总结 内化和泛素化的信号受体在运输(ESCRT)机械所需的内质体分选复合体的辅助下,通过其在管腔内萌芽进入多泡体而沉默。 HD-PTP是一种ESCRT蛋白,与ESCRT-0、-I和-III蛋白形成复合物,并与Endofin结合,后者是一种限制在内切体中的FYVE域蛋白,其作用知之甚少。 通过近距离生物素化,我们发现内啡肽与ESCRT成分形成复合物,内啡肽的缺失通过阻碍其溶酶体传递,增加了整合素α5和EGF受体的质膜密度和稳定性。 这与持续的受体信号和增加的细胞迁移相一致。 用野生型Endofin或HD-PTP补充Endofin-或HD-PTP-depleted细胞,但不与含有受损Endofin/HD-PTP关联或细胞溶质Endofin的突变体互补,恢复EGFR溶酶体递送。 Endofin还促进Hrs与HD-PTP的间接相互作用。 我们的研究结果共同表明,HD-PTP和ESCRT-0相互依赖的泛素化跨膜货物分拣需要Endofin,以确保有效的受体脱敏和溶酶体递送。
图形摘要
结果 内啡肽与ESCRT和EGF受体形成复合物 内皮素直接与Tom1和HD-PTP结合,两者在受体贩运中都有直接作用( Doyotte等人。, 2008 ; Gahloth等人。, 2017年a ; Katoh等人。, 2004 ; Kharitidi等人。, 2015 ; Seet等人。, 2004 ; Yamakami等人。, 2003 ). 内啡肽的相互作用组可能反映其功能; 然而,它的特征仍然很差。 为了进一步详细研究HD-PTP和Endofin的结合伙伴,设计了两个蛋白质组工作流程。 在第一个实验中,我们进行了依赖邻近性的生物素标记(BioID),其中一种生物素连接酶(BirA*)融合到HD-PTP和Endofin的N或C末端,作为“诱饵”,用生物素共价标记附近的蛋白质(“普利斯”),然后可以通过MS纯化和鉴定( Lambert等人。, 2015 ). 随后,采用亲和纯化质谱(AP-MS)方法,诱导表达标记的HD-PTP和Endofin作为诱饵。 我们的研究结果通过BioID和AP-MS揭示了Endofin是HD-PTP的一个强大的邻近相互作用物( 图1 A和 S1(第一阶段) B) ,并且HD-PTP与来自不同ESCRT复合物(ESCRT-0、-I和-III)的各种ESCRT蛋白形成复合物( 图1 A) ●●●●。 同样,HD-PTP被确定为Endofin-BirA的邻近相互作用物*( 图1 A) BioID未检测到Endofin和ESCRT蛋白之间的相互作用( 图1 A) AP-MS显示,各种ESCRT蛋白(HD-PTP、ESCRT-I和-III)与Endofin复合( 图S1 B) ●●●●。 此外,我们的结果表明HD-PTP是氯氰菊酯重链1(CLTC)和细胞内更广泛的内吞和转运机制的强大相互作用体( 图S1 B) ●●●●。 总的来说,这些蛋白质组分析表明,内啡肽和HD-PTP与内吞机制和ESCRT形成复合物(进一步注释于 图S1 A) ●●●●。 为了更好地了解内啡肽在HD-PTP中的作用,使用STRING数据库对BioID数据进行了进一步分析,通过该数据库设计交互网络并将其聚集到不同的细胞过程中。 为了支持我们的观察,Endofin的网络与受体贩运有关,这一发现与HD-PTP的发现类似( 图S2 和 第3章 ) ( Doyotte等人。, 2008 ).
图1。
内啡肽与ESCRT和EGF受体形成复合物
(A) BioID屏幕的点图显示ESCRT蛋白与HD-PTP和Endofin非常接近。 将标记生物素连接酶(BirA*)融合到HD-PTP和Endofin的N或C末端,并用作诱饵。 将构建物转染到Flp-In T-REx 293T细胞中。 点图中的填充阴影表示平均光谱计数(Avg Spec),点的大小表示所有猎物的相对丰度,外圆颜色表示BFDR值。
(B) 与模拟转染细胞相比,对瞬时转染myc-Endofin的293T细胞进行共免疫沉淀(Co-IP)。 免疫沉淀Myc-Endofin(Myc抗体),用Western blots检测HD-PTP、ESCRT-0(Hrs和STAM2)和ESCRT-I(Tsg101和UBAP1)的co-IP。 加载全细胞裂解液(WCL)以显示蛋白质含量。 显示了小时的低暴露(LE)和高暴露(HE)免疫印迹。数据来自n=3个独立实验。
(C) TurboID屏幕的点图显示了在存在或不存在EGF刺激的情况下(100 ng/mL,15分钟),ESCRT蛋白和其他与内吞作用和EGFR激活密切相关的蛋白。 将快速作用的miniTurbo生物素连接酶(MT)融合到EGFR(EGFR-MT)的C末端,并转染到Flp-In T-REx-HeLa细胞中,用作诱饵。
为了验证内吞素、HD-PTP和ESCRT在早期内吞体上形成复合物,对293T细胞器进行了等密度蔗糖梯度离心。 虽然含有LAMP1和Tom20的细胞组分中不存在内啡肽和HD-PTP,但溶酶体和线粒体标记物分别不存在( 图S1 C) ,它们一起存在于含有ESCRT蛋白(Hrs、Tsg101和UBAP1)的细胞级分中。 Tom1和早期内体标记物EEA1也存在于这些级分中。 此外,亚细胞分馏后的尺寸排除色谱(HPLC)显示,内啡肽、HD-PTP和ESCRT-0组分(Hrs和STAM2)是525至927 kDa范围内的大蛋白复合物的一部分,但ESCRT-I组分Tsg101主要存在于分子量较低的复合物中( 图S1 D) ●●●●。 作为与ESCRT蛋白复合物形成的进一步验证,对瞬时表达的myc-Endofin进行联合免疫沉淀。 事实上,内啡肽与HD-PTP、ESCRT-0(Hrs和STAM2)和ESCRT-I(Tsg101和UBAP1)形成复合物( 图1 B) 其中STAM2、Tsg101和UBAP1直接与HD-PTP交互( Ali等人。, 2013 ; Gahloth等人。, 2016 ; Ichioka等人。, 2007 ). 该数据进一步支持了内吞是早期内吞体上含有HD-PTP和ESCRT的复合物的一部分。
最后,我们使用MiniTurbo BioID探讨了EGFR激活是否引发Endofin募集,与BirA*酶相比,MiniTurbo BioID可以缩短标记时间。 结果表明,在EGF刺激下,内皮素和HD-PTP与常见的内吞机制一起迅速被招募到EGFR中( 图1 C) ●●●●。 总之,这些数据表明,内啡肽与HD-PTP、ESCRT和EGFR形成复合物,表明内啡肽可能在调节受体贩运中发挥作用。
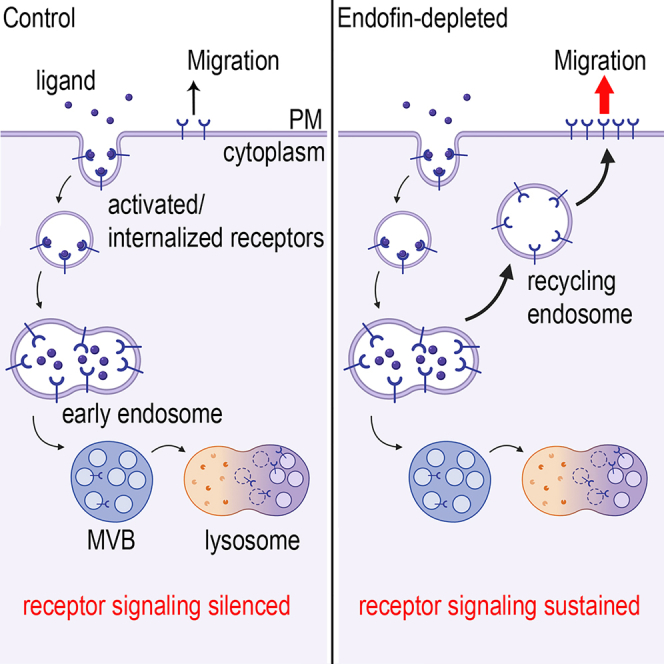
内皮素调节整合素α5和EGFR的质膜密度、稳定性和溶酶体降解 众所周知,HD-PTP在调节EGFR、整合素α5和PDGFRβ溶酶体内膜转运中起直接作用( Belle等人。, 2015 ; Doyotte等人。, 2008 ; Kharitidi等人。, 2015 ). 由于Endofin直接与HD-PTP相互作用,并与不同的ESCRT形成复合物( Gahloth等人。, 2017年b )和EGFR,我们想研究内啡肽在受体贩运中的作用。 我们使用HD-PTP-depleted HeLa细胞作为阳性对照来阻断受体贩运( 图S4 A) ●●●●。 HeLa细胞被shRNA清除Endofin-depleted,并感染非靶向shRNA(NT)作为耗尽的阴性对照( 图S4 A) ●●●●。 对Endofin和HD-PTP-depleted细胞的总细胞裂解物进行Western blotting,结果显示整合素α5的细胞表达水平至少增加了2.5倍( 图S4 B) 和~EGFR水平增加1.2–1.6倍( 图S4 C) ●●●●。
几项研究表明,配体活化、泛素化和整合素α5和EGFR的内化促进了它们向溶酶体降解方向的分选( Alwan等人。, 2003 ; Kharitidi等人。, 2015 ; Lobert等人。, 2010 ). 接下来,我们评估了与HD-PTP缺失相比,内皮素缺失对整合素α5和EGFR溶酶体降解动力学的影响。 在环己酰亚胺(CHX)存在下,用纤维连接蛋白(3和6 h)或EGF(2和4 h)刺激血清饥饿的细胞以抑制蛋白质合成。 为了抑制溶酶体降解,在CHX存在下,用巴氟霉素A1预处理细胞(1小时)。 Western blot显示,在内皮素和HD-PTP缺失后,6小时和2小时追踪后,整合素α5和EGFR的溶酶体降解分别延迟>60%和~40%( 图2 A) ,表明内皮素在整合素α5和EGFR溶酶体降解中发挥作用。
图2。
内皮素介导整合素α5、EGFR和CD4多泛素化货物模型对溶酶体降解的有效分选
(A) Endofin-和HD-PTP-depleted HeLa细胞中总受体水平的整合素α5(左面板)和EGFR(右面板)环己酰亚胺追踪(10μg/mL CHX)。 细胞需要血清(2 h),用CHX预处理(1 h),然后用纤维连接蛋白(10μg/mL FN,37°C)或EGF(50 ng/mL,37°C)激活受体。 整合素α5被追踪0、3和6小时,而EGFR在CHX存在下追踪0、2和4小时。 对照细胞也用Bafilomycin A1(200 nM Baf+CHX,1 h)预处理,以抑制溶酶体酸化,从而抑制溶酶体降解(6 h FN+Baf或4 h EGF+Baf)。 利用ImageJ软件对细胞提取物进行蛋白质印迹,通过密度分析测定降解动力学。 整合素α5和EGFR水平归一化为eEF2。
(B) 用细胞表面ELISA(cs-ELISA)测定HeLa细胞中整合素α5(左侧)和EGFR(右侧)质膜(PM)的稳定性。 血清饥饿后,用纤维连接蛋白(10μg/mL FN,4 h,37°C)或EGF(50 ng/mL,20 min,37°C)激活受体。 α5β1整合素阻断抗体(10μg/mL)和吉非替尼(2μM)作为阴性对照,阻断Endofin-depleted细胞的受体内化。 对于每个细胞系,整合素α5和EGFR的水平绘制为细胞表面与未刺激细胞相比剩余的百分比。
(C) 在缺乏血清的HeLa细胞上进行cs-ELISA,以量化稳定状态下整合素α5(左侧面板)和EGFR(右侧面板)PM受体密度。 将内皮素和HD-PTP-depleted细胞与对照NT细胞进行比较。
(D) (上面板)受体转运期间囊泡pH值变化的示意图(EE:早期内体,RE:再循环内体,LE:晚期内体,Lys:溶酶体)。 (下面板)血清饥饿HeLa细胞中EGFR和整合素α5内吞动力学的FRIA分析。 用FRIA测定内皮素、HD-PTP-和Hrs-depleted HeLa细胞与NT细胞中FITC-标记的EGFR-或整合素α5的内吞小泡的平均pH值。 血清饥饿后,用EGF(50 ng/mL,30或60 min,37°C)或FN(10μg/mL,4 h,37°C)刺激细胞。
