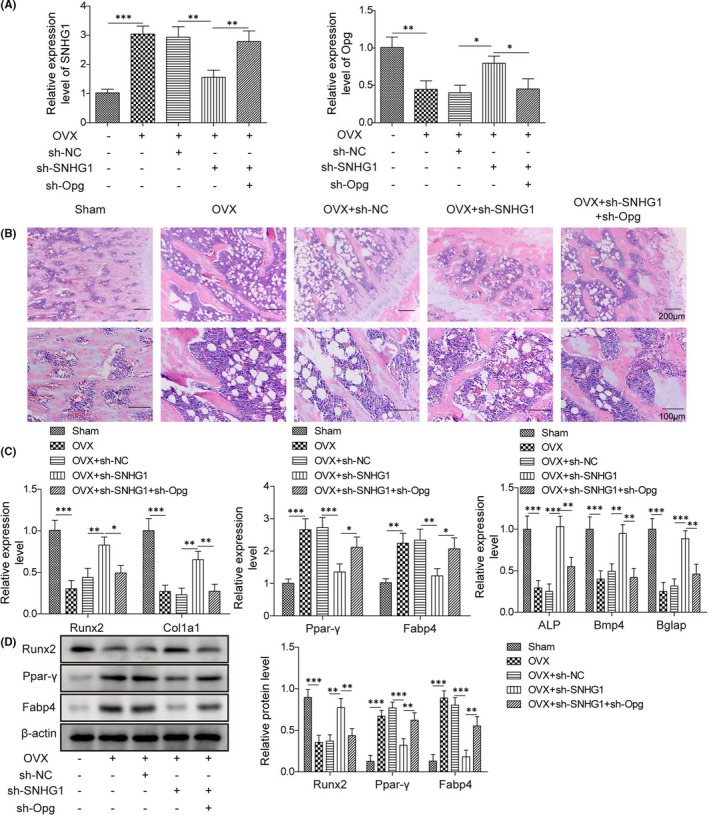
3.8. LncRNA SNHG1沉默在体内改善骨质疏松症 我们进行的体外研究表明,lncRNA SNHG1通过影响Opg的甲基化状态抑制Opg的表达,导致BMSC脂肪分化增强,这可能导致骨质疏松。 为了证实这一点,我们进一步研究了lncRNA SNHG1在骨质疏松小鼠体内模型中的作用。 用sh-SNHG1或sh-Opg转染成功抑制了SNHG1或Opg的表达(图 8安 ). 小鼠股骨的HE染色进一步证实了OVX组的骨质疏松症,其中小梁排列不良,数量和厚度减少,而小梁分离减少,如前所述
31 (图 8B类 ). sh-SNHG1转染显著改善了OVX组的骨质疏松性变化,而与sh-Opg共转染部分逆转了SNHG1沉默的作用(图 8B类 ). 此外,qPCR显示,SNHG1沉默显著上调骨生成相关基因(Runx2、ALP、Bmp4、Bglap和Col1a1)的基因表达,但下调脂肪生成相关基因的基因表达(Ppar‐γ和Fabp4)(图 8摄氏度 ). Western bolt在这些基因的蛋白质水平上显示了一致的结果(图 第8天 ). 因此,我们得出结论,SNHG1在骨质疏松体内模型中的骨质疏松变化中发挥作用,这可能是由Opg介导的。
图8。
LncRNA SNHG1沉默可在体内改善骨质疏松症。 通过尾静脉用sh‐SNHG1和/或sh‐Opg治疗OVX小鼠3周。 (A) qPCR检测SNHG1和Opg mRNA的表达; (B) 小鼠股骨的血红素和曙红染色; (C) qPCR检测Col1a1、Runx2、Ppar‐γ、ALP、Bmp4、Bglap和Fabp4的mRNA表达; (D) western blot检测Runx2、Ppar‐γ和Fabp4蛋白水平。 使用单因素方差分析和事后分析进行比较* 第页 < 0.05; ** 第页 < 0.01; *** 第页 < 0.001
4.讨论 lncRNA作为表观遗传调控因子之一,在基因表达和多种生物过程中发挥着重要作用。
32 一些研究表明,lncRNAs通过影响成骨细胞和破骨细胞的增殖和功能以及BMSCs的分化,参与骨重塑。
33 例如,已证明lncRNA ANCR可抑制成骨细胞分化,并对维持成骨细胞处于未分化状态至关重要。
34 相反,lncRNA H19促进了BMSCs的成骨细胞分化,Runx2和ALP的表达增加证明了这一点。
9 在本研究中,lncRNA SNHG1被发现可以增强BMSC的成脂分化,但抑制BMSC的成骨分化,表明其在骨质疏松症中的作用。 事实上,SNHG1是一种新的致癌lncRNA,在许多癌症中异常表达,与细胞生长、迁移和侵袭有关。
35 ,
36 与我们的观察结果一致,之前有报道称lncRNA SNHG1通过负向调节p38 MAPK信号通路抑制BMSCs的成骨分化。
11 因此,lncRNA SNHG1可能是研究骨质疏松症的一个有趣的靶点,骨质疏松症是由BMSC分化为脂肪细胞而非成骨细胞引起的。
据报道,一些lncRNA通过合作或竞争相互作用结合RBP来调节基因表达。
37 PTBP1是一种参与RNA生物生成所有步骤的RBP,通过与lncRNA(如H19、HCG22和MACC1‐AS1)的相互作用与代谢和癌症相关。
38 ,
39 ,
40 在我们的研究中,lncRNA SNHG1和PTBP1之间的相互作用也通过RNA下拉和免疫沉淀试验得到了证实。 此外,通过与PTBP1相互作用,lncRNA SNHG1上调DNMT1的表达,进而促进Opg超甲基化。 通过DNMT将CH3甲基添加到胞嘧啶中的DNA甲基化被认为是一种长期、相对稳定的表观遗传修饰,导致转录抑制和靶基因下调。
41 在这方面,越来越多的证据表明,Opg的甲基化状态可能在骨质疏松症的发病机制中起到“主开关”的作用。
19 ,
42 Behera等人的观察结果。
43 在代谢物诱导的骨质疏松小鼠模型中,DNMT1表达增加和Opg基因的高甲基化进一步支持了DNA甲基化在骨质疏松症Opg表达调控中的作用。
对转基因动物的一系列研究也表明Opg对BMSC分化和脂肪生成的调节作用。
44 ,
45 在Opg基因敲除小鼠中,小鼠出现了早期骨质疏松症,其特征是骨髓中的小梁孔隙度和脂肪细胞积聚增加,
17 而给予Opg蛋白可有效逆转Opg缺乏小鼠的骨质疏松性骨表型。
44 我们发现,与转染对照载体的细胞相比,Opg过度表达抑制了BMSCs中的脂肪生成,但促进了骨生成。 有趣的是,我们还发现lncRNA SNHG1调节骨质疏松小鼠Opg的表达; 敲除SNHG1可改善骨质疏松性改变,同时敲除Opg可部分逆转SNHG1-沉默对骨量丢失的有利影响。然而,在我们的研究中,缺乏治疗期间基因和蛋白表达的比较。 此外,未进行MicroCT(或µCT)分析以支持SNHG1在骨质疏松症中的功能。 这些是本研究的局限性。
总之,我们的研究表明,lncRNA SNHG1通过与PTBP1的相互作用上调DNMT1的表达,导致Opg高甲基化并降低Opg的表达,从而增强BMSC的脂肪分化并导致骨质疏松。 本研究进一步揭示了lncRNA SNHG1/PTBPT1/DNMT1/Opg通路在骨质疏松症发病机制中的分子机制,并为制定新的骨质疏松症防治措施提供了新思路。
作者贡献
