越来越多的证据表明α-突触核蛋白与帕金森病(PD)的发病机制有关(Polymeropoulos等人,1997年;Kruger等人,1998年;Spialantini等人,1998年;Takeda等人,1998年;Giasson等人,2000b). 发现α-突触核蛋白基因的点突变在罕见的家族性PD病例中可能致病(Polymeropoulos等人,1997年;Kruger等人,1998年)导致确定α-突触核蛋白是路易小体和营养不良性神经节的主要成分,即PD的病理学霍尔标志(Spialantini等人,1997年,1998;Baba等人,1998年;Irizarry等人,1998年;武田等人,1998年;Braak和Braak,2000年). 后续在体外研究表明,α-突触核蛋白可能与泛素蛋白体加工、氧化损伤和/或线粒体功能障碍相互作用,导致神经变性和细胞死亡(Ostrerova等人,1999年;Giasson等人,2000b;Hsu等人,2000年;神田等人,2000年;Tabrizi等人,2000年;McNaught等人,2001年;Shimura等人,2001年). 为了支持这一概念,最近体内研究表明,野生型(wt)或突变的α-突触核蛋白在转基因小鼠中的过度表达可能导致神经病理学改变、轴突变性,偶尔也可能导致细胞死亡(Kahle等人,2000年;Masliah等人,2000年;van der Putten等人,2000年).
然而,在迄今为止产生的转基因小鼠中,α-突触核蛋白的积累和轴突病理学的征象广泛存在,并且在黑质纹状体系统以外的结构中最为突出。尽管Masliah等人(2000年)观察到黑质(SN)神经元中α-突触核蛋白的积累和纹状体中酪氨酸羟化酶(TH)阳性轴突终末的减少(仅在转基因表达最高的小鼠中获得),在这些小鼠中均未观察到黑纹状体多巴胺(DA)神经元的变性或丢失。事实上,随后对表达A30P的转基因小鼠的研究(Matsuoka等人,2001年;Rathke-Hartlieb等人,2001年)或A53T(Matsuoka等人,2001年)突变形式的α-突触核蛋白没有发现任何黑质神经病理学。因此,目前尚不清楚α-突触核蛋白的突变形式在啮齿类动物的黑质纹状体DA神经元中起到什么作用,或者α-突触核蛋白的过度表达在多大程度上足以引起帕金森样病理学和神经变性。其他罕见的家族性帕金森病已被描述为与蛋白体加工途径有关的帕金森病病因。因此,除了α-突触核蛋白的突变外,一种E3泛素连接酶parkin的错义突变(Shimura等人,2000年)和泛素C末端羟化酶-L1,一种E1泛素激活酶(Leroy等人,1998年)都会诱发家族性PD。事实上,22kDaO(运行)-α-突触核蛋白的糖基化亚型被发现是E3泛素连接酶parkin的特异性底物(Shimura等人,2001年). 突变形式的α-突触核蛋白(A53T和A30P)不能与帕金结合,从而取消了其正常的多泛素化和随后的蛋白质组加工。因此,这些数据表明,α-突触核蛋白的积累,如特发性帕金森病患者路易小体中发生的,可能是通过抑制正常蛋白体处理或过度表达,导致黑质多巴胺神经元的致病。
在本研究中,我们使用重组腺相关病毒(rAAV)载体在成年大鼠黑质纹状体DA神经元中高水平表达wt和突变的α-突触核蛋白。rAAV载体整合并在非分裂细胞(主要是中枢神经系统的神经元)中稳定表达其转基因。由于96%的病毒基因组已被清除,载体不表达任何病毒蛋白,从而将宿主免疫反应的风险降至最低(Muzyczka,1992年). 该载体系统对致密部SN神经元具有特殊的亲和力,使成年大鼠黑质纹状体DA神经元中的蛋白质能够稳定、高水平表达(Klein等人,1998年;Kirik等人,2000年). 此外,转导可以限制在一个半球,这样对侧可以作为对照。