(E) 用CD4Tl(溶酶体降解受体分选阴性对照)或CD4Tl-泛素嵌合体(CD4T1-Ub,作为多泛素化货物的模型)瞬时转染内皮素和HD-PTP-depleted 293T细胞。 用CHX(100μg/mL)预处理血清饥饿的293T细胞,以追踪CD4Tl和CD4Tl-Ub的总蛋白水平2 h。用ImageJ软件对细胞提取物进行蛋白质印迹(CD4抗体),以通过密度分析测量降解动力学。 CD4Tl和CD4Tl-Ub水平归一化为eEF2。
(F和G)(F)在15、30、60和120分钟追踪(37°C)后,通过FRIA测定Endofin-depleted HeLa细胞与NT细胞中含有CD4Tl-Ub-和(G)CD4TCC-UbAllRΔG的内吞小泡的平均小泡pH值。 CD4TCC UbAllRΔG作为四聚体单泛素化货物的模型。 数据为n≥3个独立实验的平均值±SEM。 未配对学生t检验:*p<0.05,*p<0.01,***p<0.001。
考虑到ESCRT和泛素化跨膜蛋白之间的串扰,这些蛋白优先针对溶酶体降解,而不是再循环回PM,我们询问Endofin耗竭是否可以延迟活化受体的PM周转。 通过细胞表面ELISA(cs-ELISA)分别在纤连蛋白(4h)和EGF(20min)刺激下测量活化的整合素α5和EGFR的细胞表面稳定性。 与NT细胞相比,约20%的整合素α5在PM处稳定,类似于整合素β5β1阻断抗体的作用,该抗体干扰受体的激活和内化( 图2 B) ●●●●。 类似地,内皮素耗竭后,约20%的EGFR在PM稳定。 吉非替尼(一种EGFR抑制剂,可阻断受体激活、泛素化和内化)的作用类似( 图2 B) ●●●●。 值得注意的是,内皮素和HD-PTP耗竭对EGF刺激后EGFR的内化率没有影响(5分钟)( 图S4 D) ●●●●。 此外,我们使用cs-ELISA测量了内毒素和HD-PTP耗竭后的稳态PM受体密度。 整合素α5和EGFR细胞表面密度比NT细胞增加至少1.5倍( 图2 C) ●●●●。 因此,内皮素耗竭可能会延迟整合素α5和EGFR的溶酶体降解,并通过促进早期内体水平的再循环增加其PM密度和稳定性。
内酯是有效溶酶体传递整合素α5和EGFR所必需的 如果整合素α5和EGFR的溶酶体降解在内皮素耗竭后延迟,则内皮素可能在ESCRT依赖性受体从早期内体向MVB和溶酶体的分选中发挥作用。 因此,为了更好地了解内啡肽在溶酶体内转移动力学中的作用,通过监测含受体小泡的pH值(pH值)来确定溶酶体的受体传递 v(v) )通过荧光比率图像分析(FRIA)( 图2 D)( Barriere和Lukacs,2008年 ). HD-PTP和Hrs-depleted HeLa细胞( 图S4 A) 被用作阻断受体贩运的积极控制( Kharitidi等人。, 2015 ; Wenzel等人。, 2018 ). 首先,在冰上用抗EGFR或抗整合素α5标记缺乏血清的细胞,然后用F(ab')标记 2 与pH敏感的异硫氰酸荧光素(FITC)偶联的二级抗体。 接下来,为了诱导同步的受体内化,用EGF(30和60分钟追逐,37°C)或纤维连接蛋白(4小时追逐,37℃)和pH值刺激细胞 v(v) 如预期,在HD-PTP和Hrs-depleted细胞(pH值 v(v) ~5.7±0.09和pH v(v) ~5.7±0.1),与NT细胞(pH v(v) ∼ 5.2 ± 0.07) ( 图2 D和 S4系列 E) ●●●●。 引人注目的是,在内皮素缺失细胞中追踪30分钟EGFR后,受体局限于早期内体(pH v(v) ~6±0.17),然而,在NT细胞中,EGFR被传递到晚期内体(pH v(v) ∼ 5.4 ± 0.07) ( 图2 D和 S4系列 E) ●●●●。 此外,60分钟追踪后,EGFR被传递到Endofin-depleted细胞(pH v(v) ~5.4±0.23),而在NT细胞中,EGFR被传递到溶酶体(pH v(v) ∼ 4.5 ± 0.1) ( 图2 D和 S4系列 E) ●●●●。 内皮素耗竭也抑制了整合素α5从早期内体(pH值 v(v) ~6.4±0.1),而在对照NT细胞中,受体被传递到晚期内体(pH v(v) ∼ 5.7 ± 0.1) ( 图2 D) ●●●●。 FRIA结果清楚地表明,内啡肽在受体从早期内体向MVB和溶酶体的转运中起着关键作用。 HD-PTP和Hrs耗竭对EGFR贩运的相同影响就是一个例证。
内酯是多泛素模型货物溶酶体高效输送所必需的 ESCRT对泛素化受体的识别是内化受体向MVB和溶酶体分选过程中的关键步骤。 为了检查Endofin是否也有助于不能发出受体信号的泛素化货物的内溶酶体分选,我们使用了已建立的CD4-Ub嵌合体,该嵌合体经历组成型多泛素化。 为此,使用了两种不同的模型受体。 一个是截断的CD4(CD4Tl),其中细胞质尾部被删除并替换为连接子,因此缺少泛素受体位点和分选信号( Barriere等人。, 2006 ). 此外,我们使用了CD4Tl-Ub,这是一种嵌合体,其中连接子已融合到泛素(Ub)部分。 单个Ub部分与连接子的融合足以诱导CD4Tl-Ub的组成性多泛素化,进而加速其依赖于ESCRT的溶酶体递送( Apaja等人。, 2010 ; Barriere等人。, 2007 ).
用CD4Tl和CD4Tl-Ub构建体瞬时转染Endofin-和HD PTP缺失的细胞,以通过CHX-chase评估其降解动力学(2小时)。 Western blotting表明,与NT细胞相比,内切芬和HD-PTP耗竭后,~40%CD4Tl-Ub的溶酶体降解延迟( 图2 E) ●●●●。 正如预期的那样,内啡肽和HD-PTP缺乏对CD4Tl水平没有显著影响( 图2 E) ●●●●。 此外,FRIA显示,内皮素耗竭将CD4Tl-Ub限制在再循环的内皮体内(pH v(v) ~6.4±0.07)与NT细胞相比,NT细胞将其输送至溶酶体(pH v(v) ~4.8±0.07)追逐2小时后( 图2 F) ●●●●。 因此,即使在缺乏第二信使信号活动的情况下,内啡肽也是多-泛素化货物溶酶体高效降解所必需的。 为了评估内啡肽依赖的内溶酶体分选是否优先于特定的泛素化产物,FRIA还测定了四聚体单-泛素化物(CD4TCC-UbAllRΔG)在内啡肽耗尽时的溶酶体递送。 在CD4TCC-UbAllRΔG中,在Ub部分之前插入一个四聚体信号,Ub部分的所有七个Lys残基被Arg取代,羧基末端Gly残基被删除( Barriere等人。, 2006 ). 与HD-PTP缺失对CD4TCC-UbAllRΔG贩运的影响一致( Kharitidi等人。, 2015 ),内啡肽的消耗并不影响四聚体单-泛素化货物的贩运( 图2 G) 尽管之前已经证明四聚体单-双肽可以作为一种有效的内吞信号( Barriere等人。, 2006 ). 这些数据表明,内啡肽不仅在EGFR和整合素α5的贩运中发挥作用,而且还调节多-泛素化货物向溶酶体降解的特定分类。
内啡肽耗竭减少氯氰菊酯向早期内体的补充 Endofin通过Tom1向早期内体招募网格蛋白( Seet和Hong,2005年 )Tom1直接与Endofin和clathrin重链相互作用( Seet和Hong,2005年 ; Seet等人。, 2004 ) ( 图S5 C) ●●●●。 我们的数据表明,在15分钟EGFR追踪后,氯氰菊酯与EGFR共定位达到峰值(~0.65±0.09)( 图S5 A和S5B)。 为了更好地理解为什么内皮素耗竭后受体贩运会延迟,我们首先通过免疫荧光法评估了内皮素耗损对早期内体中氯氰菊酯补充的影响。 用GTPase缺陷突变体Rab5Q79L瞬时转染内切酶缺失的HeLa细胞,以诱导扩大的内切体的形成( Raiborg等人。, 2001 ; Wegener等人。, 2010 ). 内皮素耗竭后,与NT细胞相比,网格蛋白募集减少了33%±3%( 图S5 D和S5E)。 此外,我们评估了在Endofin耗竭的细胞中Tom1与网格蛋白重链的相互作用,co-IP显示,网格蛋白与Tom1的相互作用在Endofin耗竭后减少( 图S5 F) ●●●●。 因为在MVB生物发生过程中,氯菊酯对于内胚体表面的受体聚集及其在ILV中的积累是必不可少的( Wenzel等人。, 2018 ),这可能导致EGFR溶酶体传递延迟。 如Seet等人之前所述,为了模拟Tom1蛋白通过Endofin向早期内体募集。, 我们使用2xFYVE-Tom1嵌合体构建来恢复内皮素缺失细胞中Tom1/网格蛋白的相互作用( Seet和Hong,2005年 ). 值得注意的是,在Endofin-depleted细胞中的2xFYVE-Tom1嵌合体一旦被表达,就缺乏与Endofin结合伙伴形成复合物的能力,包括HD-PTP。 我们的结果表明,用2xFYVE-Tom1嵌合物补充Endofin-depleted HeLa细胞无法恢复EGFR溶酶体递送( 图S5 G) ●●●●。 因此,尽管内啡肽耗竭减少了与Tom1的氯氰菊酯相互作用,并减少了氯氰菊酯向早期内体的补充,但内啡肽耗尽后EGFR溶酶体递送的延迟与内啡肽和Tom1之间相互作用的丧失或Tom1在受体贩运中的作用没有直接联系。
内皮素表达增强HD-PTP与EGFR、Hrs和CHMP4B的共定位 HD-PTP直接与ESCRT-0、-I和-III蛋白相互作用,促进EGFR向MVB的分选和溶酶体降解( Doyotte等人。, 2008 ; Kharitidi等人。, 2015 ). Endofin与HD-PTP的Bro1结构域结合,该结构域与STAM2(ESCRT-0)和CHMP4B(ESCRT-III)具有相同的结合位点,然而,Endofin结合HD-PTP具有比STAM2和CHMP4 B更高的亲和力( Gahloth等人。, 2017年b ). 此外,STAM2还与HD-PTP富含脯氨酸区域的基序结合,该基序是STAM2相互作用的第二个位点( Ali等人。, 2013 ). 为了研究Endofin是否可以通过ESCRT调节HD-PTP复合物的形成,在Endofin-depleted细胞中免疫沉淀内源性HD-PTP,并评估ESCRT-0蛋白Hrs的co-IP效率。 内皮素耗竭后,HD-PTP与Hrs的间接相互作用减少了约80%( 图3 A) ●●●●。
图3。
Endofin促进HD-PTP与EGFR、Hrs和CHMP4B的共定位
(A) 在EGF刺激下(50 ng/mL,15 min,37°C),对对照组和Endofin-depleted 293T细胞进行Co-IP。 对内源性HD-PTP进行免疫沉淀,并通过蛋白质印迹评估Hrs的共IP。 加载全细胞裂解液(WCL 1%)以显示蛋白质含量。