小余: 资金获取(同等); 监督(同等); 书写原稿(同等)。 宋梦生: 数据管理(同等)。 彭泽荣: 调查(同等); 验证(等同)。 陈宪军(Xian‐Jun Chen): 方法(同等); 可视化(相等)。 林石(Lin Shi): 形式分析(相等)。 王成浩: 概念化(平等); 书写原稿(同等)。 清江庞: 方法(同等); 写作-审查和编辑(同等)。
道德认可 所有动物程序均按照美国国立卫生研究院(National Institutes of Health Guide)关于实验动物护理和使用的指南进行。 所有方案均经我院动物伦理委员会批准。 该临床研究得到了我院伦理委员会的批准,并获得了所有参与者的书面知情同意。
Yu X,Song M‐S,Rong P‐Z等。LncRNA SNHG1通过与PTBP1相互作用促进DNMT1介导的Opg超甲基化,从而调节BMSC的成脂分化。 细胞分子医学杂志2022; 26:60–74. 10.1111/jcmm.16982
资金筹措信息
本研究得到了浙江省医药卫生科技项目(2018ky156)、宁波市自然科学基金(2019A610242)和中国科学院华美医院华美重点研究基金(2021HMZD04)的资助
数据可用性声明 本研究中产生或分析的所有数据均包含在本文中。
参考文献
1 Aspray TJ,Hill TR。骨质疏松和老化骨骼。 亚细胞生物化学。 2019; 91:453‐476. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
2 Jordan KM,Cooper C.骨质疏松症流行病学。 最佳实践研究临床风湿病。 2002; 16:795‐806. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
三。 Tarrant SM,巴洛赫ZJ。 骨质疏松性骨折手术治疗的全球负担。 《2020年世界外科杂志》; 44:1009‐1019. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
4 Gatti D,Fassio A.《绝经后妇女骨质疏松症的药理学管理:最新进展》,《大众临床药理学杂志》。 2019; 第26页:e1‐e17。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
5 Armas LA,Recker RR。骨质疏松症的病理生理学:新的机制见解。 《北美内分泌代谢临床》2012; 41:475‐486. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
6 胡力,尹C,赵F,阿里A,马J,钱A.间充质干细胞:成骨细胞或脂肪细胞的细胞命运决定及其在骨质疏松症治疗中的应用。 国际分子科学杂志。 2018; 19:360. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
7 Hombach S,Kretz M.非编码RNA:分类、生物学和功能。 高级实验医学生物。 2016; 937:3‐17. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
8 Huynh NP、Anderson BA、Guilak F、McAlinden A.长时间非编码RNA在骨骼生物学和疾病中的新兴作用。 连接组织研究2017; 58:116‐141. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
9 Huang Y,Zheng Y,Jia L,Li W.Long非编码RNA H19通过衍生miR‐675,通过TGF‐beta1/Smad3/HDAC信号通路促进成骨细胞分化。 干细胞。 2015; 33:3481‐3492. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
10 王毅,罗TB,刘力,崔志强。 LncRNA LINC00311通过靶向DLL3的notch信号通路促进骨质疏松大鼠破骨细胞的增殖和分化。 细胞生理生化。 2018; 47:2291‐2306. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
11 姜毅,吴伟,焦庚,陈毅,刘海林,LncRNA SNHG1通过Nedd4调节p38 MAPK通路,从而抑制骨髓间充质干细胞的成骨分化。 生命科学。 2019; 228:208‐214. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
12 Ferre F,Colantoni A,Helmer‐Citterich M.揭示蛋白质-lncRNA相互作用。 简要生物信息。 2016; 17:106‐116. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
13 Vuong JK、Lin CH、Zhang M、Chen L、Black DL、Zheng S.PTBP1和PTBP2在神经元前体mRNA剪接中具有特异和冗余功能。 细胞报告2016; 17:2766‐2775. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