材料和方法
动物和手术。年轻成年Sprague-Dawley大鼠(B&K Universal AB,斯德哥尔摩,瑞典)被关在笼子里,六只随意根据隆德大学研究伦理委员会制定的规则,在12小时的光/暗周期内获取食物和水。在氟烷麻醉下,将2μl载体悬浮液立体定向注射在右SN上,位于前角后方5.2 mm、外侧2.0 mm、硬脑膜腹侧7.2 mm处。在缓慢拔出之前,将针再放置5分钟。rAAV-CBA-α-突触核蛋白(8.2×1011IU/ml),rAAV-CBA-μ-α-突触核蛋白(1.4×1012IU/ml)和rAAV-CBA-GFP(1.4×1011IU/ml),并按所述进行滴定(McLaughlin等人,1988年;Conway等人,1997年;Zolotukhin等人,1999年). 如前所述,这里使用的CBA启动子是一个鸡β-肌动蛋白启动子,带有来自巨细胞病毒启动子的增强子元件(Xu等人,2001年). 为了进行组织学分析,在注射[绿色荧光蛋白(GFP)组,n个= 5; α-突触核蛋白基团,n个= 5–6; 和突变的α-突触核蛋白组,n个所有时间点=5,注射后3、8和27周进行生化分析(n个=3周时为4–5,以及n个=8周和27周时每组5人)。第二组动物(n个18例)双侧纹状体注射0.2μl 2%氟金(FG)溶液(前侧1.0 mm、外侧3.0 mm和腹侧5.0 mm),5天后进行单侧载体注射,如上所述(n个=每组6人)。这些动物在注射病毒后3周和8周被杀死(n个在每个时间点=9)。第三组动物(n个=7)在右侧单侧黑质注射rAAV-CBA-α-突触核蛋白之前,在SN中双侧注射rAAV-CBA-GFP载体。这些动物在3点被杀(n个=3)和8周(n个=4)注射后。
一组单独的动物(n个=30)注射rAAV-CBA-GFP(n个=10),rAAV-CBA-α-突触核蛋白(n个=10),或rAAV-CBA突变型μ-α-synuclein(n个=10)载体,用于α-甲基治疗-d、 我-对位-酪氨酸甲酯如下所述。
免疫组织化学。在戊巴比妥麻醉下,动物通过升主动脉灌注生理盐水,然后灌注4%冰镇多聚甲醛。将大脑在相同溶液中固定2小时,转移到25%蔗糖中,并在冠状面40μm的冷冻切片机上切片。使用针对TH(小鼠IgG,1:2000;加州特梅库拉Chemicon)、GFP(鸡IgG 1:5000;明尼苏达州明尼阿波利斯R&D systems)、囊泡单胺转运蛋白-2(VMAT-2)(兔IgG:1:5000;Chemicon。Goldman,康奈尔大学)和人类α-突触核蛋白(小鼠IgG,1:2000;由宾夕法尼亚大学弗吉尼亚·M·李博士提供)。在每个培养期之间,将切片在磷酸钾缓冲液(KPBS)中冲洗三次。所有培养液均含有0.25%Triton X-100 in KPBS。在3%H中淬火10 min2O(运行)2/10%甲醇。用5%的正常马血清(NHS)预孵育2小时后,在室温下用一级抗体在2%NHS中孵育,用1:200稀释的生物素化马抗鼠抗体(BA2001;Vector Laboratories,Burlingame,CA)在2%NHS中孵养,然后用亲和素-生物素-过氧化物酶复合物(ABC Elite;Vector Laboratories,Burlingame,CA),并使用3,3-二氨基联苯胺作为显色剂进行可视化,安装在镀铬-明矾玻璃载玻片和盖玻片上。对于α-突触核蛋白和TH的双重荧光免疫组织化学分析,使用Alexa 488和Cy3-共轭二级抗体。