(B–D)(B和C)利用HD-PTP对Endofin-depleted HeLa细胞中的EGFR(B)、Hrs(C)和CHMP4B(D)进行Colocalization分析。 用EGF(50 ng/mL,5 min,37°C)刺激血清饥饿的细胞,然后在37°C下追踪EGFR 0,10,20和30 min。 使用ImageJ软件(n=30)量化Mander的结肠化系数(MCC)。 每个时间点都显示有代表性的免疫荧光图像。 数据为n≥3个独立实验的平均值±SEM。 未配对学生t检验:*p<0.05,*p<0.01,***p<0.001。
由于HD-PTP与早期内体上的ESCRT-0和-III的相互作用对于活化EGFR向MVB和溶酶体的分选至关重要( Ali等人。, 2013 ) ( Doyotte等人。, 2008 ) ( Wenzel等人。, 2018 ),我们通过脉冲相实验评估了EGFR、红色荧光蛋白标记的Hrs(RFP-Hrs)和mCherry标记的CHMP4B(mCherry-CHMP4B)与HD-PTP在Endofin-depleted细胞中的共定位。 与NT细胞(0.54±0.03)相比,经5分钟EGF刺激和10分钟EGFR追踪后,内皮素耗竭后EGFR与HD-PTP共定位显著降低(0.4±0.03( 图3 B) ●●●●。 与co-IP结果一致,在5分钟EGF刺激和0分钟EGFR追踪后,与NT细胞(0.71±0.03)相比,内皮素耗竭后与HD-PTP共定位的Hrs显著降低(0.44±0.01)( 图3 C) ●●●●。 此外,在5分钟EGF刺激和20分钟EGFR追踪后,与NT细胞(0.62±0.05)相比,内皮素耗竭后CHMP4B与HD-PTP的共定位也降低了(0.44±0.06)( 图3 D) ●●●●。 因此,该数据表明,内皮素促进EGFR、ESCRT-0(Hrs)和ESCRT-III(CHMP4B)与HD-PTP的共定位和/或复合物形成,从而通过ESCRT依赖机制促进EGFR向MVB的分选和溶酶体降解。
接下来,还通过脉冲追逐实验在HD PTP耗竭的细胞中评估EGFR和RFP-Hrs与Endofin的共定位。 我们发现,在5分钟EGF刺激和20分钟EGFR追踪后HD-PTP耗竭后,EGFR和Hrs与Endofin的共定位稳定。 然而,EGFR和Hrs在NT细胞中与Endofin动态共定位( 图S6 A和S6B)。 这表明EGFR和Hrs与Endofin的动态共定位受HD-PTP调节。 这种动态共定位可能反映了货物从早期内体向MVB和溶酶体的出口,这确实受到HD-PTP的调节( Doyotte等人。, 2008 ). 另一方面,当HD-PTP耗尽时,EGFR和Hrs与Endofin的稳定共定位可能反映了从早期内体到MVB和溶酶体的货物分拣延迟。
内啡肽/HD-PTP相互作用可以有效地通过溶酶体传递活化的EGFR 内吞直接结合HD-PTP并将其招募到早期内吞体( Gahloth等人。, 2017年b ). L202D/I206D-HD-PTP双突变消除了CHMP4B和Endofin与HD-PTP的结合,即使在L202D/I206D-HD-PTP与myc Endofin共表达时,也能阻止L202D/I206D-HD-PTP募集到早期内体( Gahloth等人。, 2017年b ). 我们的数据表明,EGFR和ESCRT-0与HD-PTP共定位被Endofin稳定。 为了评估Endofin/HD-PTP相互作用在调节向MVB和溶酶体的货物分拣方面的功能,我们生成了shRNA抗性野生型(WT)和突变Endofin构建物。
FYVE域C753S-Endofin突变体的瞬时过表达显示了对早期内体的定位受损,我们在这里也发现了这一结果( 图S6 C)( Seet和Hong,2001年 ). 削弱内啡肽与HD-PTP Bro1结构域的相互作用( Gahloth等人。, 2017年a ),我们采用L15P-内啡肽突变体。
研究内皮素互补对EGFR内溶酶体转运、pH值的影响 v(v) FRIA监测含EGFR的囊泡。用WT-或突变的Endofin瞬时转染Endofin-depleted HeLa细胞( 图S6 D) 和平均pH值 v(v) 在EGF刺激30分钟后进行评估。 WT-Endofin互补恢复EGFR溶酶体递送(平均pH v(v) ~5.1±0.03)与模拟转染的Endofin-depleted细胞(平均pH值 v(v) ~5.9±0.09)和NT细胞(平均pH v(v) ∼ 5.3 ± 0.04) ( 图4 A) ●●●●。 L15P-内啡肽互补部分恢复EGFR溶酶体递送(平均pH v(v) ~5.5±0.02),与WT-Endofin相比( 图4 A) 与L15P-Endofin与HD-PTP的部分生物化学相互作用被破坏相一致( 图S6 E) ●●●●。 相反,C753S-内啡互补完全不能挽救EGFR溶酶体递送(平均pH v(v) ∼ 5.9 ± 0.05) ( 图4 A) 通过大幅减少早期内吞体对内吞素的补充。
图4。
内啡肽/HD-PTP相互作用可以有效地通过溶酶体传递活化的EGFR
(A和B)在(A)与mCherry质粒和Flag-tagged WT-、L15P-或C753S-Endofin构建物瞬时共转染的Endofin-depleted HeLa细胞中,在EGFR追踪30分钟后(50 ng/mL EGF,37°C),由FRIA测定FITC-标记的EGFR-内吞小泡的平均小泡pH值,或(B) HD-PTP-depleted HeLa细胞与mCherry质粒、WT-或T145K-HD-PTP瞬时共转染(干扰HD-PTP与Endofin的相互作用)构建物。 对表达mCherry的细胞进行单细胞FRIA分析(囊泡:n>190,显微镜视野:n>10)。 绘制了EGFR追踪30分钟后内化EGFR的水泡pH分布。 结果表明,NT、shEndofin、WT-Endofin、L15P-Endofen、C753S-Endofein、shHD-PTP-和T145K-HD-PTP-HeLa细胞中不同峰的平均囊泡pH值和囊泡数量。
(C) BioID屏幕的点图显示与泛素化(Ub)和早期内体(EE)相关的蛋白质与WT-Endofin和C753S-Endofin非常接近。 将标记生物素连接酶(BirA*)融合到WT-Endofin和C753S-Endofin的N末端,并用作诱饵。 将构建物转染到Flp-In T-REx 293T细胞中。 点图中的填充阴影表示平均光谱计数(Avg Spec),点的大小表示所有猎物的相对丰度,外圆颜色表示BFDR值。 数据为n≥3个独立实验的平均值±SEM。 未配对学生t检验:*p<0.05,*p<0.01,***p<0.001。
重要的是,用WT-HD-PTP补充HD-PTP-depleted HeLa细胞( 图S6 D) 恢复EGFR溶酶体输送(平均pH v(v) ∼ 5.4 ± 0.15) ( 图4 B) ●●●●。 T145K-HD-PTP,部分破坏HD-PTP/内啡肽相互作用( 图S6 F)( Gahloth等人。, 2017年c ),未能挽救EGFR溶酶体输送(平均pH v(v) ~5.9±0.05)与模拟转染HD-PTP-depleted(平均pH v(v) ~6.1±0.08)和NT(平均pH v(v) ~5.4±0.09)个电池( 图4 B) ●●●●。
为了更好地理解Endofin的FYVE结构域影响HD-PTP向早期内体募集的机制,我们将我们的BioID研究扩展到C753S-Endofin-N-末端BirA*标记毒饵。 与我们的研究结果一致,FYVE结构域的破坏并没有减少内皮素与HD-PTP的相互作用,但确实破坏了其与泛素和一些早期内体成分的邻近相互作用, 深入了解内吞素支持HD-PTP在内吞囊泡上向泛素化货物募集的潜在机制( 图4 C) ●●●●。 这些结果强烈表明,通过FYVE结构域在早期内体上定位内啡肽及其与HD-PTP的相互作用对于有效的激活EGFR内溶酶体转移是必要的。
内皮素耗竭维持整合素α5和EGFR下游信号 我们之前的研究表明 PTPN23型 常见于几种人类癌症( Manteghi等人。, 2016 ). 使用cBioPortal数据库,我们在这里显示 ZFYVE16型 (内啡肽基因)在几种人类癌症中也很常见( 图5 A) ●●●●。 此外,PRECOG数据库通过计算Z评分来衡量mRNA表达谱与癌症预后之间的关系,结果显示Z评分为负 ZFYVE16型 几种人类癌症的mRNA表达( 图5 B) ●●●●。
图5。
内皮素耗竭维持整合素α5和EGFR下游信号传导并增加细胞迁移
(A) 杂合性缺失频率 ZFYVE16型 不同类型癌症患者的(内啡肽基因)。 从cBioPortal数据库中提取数据并绘制为 ZFYVE16型 杂合性丢失。
(B) 不同类型癌症患者内啡肽Z评分分析。 从PRECOG数据库中提取并绘制每种癌症类型的Endofin Z评分。
(C) Western blot显示在纤维连接蛋白刺激后(10μg/mL FN,7 h,37°C),内皮素和HD-PTP-depleted HeLa细胞与NT细胞中整合素α5受体下游效应器的磷酸化水平:FAK、Src和Erk1/2。 eEF2螺栓用作加载控制。
(D) 与NT细胞相比,用EGF(5 ng/mL,37°C)刺激内皮素和HD-PTP-depleted HeLa细胞0、15、30和60分钟。进行蛋白质印迹以揭示EGFR(左面板)及其下游效应物MEK(右面板)的磷酸化状态。 eEF2印迹用作加载对照。
(E) 实时细胞分析(RTCA)测量内皮素和HD-PTP缺失HeLa细胞与NT细胞的细胞迁移。 添加10%FBS的细胞培养基被用作化学引诱剂来触发细胞迁移。 使用无血清培养基(不含化学引诱剂)作为阴性对照。 使用整合素α5β1阻断抗体(10μg/mL)作为阴性对照,以抑制Endofin-depleted细胞中的细胞迁移。
(F) 用非靶向(NT)shRNA或HD-PTP shRNA瞬时转染Endofin-depleted或对照HeLa细胞。通过Western blotting评估受体水平,并使用ImageJ软件通过密度分析进行量化。
(G) 如(E)所示进行实时细胞分析,以比较瞬时转染HD-PTP shRNA或NT对照的Endofin-depleted细胞的迁移率。 数据为n≥3个独立实验的平均值±SEM。 未配对学生t检验:*p<0.05,*p<0.01,***p<0.001。