14 Fu X,Xie F,Gong F,等。PTBP1信号的抑制是间充质干细胞诱导低恶性肿瘤细胞侵袭的原因。 生物化学生物物理学报Mol Cell Res.2018; 1865:1552‐1565. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
15 Li J,Yang Y,Fan J,等。长非编码RNA ANCR通过促进PTBP1与ID2的结合抑制间充质干细胞向最终内皮细胞的分化。 细胞死亡疾病。 2019; 10:492. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
16 Gowher H,Jeltsch A.哺乳动物DNA甲基转移酶:新发现和开放问题。 生物化学Soc Trans。 2018; 46:1191‐1202. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
17 张磊,刘敏,周旭,等。骨保护素(OPG)在骨髓脂肪生成中的作用。 细胞生理生化。 2016; 40:681‐692. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
18 Hsu H,Lacey DL,Dunstan CR,等。肿瘤坏死因子受体家族成员RANK介导骨保护素配体诱导的破骨细胞分化和活化。 美国国家科学院院刊1999; 96:3540‐3545. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
19 王平,曹毅,詹德,等。DNA甲基化对原发性骨质疏松症OPG/RANKL表达的影响。 国际医学科学杂志。 2018; 15:1480‐1485. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
20 Hu Y,Ma Z,He Y,Liu W,Su Y,Tang Z.LncRNA‐SNHG1通过调节DNMT1促进胃癌细胞增殖。 生物化学与生物物理研究委员会。 2017; 491:926‐931. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
21 Li SJ,Wang L,Sun ZX,Sun SJ,Gao J,Ma RL。LncRNA SNHG1促进肝癌的发展。 欧洲药理学评论。 2019; 23:2768‐2776. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
22 Lee EJ、Kim JL、Kim YH、Kang MK、Gong JH、Kang-YH。 Phloretin促进小鼠巨噬细胞破骨细胞凋亡,并抑制雌激素缺乏诱导的小鼠骨质疏松症。 植物学。 2014; 21:1208‐1215. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
23 乔莉,刘迪,李CG,王勇杰。 MiR‐203对于绝经后骨质疏松症中间充质干细胞从成骨分化向成脂分化的转变至关重要。 欧洲药理学评论。 2018; 22:5804‐5814. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
24 Yang F,Yang L,Li Y,等。褪黑素保护骨髓间充质干细胞免受铁超载诱导的异常分化和衰老。 皮纳尔研究杂志,2017; 63(3):e12422‐e12472。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
25 古普塔A,Mo YY。 原位杂交检测培养细胞和石蜡包埋组织标本中的microRNA。 方法分子生物学。 2011; 676:73‐83. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
26 Zhang L,Yang Z,Trottier J,Barbier O,Wang L.长非编码RNA MEG3通过与PTBP1相互作用诱导胆汁淤积性肝损伤,以促进shp mRNA衰变。 肝病学。 2017; 65:604‐615. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
27 Sasaki M、Anast J、Bassett W、Kawakami T、Sakuragi N、Dahiya R.亚硫酸氢盐转化特异性和甲基化特异性PCR:准确评估CpG甲基化的敏感技术。 生物化学与生物物理研究委员会。 2003年; 309:305‐309. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
28 Wang X,Duan CG,Tang K,等。RNA结合蛋白通过控制内含子异染色质基因IBM1的mRNA处理来调节植物DNA甲基化。 美国国家科学院院刊2013; 110:15467‐15472. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
29 Lopez de Silanes I、Gorospe M、Taniguchi H等。RNA结合蛋白HuR通过稳定DNMT3b mRNA调节DNA甲基化。核酸研究,2009; 37:2658‐2671. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