根据光学分馏器原理,使用OlympusDenmark A/S(Albertslund,Denmarks)CAST-Grid系统评估SN中TH阳性神经元的总数,如下所述(Kirik等人,1998年). 计数程序中包括涵盖序列号整个范围的每八个部分。误差系数小于0.10是可以接受的。纹状体TH阳性纤维密度通过密度测定法在三个冠状水平(+1.0、0.0、-1.0)下测量(相对于前角)。数据表示为三个水平的平均值,并表示为完整控制侧的百分比。
生化分析。组织DA和DOPAC含量(Schmidt等人,1982年)和在体外TH酶活性(Reinhard等人,1986年)在来自纹状体和SN(在所有时间点)以及来自27周组动物的伏隔核和前额叶皮层的组织样本上测定。称量组织块并将其分别冷冻在干冰上,并在−80°C下保存,直到进行分析。
行为测试。在载体注射后8周、13周、19周和27周以及皮下注射0.25 mg/kg脱吗啡HCl(西格玛,圣路易斯,密苏里州)后评估药物诱导的旋转,并在自动旋转仪中监测40分钟。数据表示为每分钟净满圈数,对侧圈数为负值。
如前所述,在载体注射后24-25周进行爪伸楼梯试验(Kirik等人,1998年). 剥夺食物2天后,将动物放在试验箱中,在四个步骤中的每一步(45 mg)用10粒食物颗粒进行双边诱饵。使用标准平台(27 mm)连续9天进行15分钟以上的测试,使用更宽的平台(34 mm)再进行5天。在宽平台测试的最后3天内,成功回收球团的平均总数构成因变量。
按所述进行步进测试(Kirik等人,1998年)注射病毒载体后8周。动物(n个=每组10人),连续3天每天测试两次,以确定基线表现。第四天,在腹腔注射60 mg/kgα-甲基之前,对动物进行一次测试-数字图书馆-对位-酪氨酸甲酯(溶于0.02%抗坏血酸盐;Sigma);注射后6、24和48小时再次注射。当大鼠沿着桌子表面正手方向侧向移动时(5秒内90厘米),计算调整步数。
统计分析。进行了双向重复测量方差分析临时的使用简单的主效应进行分析,并对可接受的α水平进行Bonferroni校正对= 0.05. 为了比较各组之间的黑质细胞数量和纹状体纤维密度测量值以及存活时间,采用双向因子方差分析和临时的使用简单的主效应进行分析,并对可接受的α水平进行Bonferroni校正对= 0.05.
结果
为了研究α-突触核蛋白过表达对黑色素的影响,大鼠接受了一次2μl的高滴度rAAV载体注射,该载体在SN致密部(SNc)单侧含有人类α-突触素基因的wt或A53T突变。对照组大鼠接受了编码GFP的rAAV载体的相同注射。免疫组织化学,使用识别人类(wt和突变)而非大鼠α-突触核蛋白的抗体(Giasson等人,2000a)α-突触核蛋白在SNc内的几乎所有神经元、SN网状部(SNr)内的大量神经元以及SN背侧中脑网状结构内的神经元中均有表达(图。2E–L公司). 腹侧被盖区(VTA)中数量不等的细胞也呈阳性。在注射rAAV-GFP的对照动物中也观察到类似的细胞表达模式(图。1). 双TH/α-突触核蛋白免疫荧光显示,SNc中90%以上的TH阳性神经元表达α-突触核蛋白转基因(图。2M–O型).