细胞表面受体能够在ILV形成之前从早期内体或MVB发出信号( Mamiñska等人。, 2016 ; Rodahl等人。, 2009 ; Wegner等人。, 2011 ). 此外,HD-PTP缺失可延迟整合素α5溶酶体的传递,从而增加其下游信号传导( Kharitidi等人。, 2015 ). 为了研究内皮素是否也影响整合素α5信号传导,用纤维连接蛋白刺激内皮素和HD-PTP-depleted HeLa细胞(7 h),通过Western blotting评估磷酸化FAK、Src和Erk1/2的水平。 与我们之前的研究一致,在稳态和纤维连接蛋白刺激下,与NT细胞相比,缺乏血清的细胞中HD-PTP的缺失增加了pFAK、pSrc和pErk1/2的水平( 图5 C) ●●●●。 值得注意的是,在内皮素耗竭时也观察到了同样的效果( 图5 C) ●●●●。 这一结果清楚地表明,除了受体稳定外,Endofin耗竭还进一步增加了整合素α5的信号传导。
接下来,评估Endofin和HD-PTP耗竭对EGFR激活的影响及其下游信号传导的动力学。 有趣的是,内皮素和HD-PTP的缺失对稳态EGFR磷酸化及其下游激酶MEK的激活没有显著影响( 图5 D) ●●●●。 然而,在内皮素和HD-PTP耗竭后,EGF刺激30分钟后,EGFR和MEK均持续激活。 即使在EGF刺激60分钟后,这些细胞中的pEGFR水平仍然较高( 图5 D) ●●●●。 总之,这些结果表明,内啡肽和HD-PTP不仅调节EGFR和整合素α5质膜稳定性,而且还控制受体激活及其下游信号传导。
内皮素耗竭增加细胞迁移 整合素α5β1在细胞迁移中的中心作用及其依赖于ESCRT的溶酶体脱敏已得到充分证实( Kharitidi等人。, 2015 ; Lobert等人。, 2010 ). 采用实时细胞分析(RTCA)方法,以胎牛血清为化学引诱剂,测定了Endofin-depleted HeLa细胞的迁移率。 与NT细胞相比,内皮素耗竭对细胞迁移率的影响与HD-PTP耗竭的影响相同,增加了约2倍。 整合素α5β1阻断抗体抑制内皮素缺失细胞的迁移( 图5 E) ●●●●。 这表明,内皮素耗竭后细胞迁移的增加与PM和总整合素α5水平的增加有关( 图2 C和 S4系列 B) ●●●●。
由于Endofin/HD-PTP相互作用可以有效地通过溶酶体传递活化的EGFR( 图4 B和4C),我们接下来讨论了同时消耗内啡肽和HD-PTP是否会进一步削弱受体贩运。 因此,我们比较了用非靶向shRNA或靶向HD-PTP的shRNA瞬时转染NT和Endofin-depleted细胞的总EGFR和整合素α5水平。 我们发现,与NT细胞相比,耗尽Endofin或HD-PTP使EGFR总水平分别增加了约2倍和约1.5倍,而双敲除细胞中的EGFR水平增加了约2.7倍( 图5 F) ●●●●。 有趣的是,对整合素α5的影响更为显著。 事实上,单独消耗内皮素或HD-PTP使整合素α5水平分别增加了约3.4倍和约2.7倍,而双敲除细胞与NT细胞相比增加了约7.7倍( 图5 F) ●●●●。 为了解决这些发现的功能相关性,我们还使用胎牛血清作为化学引诱剂进行了迁移分析。 结果表明,与Endofin-depleted细胞相比,双敲除细胞的迁移率增加了约1.5倍( 图5 G) ●●●●。 总之,这些结果表明,内啡肽和HD-PTP的同时消耗对总受体水平有加性影响,这反过来又增加了它们相对于内啡肽缺失细胞的迁移率。
在某些类型的人类癌症中,整合素α5和EGFR是众所周知的肿瘤发生驱动因素。 在这里,我们表明,内皮素缺乏增加了整合素α5和EGFR信号和细胞迁移,这是癌症进展的两个标志。 因此,Endofin表达的缺失延迟了受体溶酶体的降解,并增加了受体信号传导和细胞迁移,这可能最终导致肿瘤发生和癌症进展。
STAR★方法 关键资源表
试剂或资源 来源 标识符
抗体
内啡肽 蛋白质技术 13118-2-美联社
HD-PTP(HD-PTP) 蛋白质技术 10472-1-美联社
Myc-Tag公司 细胞信号 2276
高分辨率分光计 蛋白质技术 10390-1-美联社
STAM公司 蛋白质技术 12434-1-美联社
Tsg101型 蛋白质技术 14497-1-美联社
UBAP1公司 蛋白质技术 12385-1-美联社
电压37A 蛋白质技术 11870-1-美联社
CHMP4B型 蛋白质技术 13683-1-美联社
用于WB和IP的EGFR(A-10) 圣克鲁斯 钪-373746
EGFR(ICR10)用于cs-ELISA、IF和FRIA 阿布卡姆 抗体231
用于WB的整合素a5 蛋白质技术 10569-1-美联社
整合素a5用于cs-ELISA和FRIA BD制药 555651
抗人整合素a5b1 Millipore公司 1969年5月
用于WB的CD4(H370) 圣克鲁斯 Sc-7219号
用于FRIA的CD4(OKT4) Invitrogen公司 14-0048-82
标记标签(M2) 西格玛 1804层
电子EF2 细胞信号 2332
a-管蛋白 西格玛 T6074型
氯氰菊酯(X22) 阿布卡姆 Ab2731号
欧洲经济区1 Invitrogen公司 14-9114-82
抗大鼠HRP F(ab')2 杰克逊免疫研究 712-036-153
抗鼠HRP F(ab')2 杰克逊免疫研究 115-036-003
抗鼠FITC F(ab')2 杰克逊免疫研究 712-096-150
防鼠AF488 Invitrogen公司 A-28175号
抗鼠AF594 Invitrogen公司 A-11032号
抗兔AF488 Invitrogen公司 A-27034号
抗兔AF594 Invitrogen公司 A-11012号文件
抗鼠AF647 Invitrogen公司 A-21247号
化学品、肽和重组蛋白质
重组人表皮生长因子 吉布科 PHG0311电话
人血浆纤维结合蛋白 吉布科 33016-015
环己酰胺 西格玛 66-81-9
巴氟霉素A1 西格玛 B1793号
CCCP公司 西格玛 C2759元
莫能新钠 西格玛 M5273型
尼日利亚菌素钠盐 西格玛 N7143号
Amplex红色 Invitrogen公司 A12222号
存放的数据
AP-MS数据 https://massive.ucsd.edu/ProteoSAFe/static/massive.jsp 质量:MSV00087704
BirA BioID MSPLIT数据 https://massive.ucsd.edu/ProteoSAFe/static/massive.jsp 质量:MSV00087705
MiniTurbo BioID MSPLIT数据 https://massive.ucsd.edu/ProteoSAFe/static/massive.jsp 质量:MSV00087706
实验模型:细胞系
赫拉NT 暂停实验室
HeLa sh54内啡肽 暂停实验室
HeLa sh75内啡肽 暂停实验室
希拉sh47HD-PTP 暂停实验室
希拉sh51HD-PTP 暂停实验室
希拉sh98小时 暂停实验室
293T北卡罗来纳州 暂停实验室
293T sh54内啡 暂停实验室
293T sh75内啡 暂停实验室
293T sh47HD云台 暂停实验室
寡核苷酸
克隆和诱变引物的详细信息见 表S1
重组DNA
pcDNA3-Flag-Endofin公司 暂停实验室
pcDNA3-Flag-内啡抗shRNA54 暂停实验室
pcDNA3-Flag-L15P-抗shRNA54的内啡肽 暂停实验室
pcDNA3-Flag-C753S-内啡抗shRNA54 暂停实验室
pcDNA3标志HD PTP 暂停实验室
pcDNA3-Flag-HD-PTP抗shRNA47 暂停实验室
pcDNA3-Flag-T145K-HD-PTP抗shRNA47 暂停实验室
pcDNA3-CD4Tl 卢卡奇实验室
pcDNA3-CD4Tl-Ub 卢卡奇实验室
pcDNA3-CD4TCC-UbAllRΔG 卢卡奇实验室
软件和算法
MetaMorph/MetaFluor软件 分子器件
原始软件 原始实验室 https://www.originlab.com/
ImageJ软件 https://imagej.nih.gov/ij/
资源可用性 材料可用性 本研究中产生的质粒和细胞系可通过铅接触获得。
实验模型和受试者详细信息 细胞系 内皮素和HD-PTP-depleted HeLa和293T细胞是从ATCC购买的野生型细胞中生成的。 本研究中产生的所有细胞系均列于 密钥资源表 并可根据要求从潜在联系人处获得。
方法详细信息 构筑物、试剂和细胞培养 Myc-tagged Endofin是王博士送的一份礼物( Seet和Hong,2001年 ). 以myc-Endofin为模板,通过PCR扩增产生pcDNA3-Flag-Endofin。 pcDNA3-Flag-Endofin对shRNA54具有抗性,通过诱变在shRNA靶向区域引入沉默点突变产生。该构建物用于创建从BioBasic获得的C753S和L15P Endofin突变体。 先前Gingras等人描述了pcDNA3.1中的标记HD-PTP。 ( Gingras等人。, 2009 ). pcDNA3-Flag-HD-PTP对shRNA47产生抗性,通过诱变在shRNA靶区引入沉默突变。 进一步对该结构进行突变,获得T145K HD-PTP突变体(BioBasic)。 FYVE-Tom1嵌合体是在pcDNA3-Flag载体中通过子克隆Endofin FYVE结构域编码区的串联重复序列以及Tom1(Biomatik)的连接子和编码序列而生成的。 如Barriere et al。 ( Barriere等人。, 2006 ). pCS2-RFP-Hrs是E.M.De Robertis博士的礼物(Addgene质粒#29685)。 pLNCX2-mCherry-CHMP4B是S.Simon博士的礼物(Addgene质粒#116923)。 引物的完整列表包含在 表S1 .
HeLa(ATCC®CCL-2™)和293T(ATCC™CRL-3216™)细胞在DMEM、10%FBS(Wisent)中培养。 人类内啡肽MISSION®shRNA慢病毒质粒pLKO.1-puro(克隆ID: NM_014733号 .2-4263s1c1(第54页), NM_014733号 .3-3858s21c1(sh75)),人类HD-PTP(克隆ID: NM_015466号 .x-571s1c1(第47页), NM_015466号 .x-887s1c1(sh51)),人时(克隆ID: NM_004712号 .3-494s1c1(sh98))和空载体(MISSION®pLKO.1-puro空载体控制质粒DNA,SHC001)购自Sigma。
本研究中使用的抗体的完整列表包括在 密钥资源表 进行了补充实验 通过 根据制造商的说明,使用Lipofectamine 2000(Invitrogen)进行瞬时转染。 本研究中使用的试剂的完整列表包括在 关键资源表 .