30 Rhee I、Bachman KE、Park BH等。DNMT1和DNMT3b协同沉默人类癌细胞中的基因。 自然。 2002; 416:552‐556页。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
31 李斌,赵杰,马JX,等。DNMT1的过度表达导致H19启动子的高甲基化和Erk信号通路在废用性骨质疏松症中的抑制。 骨头。 2018; 111:82‐91. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
32 朱杰,傅赫,吴勇,郑霞。lncRNAs的功能和lncRNA-蛋白质相互作用的途径。 中国生命科学。 2013; 56:876‐885. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
33 吴庆英,李旭,缪振宁,等。长非编码RNA:骨质疏松症的新调控编码。 前内分泌。 2018; 9:587. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
34 Zhu L,Xu PC。下调的LncRNA-ANCR通过靶向EZH2和调节Runx2表达促进成骨细胞分化。 生物化学与生物物理研究委员会。 2013; 432:612‐617. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
35 Thin KZ,Tu JC,Raveendran S.Long非编码SNHG1在癌症中的作用。 临床化学学报。 2019; 494:38‐47. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
36 Liu Y,Yang Y,Li L,等。LncRNA SNHG1促进宫颈癌细胞增殖、迁移和侵袭。 生物化学细胞生物学。 2018; 96:38‐43. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
37 Kim C、Kang D、Lee EK、Lee JS。 氧化应激、细胞衰老和年龄相关疾病中的长链非编码RNA和RNA结合蛋白。 Oxid Med细胞寿命。 2017; 2017:2062384. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
38 姜丹,张勇,杨磊,等。长非编码RNA HCG22通过调节PTBP1抑制膀胱癌细胞的增殖和转移。 细胞生理学杂志。 2020; 235:1711‐1722. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
39 Liu C,Yang Z,Wu J,等。长非编码RNA H19与多嘧啶道结合蛋白1相互作用,重新编程肝脏脂质稳态。 肝病学。 2018; 67:1768‐1783. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
40 Zhang X,Zhou Y,Chen S,Li W,Chen W,Gu W.LncRNA MACC1‐AS1海绵多个miRNA和RNA结合蛋白PTBP1。 肿瘤发生。 2019; 8:73. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
41 Pathania R、Ramachandran S、Elangovan S等。DNMT1对乳腺和癌症干细胞维持和肿瘤发生至关重要。 国家公社。 2015; 6:6910. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
42 Delgado‐Calle J、Sanudo C、Fernandez AF、Garcia‐Renedo R、Fraga MF、Riancho JA。 DNA甲基化在人类骨骼RANKL‐OPG系统调节中的作用。 表观遗传学。 2012; 7:83‐91. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
43 Behera J、George AK、Voor MJ、Tyagi SC、Tyagi N。在小鼠高同型半胱氨酸血症期间,硫化氢通过OPG/RANKL调节表观遗传学减轻骨丢失。 骨头。 2018; 114:90‐108. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
44 Min H、Morony S、Sarosi I等。骨保护素通过抑制骨内破骨细胞逆转骨质疏松症,并通过阻断类似破骨细胞生成的过程防止血管钙化。 实验医学杂志2000; 192:463‐474. [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
45 王毅,颜强,赵毅,等。小鼠骨内黏附蛋白Pinch1和Pinch2调节骨稳态。 JCI洞察力。 2019; 4:e131692。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
关联数据 本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。
数据可用性声明 本研究中产生或分析的所有数据均包含在本文中。