图2。
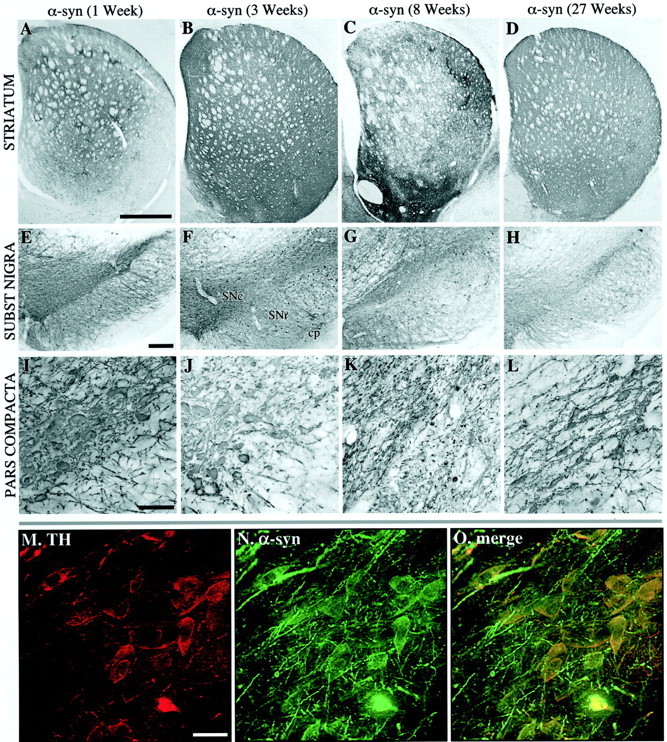
纹状体α-突触核蛋白表达综述(A–D)和序号(E–O公司)在1(A、 E、I),第3页(B、 F、J、M–O), 8 (C、 G,K)、和27(D、 H、I)注射后数周。黑质DA细胞在1(E、 我)和3(F、 J型)转导后数周。3周时,致密部中大多数TH阳性细胞体的α-突触核蛋白也呈阳性(M–O型). 同样,纹状体中的黑质纹状体纤维终末充满转基因人α-突触核蛋白(比较A、 B类). 与rAAV-GFP注射动物相比,α-synuclein的表达导致了黑质细胞体的退行性改变(G、 K(K))纹状体末端(C). 致密部细胞体的退化被视为8周时致密部SN中α-突触核蛋白免疫反应性细胞体强度的降低(K(K)). 在27周时,α-同核阳性细胞体的减少仍然很低(H、 L(左))而纹状体水平有一定程度的恢复(天). 比例尺:A类,1 mm(适用于A–C);E类,250μm(适用于E–H(E–H));我,200μm(适用于I–L型);M(M),30μm(适用于M–O型).
图1。
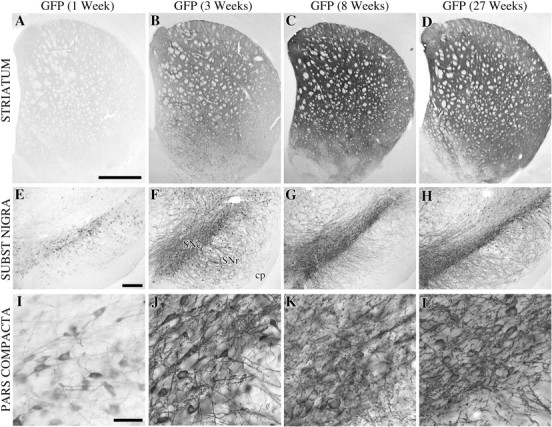
纹状体中GFP表达程度概述(A–D)和序号(E–L公司)在1(A类,E类,我), 3 (B,F类,J型), 8 (C,G公司,K(K))、和27(天,H(H),L(左))注射后数周。在SN中1周时检测到低水平的GFP蛋白(B、 C类),终端中没有或非常低的电平(A类). 3周时,细胞体的免疫反应增强,覆盖整个致密部及其网状部的树突树(F、 J)当光纤终端变得可见时(B). 8周时,纹状体终末的表达都很高(C)还有黑色素瘤,没有任何病理(G、 K(K)). 在本实验的最后一个终点,即转导27周后,GFP免疫反应与8周时获得的染色无法区分,这表明转基因表达在这两个时间点之间保持较高水平。比例尺:A类,1 mm(适用于A–C);E类,250μm(适用于E–H);我,200μm(适用于I–L).