AP-MS克隆、稳定细胞系生成和样品采集 通过Gateway克隆将ORF转移到N末端3XFLAG哺乳动物表达载体中,用于等基因稳定细胞系的生成和四环素诱导表达。 根据制造商的说明,使用脂质体2000(Invitrogen)以6孔格式转染Flip-In T-REx 293T细胞,其中含有0.2μg标记DNA(pcDNA5-FLAG蛋白)和2μg pOG44(OpenFreezer V4134)。 第2天,对细胞进行胰蛋白酶处理,并传代到10 cm的平板中。 第3天,用DMEM 10%胎牛血清、100单位/ml pen/strep和200 ug/ml潮霉素替换培养基。 培养基每隔2-4天更换一次,直到非转染细胞死亡,分离的克隆直径为~1-2 mm(13-15天)。 细胞池是通过对整个培养板进行胰蛋白酶化并在新鲜选择培养基中重新培养而产生的(培养板的大小取决于初始菌落的数量和大小)。 用1μg/ml四环素诱导约60–70%融合细胞24小时。 收集次级流入细胞(~85-95%融合细胞)进行AP-MS分析。
BioID克隆、稳定细胞系生成和样品采集 Endofin和HD-PTP的构建体被克隆到5′BirA*pcDNA5 FRT TO或pcDNA5 FRT TO 3′BirA*中。 将EGFR的构建体克隆到5’(MiniTurbo pcDNA5 FRT TO或pcDNA5 FRT TO 3’MiniTurbo。使用lipofectamine 2000(Invitrogen)以6孔形式用0.2μg克隆构建体和2μg pOG44(OpenFreezer V4134)转染Flip-In T-REx 293T细胞(Endofin BirA*和HD PTP BirA*)或Flip-In T-REx HeLa细胞(EGFR MiniTurbo), 根据制造商的说明。 第2天,对细胞进行胰蛋白酶处理,并传代到10 cm的平板中。 第3天,用DMEM 10%胎牛血清、100单位/ml pen/strep和200 ug/ml潮霉素替换培养基。 每隔2-4天更换一次培养基,直到非转染细胞死亡,分离的克隆直径为~1-2mm(13-15天)。 细胞池是通过对整个培养板进行胰蛋白酶化并在新鲜选择培养基中重新培养而产生的(培养板的大小取决于初始菌落的数量和大小)。 对于BirA*实验,用1ug/ml四环素诱导细胞约60-70%的融合24小时。 第二天,向含有四环素的培养基中添加50μM生物素,并将细胞再培养24小时。 对于EGFR-miniTurbo实验,用1ug/ml四环素在缺乏生物素的培养基中诱导细胞约60-70%的融合24小时。 第2天,细胞被切换到含四环素的无血清培养基。 在血清饥饿16小时后,在存在或不存在100ng/mL EGF的情况下,向培养基中添加50μM生物素15分钟。 为了收获,细胞在冷藏PBS中刮掉,在PBS中清洗x2,造粒并在−80°C下冷冻,直到加工MS。
AP-MS和BioID样品处理和质谱分析 对于AP-MS研究,按照St-Denis等人之前的描述进行FLAG亲和纯化。 ( St-Denis等人。, 2016 )每个样品的¼通过Velos Orbitrap质谱分析。 使用激光拉拔器(程序=4;热量=280,FIL=0,VEL=18,DEL=200)在熔融石英毛细管柱(0.75μm ID,350μm OD)上形成喷嘴。 10 cm(+/-1 cm)C18反相材料(Reprosil-Pur 120 C18-AQ,3μm)通过压力弹(在甲醇中)填充在色谱柱中。 然后在缓冲液A(6μl)中对色谱柱进行预平衡,然后将其串联至NanoLC-Ultra 2D-plus HPLC系统(Eksigent),该系统耦合至配备纳米电喷雾离子源(Proxeon Biosystems)的LTQ-Orbitrap Velos(热电)。 Xcalibur 2.0下的LTQ-Orbitrap Velos仪器在数据相关模式下运行,以在MS和最多10个后续MS/MS采集之间自动切换。 缓冲液A为100%H2O,0.1%甲酸; 缓冲液B为100 ACN,0.1%甲酸。 HPLC梯度程序提供超过125分钟的乙腈梯度。 在前20分钟内,2%B的流速为400μL/min。然后将流速降至200μL/min,溶剂B的分数线性增加至35%,直至95.5分钟。 然后在5分钟内将溶剂B增加到80%,并保持该水平直到107分钟。 然后将流动相降低至2%B,直到运行结束(125分钟)。 质谱仪上数据相关采集的参数为:1个质心MS(质量范围400–2000),然后是10个最丰富离子的MS/MS。 一般参数为:激活类型=CID,隔离宽度=1 m/z,归一化碰撞能量=35,激活Q=0.25,激活时间=10毫秒。 对于数据相关采集,最小阈值为500,重复计数=1,重复持续时间=30秒,排除大小列表=500,排除持续时间=三十秒,排除质量宽度(按质量计)=低0.03,高0.03。
对于BirA*BioID研究,如St-Denis等人。 ( St Denis等人。, 2016 )每个样品的¼在TripleTOF™5600仪器(加拿大安大略省康科德市AB SCIEX)上运行。 使用激光拉拔器(Sutter Instrument Co.,型号P-2000,参数设置为热量:280,FIL=0,VEL=18,DEL=2000),从熔融石英毛细管中生成纳米喷雾发射器,内径75μm,外径365μm,尖端开口5-8μm。 使用压力注入池将C18反相材料(Reprosil-Pur 120 C18-AQ,3μm)重新悬浮在甲醇中填充纳米喷雾发射器。 样品以400nl/min的速度直接加载到75μmx10cm纳米喷雾发射器上,持续14min。 用Eksigent ekspert™Nano Ultra 1D Plus生成的乙腈梯度从色谱柱中洗脱肽,并在TripleTOF上进行分析。 梯度以200 nl/min的速度从2%乙腈和0.1%甲酸传递到35%乙腈和0.1%甲酸,线性梯度为90 min。 随后用含有0.1%甲酸的80%乙腈洗涤10分钟,并在含有0.1%甲酸的2%乙腈中再平衡15分钟。 DDA协议的总时间为140分钟。 第一次DDA扫描的累积时间为250ms,质量范围为400-1250Da。 然后对第一次DDA扫描中识别出的前20个肽进行20次MS/MS扫描,每次MS/MS扫查的累积时间为100 MS。 要求每个候选离子的电荷状态为2-4,最小阈值为每秒250计数,并使用50mDa的窗口进行隔离。 动态排除之前分析的候选离子15秒。
对于MiniTurbo BioID研究,如Hesketh等人之前所述进行链霉亲和素下拉。 ( Hesketh等人。, 2020 )每个样品的1/6在TripleTOF™6600仪器(加拿大安大略省康科德市AB SCIEX)上运行。 样品以800 nL/min的速度直接加载到平衡的HPLC柱上,并在Hesketh等人之前报告的三重TOF仪器上进行LC-MS/MS。 ( Hesketh等人。, 2020 ). 如Hesketh等人之前报告的那样,使用仪器方法设置为数据相关采集(DDA)和数据独立采集(SWATH)模式,通过两次单独的注射对样品进行分析。 ( Hesketh等人。, 2020 ).
数据相关和独立采集数据搜索 使用ProHits实验室信息管理系统(LIMS)平台存储、搜索和分析数据相关的质谱数据。 在ProHits中,WIFF文件使用WIFF2MGF转换器转换为MGF格式,使用ProteoWizard(V3.0.10702)和AB SCIEX MS Data converter(V1.3 beta)转换为mzML格式。 然后使用吉祥物(V2.3.02)和彗星(V2016.01版本2)搜索数据。 使用从NCBI获得的RefSeq数据库(第57版,2013年1月30日)中的人类和腺病毒序列对光谱进行了搜索,并补充了马克斯·普朗克研究所的“常见污染物”( http://maxquant.org/contaminants.zip )和全球蛋白质组机器(GPM; ftp://ftp.thegpm.org/fasta/cRAP/cRAP.fasta )、正向和反向序列(标记为“gi|9999”或“DECOY”)、序列标签(BirA、GST26、mCherry和GFP)和链霉亲和素,共72481个条目。 设置数据库参数以搜索胰蛋白酶裂解,允许每个肽最多2个缺失裂解位点,对于电荷为2+至4+的前体,质量耐受性为35ppm,对于片段离子的耐受性为0.15amu。 对脱酰胺天冬酰胺、谷氨酰胺和氧化蛋氨酸进行了可变修饰。 通过iProphet管道,通过TPP(Trans-Proteomic Pipeline,v.4.7 POLAR VORTEX rev 1)分析每个搜索引擎的结果。
使用MSPLIT-DIA(版本1.0( Wang等人。, 2015 ))在ProHits 4.0中实现( Liu等人。, 2016 ). 为了生成样本特异性光谱库,通过仅保留具有最佳MS-GFDB的光谱(Beta版本1.0072(2014年6月30日)),从匹配的DDA运行(BirA*)中汇集肽谱匹配(PSM)( Kim等人。, 2010 ))每个独特肽序列和前体电荷状态的概率。 设置MS-GFDB参数以搜索前体质量耐受性为50 ppm、电荷为2+–4+的胰蛋白酶裂解。 肽长度限制为8-30个氨基酸,并选择氧化蛋氨酸作为可变修饰。 对于MiniTurbo搜索,人类SWATH地图集( Rosenberger等人。, 2014 )用作搜索库。 使用targetdecoy方法,肽级错误发现率(FDR)为1%( Elias和Gygi,2007年 ). 使用NCBI RefSeq数据库(版本57,2013年1月30日)对补充了马克斯·普朗克研究所常见污染物的总共7 36241个人类和腺病毒序列进行光谱搜索( http://141.61.102.106:8080/share.cgi?ssid=0f2gfuB )和全球蛋白质组机器(GPM; http://www.thegpm.org/crap/index.html ). 然后使用光谱库通过MSPLIT与蛋白质进行肽谱匹配,其中肽由MSPLIT-DIA通过1%FDR识别,随后使用ProHits 4.0与基因匹配( Liu等人。, 2016 ).
SAINTexpress的SAINT分析(版本3.6.1( Teo等人。, 2014 ))用于对所有数据的交互进行评分。 对于AP-MS结果,将诱饵投放(每个投放两个生物复制品)与8个阴性对照(GFP-FLAG)进行比较。 对于BirA*-MSPLIT结果,将诱饵投放(各两次生物复制)与四次阴性对照投放(BirA*-FLAG和BirA**-FLAG-GFP)进行比较。 对于MiniTurbo-MSPLIT结果,将诱饵投放(每个投放两个生物复制品)与五个阴性对照投放(GFP-FLAG-MiniTurbo和FLAG-mini Turbo)进行比较。 错误发现率(FDR)≤1%的猎物(基于SAINT平均得分在生物复制品中的分布的贝叶斯估计)被视为高置信邻近性相互作用,并使用ProHits-iz-Iz-Iz-( Knight等人。, 2017 ))(prohitsviz.lunenfeld.ca)。 在ProHits-viz中,一旦一个猎物用至少一个诱饵通过了选定的FDR阈值(此处为1%),则会检索数据集中所有诱饵的所有数量值。 边缘颜色表示低于5%FDR阈值的饵-食饵近距离相互作用。
字符串分析 使用STRING数据库生成蛋白质相互作用网络( Szklarczyk等人。, 2019 ). 彩色节点表示使用0.01的BFDR截止值和0.85以上的SAINT分数从BioID屏幕识别的第一个交互因子外壳。 白色节点表示从STRING数据库中提取的交互程序的第二个外壳。 边缘代表从STRING实验和数据库中收集的蛋白质相互作用。 边缘的厚度反映了其置信度。 第二层相互作用体中最多有50个蛋白质,并且从网络中删除了唯一的相互作用体。
免疫沉淀 在缓冲液A(50 mM Tris pH 7.5,50 mM NaCl,1.5 mM MgCl)中的冰上溶解细胞 2 1 mM EDTA、0.2%triton x-100、5%甘油、1 mM DTT和cOmplete(Roche)蛋白酶抑制剂)。 蛋白质提取物以16000g在4°C下旋转10分钟。 保留一部分上清液用于SDS-PAGE和Western blot。 将剩余部分预先清除,然后在4°C搅拌下与一级抗体孵育1小时。 用sepharose(Millipore)或磁珠(BioRad)与等量的蛋白质A/G偶联,回收蛋白质-抗原复合物。蛋白质提取物与磁珠在4°C搅拌下孵育1h。 然后将珠洗3次,并用Laemmli缓冲液洗脱,然后将样品煮沸10分钟。 免疫沉淀组分和裂解产物通过SDS-PAGE和Western blotting进行分析。
尺寸排除色谱分离 细胞在冰上缓冲液(150mM NaCl、0.1%NP40、6.25mM TrisHCl ph8、2mM EDTA、0.1mM MgCl2、1mM EGTA、蛋白酶抑制剂混合物)中溶解。 首先将裂解液在16000g下离心10min,然后将上清液在100000g下离心1h。 将上清液加载到Superdex200 HPLC柱上,收集0.3ml组分。 通过SDS-PAGE和Western blot对收集的部分进行蛋白质分析。
等密度蔗糖梯度细胞分级 293T细胞在冰上用PBS清洗,在冰上刮取少量冰冷PBS。 将细胞颗粒重新悬浮在等渗缓冲液中(20mM Hepes pH7.5;150mM NaCl;1mM DTT和蛋白酶抑制剂混合物),并通过针刺(25号针10次,然后是27号针20次)逐步分解细胞。 以400g旋转裂解液以消除细胞核,将细胞溶质上清液加载在10-40%蔗糖梯度上,在Beckman Ti-55转子中以100000g离心16h。 收集组分(0.25 ml),并使用所示标记物通过Western blotting进行分析。
环己酰亚胺外壳 细胞在无血清DMEM培养基中饥饿2 h,并用环己酰亚胺(CHX,Sigma)预处理1 h(HeLa细胞为10μg/ml,293T细胞为100μg/ml)。 作为阴性对照,细胞在CHX和Bafilomycin A1(Sigma)的存在下预处理1 h,以阻止溶酶体酸化,从而抑制受体溶酶体降解。 接下来,用EGF(50 ng/ml,2和4 h)或纤维连接蛋白(10μg/ml,3和6 h)刺激细胞,并在37°C的0.5%FBS-DMEM培养基中加入CHX。 在追逐受体达指定时间点后,在冰上裂解细胞,并通过蛋白质印迹分析受体水平。
使用cs-ELISA测量细胞表面 根据Apaja等人的描述,在活细胞中进行基于细胞表面ELISA的分析。 ( Apaja等人。, 2010 ). 简而言之,细胞在无血清DMEM培养基中饥饿2小时,在冰上用整合素α5或EGFR抗体标记,并用辣根过氧化物酶(HRP)结合二级抗体(Jackson ImmunoResearch)和Amplex Red(Life Technologies)检测。 在HeLa细胞中,37°C下EGF刺激5分钟(50 ng/ml)后测量细胞内吞。 对于质膜上的受体稳定性实验,分别用纤维连接蛋白(10μg/ml,4 h)和EGF(50 ng/ml,20 min)刺激整合素α5和EGFR。
脉冲追踪实验和免疫荧光 将HeLa细胞接种在玻璃盖玻片上,并在0.5%FBS DMEM中饥饿2小时。 对于EGFR内化,在0.5%FBS DMEM中用EGF(50 ng/ml,5 min,37°C)刺激细胞。 接下来,用PBS清洗EGF,并添加新鲜的0.5%FBS DMEM培养基,以在37°C的指定时间点追踪EGFR。 将HeLa细胞用4%多聚甲醛固定15分钟,并在室温下用0.1%Triton X-100透化10分钟。 在室温下用一级抗体标记细胞1小时,然后用荧光二级抗体(Molecular probes,Invitrogen)标记并安装用于成像。 使用Plan-Apochromat 63x/NA 1.4物镜,在LSM780共焦显微镜(Carl Zeiss MicroImaging,Inc)上进行连续图像采集。
利用FRIA进行补充实验和囊泡货物跟踪 Kazan等人描述了使用FRIA测定活细胞中货物标记的囊泡pH值的方法。 和Barriere等人。 ( Barriere和Lukacs,2008年 ; Kazan等人。, 2019 ). 在冰上用一级抗体和异硫氰酸荧光素(FITC)结合二级Fab(Jackson ImmunoResearch)标记整合素α5和EGFR,并追踪指定时间。 Apaja等人在之前使用瞬时表达的CD4-Ub或CD4tCC-UbAllRΔG。 ( Apaja等人。, 2010 ). 在互补实验中,接种在6孔板中的HeLa细胞被Lipofectamine 2000(Invitrogen)与0.5μg Endofin或HD-PTP构建物以及100 ng mcherry构建物瞬时共转染。 将转染后24 h的细胞接种在玻璃盖玻片(1.5 mm厚,Fisher Scientific)上进行FRIA。FITC-右旋糖酐(10 kDa,50μg/ml,分子探针)用作溶酶体递送的对照。 右旋糖酐被内吞1小时,并在37°C下追逐2小时。 使用配备X-Cite 120Q系统(Lumen Dynamics Group Inc.)和MetaFluor软件(Molecular Devices)的蔡司观察者Z1(卡尔蔡司显微成像)测量荧光强度。
HeLa细胞迁移试验 使用xCELLigence系统(ACEA Biosciences)进行迁移分析。 30000个HeLa细胞在无血清DMEM培养基中迁移24小时至作为化学引诱剂的10%FBS培养基。 作为阴性对照,用α5β1整合素阻断抗体(10μg/ml)处理细胞。 通过绘制每个被测细胞系的实时迁移曲线的斜率来分析细胞迁移率。
信号发送实验 HeLa细胞在37°C的指定时间点用EGF(5 ng/ml)或纤维连接蛋白(10μg/ml)处理前饥饿2 h。 收集细胞并在缓冲液B(50 mM Tris pH 7.5,150 mM NaCl,1.5 mM MgCl)中的冰上溶解 2 、1 mM EDTA、1%triton x-100、5%甘油、20 mM NaF、5 mM NaPPi、1 mM-Na 三 VO(旁白) 4 、2 mM咪唑、175μg/ml PMSF和cOmplete(Roche)蛋白酶抑制剂)。 蛋白质提取物用SDS-PAGE分离并用Western blotting分析。
量化和统计分析 使用imageJ软件对蛋白质印迹和免疫荧光图像进行定量。 使用MetaFluor软件量化FRIA数据,绘制pHv峰值,并使用Origin软件进行进一步分析。 使用Microsoft Excel软件进行统计分析。 显著性通过配对双尾Student t检验确定,p值为:*<0.05、*<0.01和**<0.001的数据被视为显著。
致谢 我们要感谢Paola Blanchette博士的帮助和建议。 J.M.K拥有加拿大博士奖Fonds de Recherche Sante(FRQS)。 G.D拥有Fonds de Recherche Sante(FRQS)博士后奖。 这项工作得到了加拿大卫生研究院(CIHR)的支持,加拿大拨款245969,加拿大癌症学会研究所(CCSRI)的拨款245095。 图6 使用创建 BioRender.com网站 .