两种类型的α-突触核蛋白(wt和突变体)都能有效地在异体内运输到各自的终末区:从SNc到纹状体,从VTA到边缘和皮质前脑区,从SNr到上丘和丘脑,从网状结构到广泛的间脑区。在黑质纹状体通路中,细胞体和轴突,包括纹状体终末的一部分,在1周时表达弥漫的细胞质α-突触核蛋白免疫反应(图。2A、 E类). 3周时,整个纹状体终末网络被填满(图。2B)包括VTA投射到嗅结节、伏隔核、外侧隔和前扣带回皮质的可变部分。在注射rAAV-GFP的对照动物中,GFP的转导和转运同样有效,完全填充了SNc神经元的细胞体、树突、轴突和轴突末端(图。1). 在最初几周内,表达逐渐增加;GFP或α-synuclein蛋白的最大表达在3-8周时观察到(图。1E–G公司,2E–G公司)并且在27周龄的动物中保持在较高水平(图。1D、 高、低,2D、 高、低). 与之前的观察结果一致(参见。Kirik等人,2000年)甲酚紫和苏木精染色切片在任何时间点均未观察到任何中毒或炎症反应的迹象。
α-突触核蛋白诱导的神经病理学和细胞死亡
在载体注射后的前2个月内,神经元病理学症状逐渐发展。这包括α-同核阳性细胞质内含物和颗粒沉积物的出现(图。三A–C,箭头),肿胀的α-突触核蛋白阳性营养不良和碎裂的神经炎(图。三B、 D–F型),和致密、萎缩的神经元胞周,胞浆中有致密的外观和强烈的α-突触核蛋白免疫反应性(图。三F类). 一些较大的包涵体有一个清晰的免疫阴性核心,周围环绕着α-同核阳性晕(图。三C、 D类). 这些变化在3周时首次在SNc神经元中观察到,并在注射后8周进一步增加,类似于特发性帕金森病中的α-突触核蛋白包涵体和营养不良神经炎(Hayashida等人,1993年;Arima等人,1998年;Braak和Braak,2000年;杜达等人,2000年;Giasson等人,2000b). 然而,它们中没有一个被泛素抗体阳性染色。
图3。
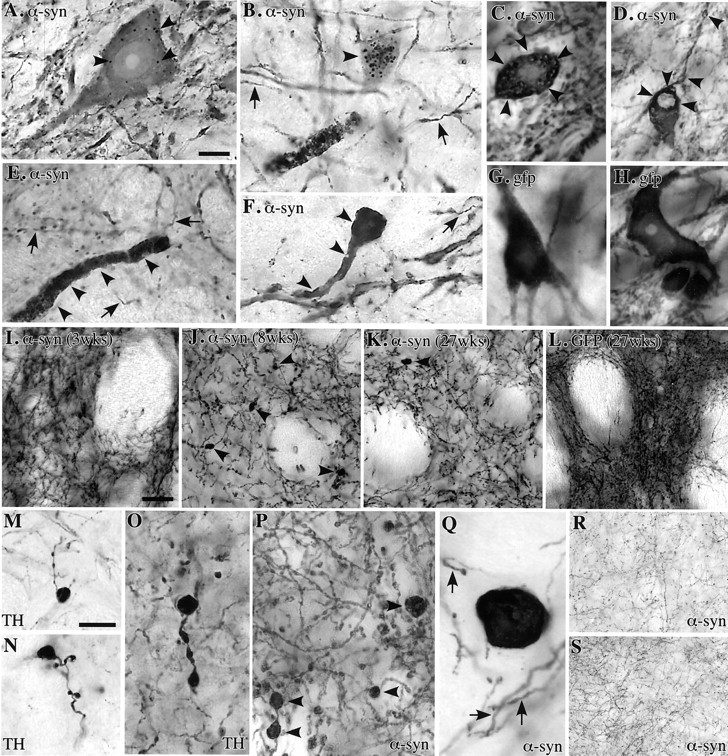
注射后8周,α-同核阳性颗粒包裹体(A、 B,箭头)以及离散的胞质内包涵体,其中α-突触核蛋白免疫反应在苍白的核周围形成晕圈(C、 D,箭头)大量出现在SNc细胞中,与肿胀的营养不良性α-同核阳性突起相关(B,E类,F类)和萎缩性外周(F类).箭头在里面B,E类,F类、和G公司显示正常大小的完整轴突;G公司,H(H)GFP过度表达的SNc神经元无上述变化;I–K型纹状体α-突触核蛋白免疫反应终末在3周时形态正常(我)但在8周时数量减少,表现出大量肿胀、营养不良的特征(J、 P,箭头). 27周时,发现营养不良纤维较少,但α-同核阳性终末的密度仍然减少(K(K));L(左)在转导GFP的对照组中未观察到任何轴突改变,尽管GFP的表达在27周前仍保持在较高水平;M–Q月TH显示纹状体中肿胀、营养不良的轴突终末(M–O型)和α-突触核蛋白抗体(P、 问);R(右),S公司α-突触核蛋白的表达不会引起上丘非多巴胺能投射的任何病理改变(R(右))和丘脑(S公司)(存活8周)。比例尺:A、,10μm(适用于A–H、O–Q);我,50μm(适用于I–L、R、S);M(M),20μm(适用于M(M),N个).