作者贡献 J.M.K.、G.D.、G.L.L.和A.P.进行了研究概念化和实验设计。 J.M.K.、G.D.和C.E.M.进行了实验和分析。 H.J.、D.K.、P.M.A.、A.R.和N.S.D.进行了实验。 J.M.K.、G.D.、A.C.G.、G.L.L.和A.P.撰写并编辑了这份手稿。
参与者信息 Gergely L.Lukacs,电子邮件: gergely.lukacs@mcgill.ca。
Arnim Pause,电子邮件: arnim.pause@mcgill.ca。
工具书类
Agromayor M.、Soler N.、Caballe A.、Kueck T.、Freund S.M.、Allen M.D.、Bycroft M.、Perisic O.、Ye Y.、Mcdonald B.ESCRT-I的UBAP1亚单位通过SOUBA结构域与泛素相互作用。 结构。 2012; 20:414–428. doi:10.1016/j.str.2011.12.013。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Akutsu M.、Kawasaki M.、Katoh Y.、Shiba T.、Yamaguchi Y.、Kato R.、Kato K.、Nakayama K.、Wakatsuki S.通过Tom1-GAT域识别泛素化货物的结构基础。 FEBS信函。 2005; 579:5385–5391. doi:10.1016/j.febslet.2005.08.076。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Ali N.、Zhang L.、Taylor S.、Mironov A.、UrbéS.、Woodman P.UBPY和ESCRT交换的招募推动了EGFR到MVB的HD-PTP相关排序。 货币。 生物.2013; 23:453–461. doi:10.1016/j.cub.2013.02.033。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Alwan H.A.、Van Zoelen E.J.、Van Leeuwen J.E.配体诱导的溶酶体表皮生长因子受体(EGFR)降解是在蛋白酶体依赖的EGFR去泛素化之前进行的。 生物学杂志。 化学。 2003; 278:35781–35790. doi:10.1074/jbc。 M301326200。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Apaja P.M.,Lukacs G.L.质膜蛋白质稳态。 生理学。 2014; 29:265–277. doi:10.1152/physical.00058.2013。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Apaja P.M.,Xu H.,Lukacs G.L.质膜未折叠蛋白的质量控制。 《细胞生物学杂志》。 2010; 191:553–570. doi:10.1083/jcb.201006012。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Bache K.G.、Raiborg C.、Mehlum A.、Stenmark H.STAM和Hrs是早期内体上多价泛素结合复合物的亚单位。 生物学杂志。 化学。 2003; 278:12513–12521. doi:10.1074/jbc。 M210843200。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Bache K.G.、Stuffers S.、Maleröd L.、Slagsvold T.、Raiborg C.、Lechardeur d.、Wälchli S.、Lukacs G.L.、Brech A.、Stenmark H.表皮生长因子受体降解而非沉默需要ESCRT-III亚基hVps24。 分子生物学。 塞尔特。 2006; 17:2513–2523. doi:10.1091/mbc。 E05-10-0915。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Barriere H.,Lukacs G.L.通过单细胞荧光比率成像分析内吞贩运。 货币。 Protoc公司。 摄氏度。 生物学2008; 40:15.13.1–15.13.21. doi:10.1002/047114303.cb1513s40。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Barriere H.、Nemes C.、Du K.、Lukacs G.L.通过内胚体分选机制将多泛素识别为溶酶体靶向信号的可塑性。 分子生物学。 塞尔特。 2007; 18:3952–3965. doi:10.1091/mbc。 E07-07-0678。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Barriere H.、Nemes C.、Lechardeur D.、Khan-Mhammad M.、Fruh K.、Lukacs G.L.哺乳动物细胞膜蛋白寡泛素依赖性内化的分子基础。 交通。 2006; 7:282–297. doi:10.1111/j.1600-0854.2006.00384.x。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Belle L.、Ali N.、Lonic A.、Li X.、Paltridge J.L.、Roslan S.、Herrmann D.、Conway J.R.、Gehling F.K.、Bert A.G.酪氨酸磷酸酶PTPN14(Pez)通过改变蛋白质运输来抑制转移。 科学。 信号。 2015; 8:ra18。 doi:10.1126/scisional.2005547。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Budzinska M.I.、Villarroel-Campos D.、Golding M.、Weston A.、Collinson L.、Snijders A.P.、Schiavo G.PTPN23结合动力蛋白适配器BICD1,是神经营养素受体内吞分选所必需的。 细胞科学杂志。 2020; 133个doi:10.1242/jcs.242412。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Chen Y.-G.,Wang Z.,Ma J.,Zhang L.,Lu Z.Endofin,FYVE域蛋白,与Smad4相互作用并促进转化生长因子-β信号传导。 生物学杂志。 化学。 2007; 282:9688–9695. doi:10.1074/jbc。 M611704200。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Chen Y.、Low T.Y.、Choong L.Y.、Ray R.S.、Tan Y.L.、Toy W.、Lin Q.、Ang B.K.、Wong C.H.、Lim S.磷蛋白组学确定Endofin、DCBLD2和KIAA0582是人类癌细胞中EGF信号和Iressa的新型酪氨酸磷酸化靶点。 蛋白质组学。 2007; 7:2384–2397. doi:10.1002/pmic.200600968。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Christ L.、Raiborg C.、Wenzel E.M.、Campsteijn C.、Stenmark H.ESCRT膜断裂机制的细胞功能和分子机制。 生物化学科学趋势。 2017; 42:42–56. doi:10.1016/j.tibs.2016.08.016。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Desrochers G.,Kazan J.M.,Pause A.受体贩运和癌症中His域蛋白酪氨酸磷酸酶的结构和功能(1)生物化学。 细胞生物学。 2019; 97:68–72. doi:10.1139/bcb-2017-0322。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Doyotte A.、Mironov A.、Mckenzie E.、Woodman P.Bro1-相关蛋白HD-PTP/PTPN23是内胚体货物分类和多泡体形态发生所必需的。 程序。 国家。 阿卡德。 科学。 2008; 105:6308–6313. doi:10.1073/pnas.0707601105。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Eden E.R.,Huang F.,Sorkin A.,Futter C.E.EGF受体泛素化在调节其细胞内流量中的作用。 交通。 2012; 13:329–337. doi:10.1111/j.1600-0854.2011.01305.x。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Elias J.E.,Gygi S.P.通过质谱法提高大规模蛋白质鉴定可信度的目标诱饵搜索策略。 自然方法。 2007; 4:207–214. doi:10.1038/nmeth1019。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Gahloth D.、Heaven G.、Jowitt T.A.、Mould A.P.、Bella J.、Baldock C.、Woodman P.、Tabernero L.HD-PTP磷酸酶的开放结构为ESCRT功能的调节机制提供了新的见解。 2017年科学代表; 7:9151. doi:10.1038/s41598-017-09467-9。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Gahloth D.、Levy C.、Heaven G.、Stefani F.、Wunderley L.、Mould P.、Cliff M.J.、Bella J.、Fielding A.J.、Woodman P.ESCRT调节器HD-PTP和UBAP1之间选择性相互作用的结构基础。 结构。 2016; 24:2115–2126. doi:10.1016/j.str.2016.10.006。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Gahloth D.、Levy C.、Walker L.、Wunderley L.、Mould A.P.、Taylor S.、Woodman P.和Tabernero L.TGFβ信号调节剂SARA/Endofin与HD-PTP特定相互作用的结构基础。 结构。 2017; 25网址:10.1016/j.str.2017.05.005。 1011–1024.e4。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Gahloth D.、Levy C.、Walker L.、Wunderley L.、Mould A.P.、Taylor S.、Woodman P.和Tabernero L.TGFβ信号调节剂SARA/Endofin与HD-PTP特定相互作用的结构基础。 结构。 2017; 2011年10月25日至1024.e4。 doi:10.1016/j.str.2017.05.005。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Gingras M.-C.、Zhang Y.L.、Kharitidi D.、Barr A.J.、Knapp S.、Tremblay M.L.、Pause A.HD-PTP是一种催化活性酪氨酸磷酸酶,因为其磷酸酶结构域存在保守的差异。 《公共科学图书馆·综合》。 2009; 4:e5105。 doi:10.1371/journal.pone.0005105。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Goh J.B.、Wallace D.F.、Hong W.、Subramaniam V.N.Endofin,铁调节激素hepcidin的新型BMP-SMAD调节剂。 2015年科学报告; 5个doi:10.1038/srep13986。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Hesketh G.G.、Papazotos F.、Pawling J.、Rajendran D.、Knight J.D.、Martinez S.、Taipale M.、Schramek D.、Dennis J.W.、Gingras A.-C.GATOR–Rag GTPase途径通过溶酶体衍生氨基酸抑制mTORC1激活。 科学。 2020; 370:351–356. doi:10.1126/science.aaz0863。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
侯杰,杜艳Y.L.,黄鹏,崔海。整合素α5β1在人类癌症中的作用。 肿瘤靶点治疗。 2020; 13:13329. doi:10.2147/OTT。 S273803。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Huang F.、Kirkpatrick D.、Jiang X.、Gygi S.、Sorkin A.通过激酶域内的多泛素化对EGF受体内化和降解的差异调节。 摩尔摄氏度。 2006; 21:737–748. doi:10.1016/j.molcel.2006.02.018。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Ichioka F.、Takaya E.、Suzuki H.、Kajigaya S.、Buchman V.L.、Shibata H.、Maki M.HD-PTP和Alix共享一些与其Bro1域或富含脯氨酸区域相互作用的膜交通相关蛋白。 架构(architecture)。 生物化学。 生物物理学。 2007; 457:142–149. doi:10.1016/j.abb.2006.11.008。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Iyengar P.V.对TGF-β途径中泛素酶的调节。 国际分子科学杂志。 2017; 18:877. doi:10.3390/ijms18040877。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Katoh Y.、Shiba Y.、Mitsuhashi H.、Yanagida Y.、Takatsu H.、Nakayama K.Tollip和Tom1形成复合物,将泛素结合蛋白招募到早期内体上。 生物学杂志。 化学。 2004; 279:24435–24443. doi:10.1074/jbc。 M400059200。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Kazan J.M.、Lukacs G.L.、Apaja P.M.、Pause A.Springer; 2019.用于研究受体贩运中Escrt功能的单细胞荧光比率图像分析。 ESCRT复合体。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Kharitidi D.、Apaja P.M.、Manteghi S.、Suzuki K.、Malitskaya E.、Roldan A.、Gingras M.-C.、Takagi J.、Lukacs G.L.、Pause A.整合素α5β1泛素化、内吞分选和细胞迁移中内体pH值和配体占有率的相互作用。 细胞报告2015; 13:599–609. doi:10.1016/j.celrep.2015.09.024。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Kim S.、Mischerikow N.、Bandeira N.、Navarro J.D.、Wich L.、Mohammed S.、Heck A.J.、Pevzner P.A.串联质谱对CID、ETD和CID/ETD的生成函数:数据库搜索应用。 分子细胞蛋白质组学。 2010; 9:2840–2852. doi:10.1074/mcp。 M10.003731。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Knight J.D.、Choi H.、Gupta G.D.、Pelletier L.、Raught B.、Nesvizhskii A.I.、Gingras A.-C.ProHits-viz:一套可视化交互蛋白质组数据的网络工具。 自然方法。 2017; 14:645–646. doi:10.1038/nmeth.4330。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Lahaie S.、Morales D.、Bagci H.、Hamoud N.、Castonguay C.E.、Kazan J.M.、Desrochers G.、Klar A.、Gingras A.C.、Pause A.。细胞塌陷和脊髓运动轴突引导中ephrin-B:EphB信号需要内体分选适配器HD-PTP。 科学。 2019年报告; 9:11945. doi:10.1038/s41598-019-48421-9。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Lambert J.-P.、Tucholska M.、Go C.、Knight J.D.、Gingras A.-C.接近生物素化和亲和纯化是染色质相关蛋白复合物相互作用组定位的补充方法。 蛋白质组学杂志。 2015; 118:81–94. doi:10.1016/j.jprot.2014.09.011。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Lee J.,Oh K.-J.,Lee D.,Kim B.Y.,Choi J.S.,Ku B.,Kim S.J.在含有STAM2(ESCRT-0的一个亚单位)核心区域的复合体中的HD-PTP Bro1结构域的结构研究。 请给我一个。 2016; 11:e0149113。 doi:10.1371/journal.pone.0149113。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Liu G.、Knight J.D.、Zhang J.P.、Tsou C.-C.、Wang J.、Lambert J.-P.、Larsen B.、Tyers M.、Raught B.、Bandeira N.。ProHits 4.0中的数据独立采集分析。 蛋白质组学杂志。 2016; 149:64–68. doi:10.1016/j.jprot.2016.04.042。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Lobert V.H.、Brech A.、Pedersen N.M.、Wesche J.、Oppelt A.、Maleröd L.、Stenmark H.整合素的泛素化通过纤维连接蛋白-整合素复合物的溶酶体降解控制成纤维细胞的迁移。 开发委员会。 2010; 19:148–159. doi:10.1016/j.devcel.2010.06.010。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Lobert V.H.,Stenmark H.ESCRT机制通过调节肌球蛋白轻链调节成纤维细胞的极化。 J.凯尔。 科学。 2012; 125:29–36. doi:10.1242/jcs.088310。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Lu Q.,Hope L.W.,Brasch M.,Reinhard C.,Cohen S.N.TSG101与HRS的相互作用介导了内体转运和受体下调。 程序。 国家。 阿卡德。 科学。 2003; 100:7626–7631. doi:10.1073/pnas.09329599100。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Ma H.、Wardega P.、Mazaud D.、Klosowska-Wardega A.、Jurek A.、Engstrom U.、Lennartsson J.、Heldin C.H.含组氨酸域蛋白酪氨酸磷酸酶调节血小板衍生生长因子受体的细胞内分选和降解。 细胞信号。 2015; 27:2209–2219. doi:10.1016/j.cellsig.2015.07.020。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Mamiñska A.、Bartosik A.、Banach-Orłowska M.、Pilecka I.、Jastrzębski K.、Zdżalik-Bielecka D.、Castanon I.、Poulain M.、Neyen C.、Woliñska-Nizio L.ESCRT蛋白通过贩运细胞因子受体限制构成性NF-κB信号传导。 科学。 信号。 2016; 9:ra8。 doi:10.1126/scisignal.aad0848。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Manteghi S.、Gingras M.-C.、Kharitidi D.、Galarneau L.、Marques M.、Yan M.、Cencic R.、Robert F.、Paquet M.、Witcher M.ESCRT成分HD-PTP的单倍体不足易患癌症。 细胞报告2016; 15:1893–1900. doi:10.1016/j.celrep.2016.04.076。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Mierke C.T.、Frey B.、Fellner M.、Herrmann M.、Fabry B.整合素α5β1通过增强收缩力促进癌细胞侵袭。 J.凯尔。 科学。 2011; 124:369–383. doi:10.1242/jcs.071985。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Miller D.S.J.、Bloxham R.D.、Jiang M.、Gori I.、Saunders R.E.、Das D.、Chakravarty P.、Howell M.、Hill C.S.TGF-β信号的动力学由通过ESCRT机制的受体贩运决定。 Cell Rep.2018; 25:1841–1855.e5。 doi:10.1016/j.celrep.2018.10.056。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Norris A.、Tammineni P.、Wang S.、Gerdes J.、Murr A.、Kwan K.Y.、Cai Q.、Grant B.D.SNX-1和RME-8反对在内体上组装HGRS-1/ESCRT-0降解微结构域。 程序。 国家。 阿卡德。 科学。 2017; 114:E307–E316。 doi:10.1073/pnas.1612730114。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Okiyoneda T.、Barrière H.、Bagdány M.、Rabeh W.M.、Du K.、Höhfeld J.、Young J.C.、Lukacs G.L.外周蛋白质量控制从质膜上去除未折叠CFTR。 科学。 2010; 329:805–810. doi:10.1126/science.1191542。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Pandey A.、Fernandez M.M.、Steen H.、Blagoev B.、Nielsen M.M.,Roche S.、Mann M.、Lodish H.F.通过质谱鉴定新型免疫受体酪氨酸基活化基序包含分子STAM2及其参与生长因子和细胞因子受体信号通路。 生物学杂志。 化学。 2000; 275:38633–38639. doi:10.1074/jbc。 M007849200。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Parkinson M.D.、Piper S.C.、Bright N.A.、Evans J.L.、Boname J.M.、Bowers K.、Lehner P.J.、LUZIO J.P.。非标准ESCRT途径,包括组氨酸域磷酸酪氨酸磷酸酶(HD-PTP),用于下调病毒泛素化MHCⅠ类生物化学。 2015年7月; 471:79–88. doi:10.1042/BJ20150036。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Pashkova N.、Gakhar L.、Winistorfer S.C.、Sunshine A.B.、Rich M.、Dunham M.J.、Yu L.、Piper R.C.酵母Alix同源物Bro1作为泛素受体发挥作用,将蛋白质分选成多泡内涵体。 开发Cel。 2013; 25:520–533. doi:10.1016/j.devcel.2013.04.007。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Piper R.C.,Dikic I.,Lukacs G.L.内吞作用中泛素依赖性排序。 《冷泉港展望》。 生物学2014; 6:a016808。 doi:10.1101/cshperspect.a016808。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Platta H.W.、Stenmark H.细胞内吞和信号传导。 货币。 操作。 塞尔特。 生物杂志2011; 23:393–403. doi:10.1016/j.ceb.2011.03.008。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Raiborg C.、Bache K.G.、Gilloly D.J.、Madshus I.H.、Stang E.、Stenmark H.Hrs将泛素化蛋白质分类为早期内体的网格蛋白涂层微域。 国家委员会。 生物杂志,2002年; 4:394–398. doi:10.1038/ncb791。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Raiborg C.、Bache K.G.、Mehlum A.、Stang E.、Stenmark H.Hrs将氯菊酯招募到早期内体中。 EMBO J.2001; 20:5008–5021. doi:10.1093/emboj/20.17.5008。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Raiborg C.、Wesche J.、Malerod L.、Stenmark H.内体上的扁平网格蛋白涂层通过动态微域中的支架化Hrs介导降解蛋白质的分选。 细胞科学杂志。 2006; 119:2414–2424. doi:10.1242/jcs.02978。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Rodahl L.M.、Stuffers S.、Lobert V.H.、Stenmark H.ESCRT蛋白在细胞信号衰减中的作用。 生物化学。 社会事务处理。 2009; 37:137–142. doi:10.1042/BST0370137。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Rosenberger G.、Koh C.C.、Guo T.、Röst H.L.、Kouvonen P.、Collins B.C.、Heusel M.、Liu Y.、Caron E.、Vichalkovski A.通过SWATH-MS科学数据量化10000人类蛋白质的分析库。 2014; 1:1–15. doi:10.1038/数据.2014.31。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Row P.E.、Clague M.J.和UrbéS.生长因子诱导Hrs–STAM复合物的不同磷酸化特征:具有信号特异性的信号网络中的一个常见节点。 生物化学。 J.2005; 389:629–636. doi:10.1042/BJ20050067。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Sachse M.、UrbéS.、Oorschot V.、Strous G.J.、Klumperman J.内体液泡上的双层网格蛋白涂层参与蛋白质向溶酶体的分选。 分子生物学。 塞尔特。 2002; 13:1313–1328. doi:10.1091/mbc.01-10-0525。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Seet L.F.,Hong W.Endofin,一种内体FYVE结构域蛋白。 生物学杂志。 化学。 2001; 276:42445–42454. doi:10.1074/jbc。 M105917200。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
见L.-F.,Hong W.Endofin通过TOM1向早期内体招募网格蛋白。 J.凯尔。 科学。 2005; 118:575–587. doi:10.1242/jcs.01628。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
见L.-F.、Liu N.、Hanson B.J.、Hong W.Endofin将TOM1招募到内体。 生物学杂志。 化学。 2004; 279:4670–4679. doi:10.1074/jbc。 M311228200。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Shi W.,Chang C.,Nie S.,Xie S.,Wan M.,Cao X.Endofin在BMP信号传导中充当Smad受体激活锚。 J.凯尔。 科学。 2007; 120:1216–1224. doi:10.1242/jcs.03400。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Sigismund S.、Avanzato D.和Lanzetti L.EGFR在癌症中的新功能。 摩尔·昂科尔。 2018; 12:3–20. doi:10.1002/1878-0261.12155。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Sorkin A.、von Zastrow M.《细胞内吞和信号传递:相互缠绕的分子网络》。 自然修订版摩尔Cel。 《生物》2009; 10:609–622. doi:10.1038/nrm2748。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
St-Denis N.、Gupta G.D.、Lin Z.Y.、Gonzalez-Badillo B.、Veri A.O.、Knight J.D.、Rajendran D.、Couzens A.L.、Currie K.W.、Tkach J.M.人类磷酸酶的表型和相互作用分析确定了不同的有丝分裂调节器。 细胞报告2016; 17:2488–2501. doi:10.1016/j.celrep.2016.10.078。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Stefani F.、Zhang L.、Taylor S.、Donovan J.、Rollinson S.、Domyotte A.、Brownhill K.、Bennion J.、Pickering-Brown S.、Woodman P.UBAP1是内胚体特异性ESCRT-I复合物的组成部分,对MVB分选至关重要。 货币。 生物杂志2011; 21:1245–1250. doi:10.1016/j.cub.2011.06.028。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Sundquist W.I.、Schubert H.L.、Kelly B.N.、Hill G.C.、Holton J.M.和Hill C.P.人类TSG101蛋白对泛素的识别。 摩尔摄氏度。 2004; 13:783–789. doi:10.1016/s1097-2765(04)00129-7。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Szklarczyk D.、Gable A.L.、Lyon D.、Junge A.、Wyder S.、Huerta-Cepas J.、Simonovic M.、Doncheva N.T.、MORRIS J.H.、BORK P.STRING v11:覆盖率增加的蛋白质-蛋白质关联网络,支持全基因组实验数据集中的功能发现。 2019年《核酸研究》; 47:D607–D613。 doi:10.1093/nar/gky1131。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Szymanska E.,Budick Harmelin N.,Miaczynska M.内源性信号控制:ESCRT机制在受体介导的信号通路调节中的作用。 塞米恩。 细胞发育生物学。 2018; 74:11–20. doi:10.1016/j.semcdb2017.08.012。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Teo G.、Liu G.、Zhang J.、Nesvizhskii A.I.、Gingras A.-C.、Choi H.SAINTexpress:INTeractome软件显著性分析的改进和附加功能。 蛋白质组学杂志。 2014; 100:37–43. doi:10.1016/j.jprot.2013.10.023。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Toyoshima M.、Tanaka N.、Aoki J.、Taanaka Y.、Murata K.、Kyuuma M.、Kobayashi H.、Ishii N.、Yaegashi N.和Sugamura K.通过耗尽囊泡分选蛋白Hrs抑制肿瘤生长和转移:其对E-cadherin和beta-catenin的调节作用。 癌症研究2007; 67:5162–5171. doi:10.1158/0008-5472.CAN-06-2756。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
UrbéS.、Mills I.G.、Stenmark H.、Kitamura N.、Clague M.J.内体定位和受体动力学确定肝细胞生长因子调节酪氨酸激酶底物的酪氨酸磷酸化。 分子细胞生物学。 2000; 20:7685–7692. doi:10.1128/mcb.20.20.7685-7692.2000。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Vietri M.、Radulovic M.、Stenmark H.ESCRT的多种功能。 自然修订版分子细胞生物学。 2020; 21:25–42. doi:10.1038/s41580-019-0177-4。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Wang J.、Tucholska M.、Knight J.D.、Lambert J.-P.、Tate S.、Larsen B.、Gingras A.-C.、Bandeira N.MSPLIT-DIA:数据依赖性采集的敏感肽鉴定。 自然方法。 2015; 12:1106–1108. doi:10.1038/nmeth.3655。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Wang T.,Liu N.S.,Seet L.F.,Hong W.VHS域包含Tom1、Tom1L1和Tom1L2在膜贩运中的新兴作用。 交通。 2010; 11:1119–1128. doi:10.1111/j.1600-0854.2010.01098.x。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Wegener C.S.、Maleröd L.、Pedersen N.M.、Prodiga C.、Bakke O.、Stenmark H.、Brech A.由GTPase缺陷的Rab5诱导的巨大内体的超微结构特征。 组织化学。 塞尔特。 生物.2010; 133:41. doi:10.1007/s00418-009-0643-8。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Wegner C.S.、Rodahl L.M.、Stenmark H.ESCRT蛋白质和细胞信号。 交通。 2011; 12:1291–1297. doi:10.1111/j.1600-0854.2011.01210.x。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Wenzel E.M.、Schultz S.W.、Schink K.O.、Pedersen N.M.、Nähse V.、Carlson A.、Brech A.、Stenmark H.、Raiborg C.协调的ESCRT和网格蛋白募集波定义了腔内小泡形成的时间和形态。 国家公社。 2018; 9:1–18. doi:10.1038/s41467-018-05345-8。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Yamaguchi H.,Wyckoff J.,Condeelis J.肿瘤中的细胞迁移。 货币。 操作。 塞尔特。 生物学2005; 17:559–564. doi:10.1016/j.ceb.2005.08.002。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
Yamakami M.,Yoshimori T.,Yokosawa H.Tom1,一种含有VHS结构域的蛋白质,与tollip、泛素和网格蛋白相互作用。生物学杂志。 化学。 2003; 278:52865–52872. doi:10.1074/jbc。 M306740200。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
关联数据 本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。