在纹状体中,注射后3周出现单个α-突触核蛋白阳性肿胀轴突,到8周时,大量α-突触核蛋白阳性营养不良轴突散布在整个纹状体中,最密集地分布在背外侧部分(图。三J、 P、Q、箭头)伴随着α-同核阳性纹状体轴突终末密度的显著降低(图。2B、 C类,三I、 J). 在嗅结节和伏隔核,偶尔在隔膜和前扣带皮层,即在VTA中接受DA神经元投射的区域,也会出现不同数量的轴突扭曲。几乎所有营养不良的神经突都对DA-合成酶TH呈阳性反应(图。三M–O型),但他们没有染色泛素或VMAT-2。
在给予wt或突变的α-突触核蛋白的动物中,这些病理变化在数量和质量上都是相似的。在表达rAAV-GFP的动物中,在任何时间点均未发现细胞或轴突病理学改变(图。1,三G、 高、低)表明病毒感染的非特异性影响或任何细胞质蛋白的高表达均不足以引起α-突触核蛋白过度表达的SNc神经元的病理变化。
为了分析α-突触核蛋白过度表达对非多巴胺能神经元的影响,我们研究了在载体注射动物中有效转导的其他三种投射系统:黑素保护区和黑素丘脑投射,以及中脑网状结构到间脑的投射。α-突触核蛋白(这两种类型)在所有三个投射系统的细胞体、轴突和终末广泛表达,但除了丘脑中单个肿胀的轴突轮廓外,这些非多巴胺能系统中明显缺乏α-突触素阳性内含物或营养不良的轴突(图。三R、 S公司).
TH免疫组织化学显示,SN中TH阳性细胞体逐渐消失,wt和突变α-突触核蛋白的情况相似,3周时平均为23%,8周时为55%(图。4A类,D–H型)(来自两个α-synuclein组的数据合并)。然而,在一些动物中,TH阳性细胞的丢失明显不同,从30-40%不等(图。4G公司)其他为70-80%(图。4H(H)). 纹状体TH阳性神经支配在转导后3周仅受到轻微影响(约为−10%)(图。5B)但在8周时整个纹状体显著减少(约50%)(图。5C、 F类). VMAT-2抗体(数据未显示)和α-突触核蛋白染色(图。2C). 对相邻切片进行TH和α-突触核蛋白染色的分析表明,剩余的部分纤维(通过其α-突触核蛋白含量确定)具有非常弱的TH免疫反应性。VTA中也出现了一些可变的TH阳性细胞丢失,同时伏隔核和嗅结节中TH阳性神经支配也出现了类似可变的减少(数据未显示)。在任何时间点,rAAV-GFP注射动物的TH阳性细胞数量和纹状体TH阳性神经支配均未受到显著影响,如TH-和GFP免疫染色切片所示(图。1C、 D类,4B、 我,5E、 F类).
图4。
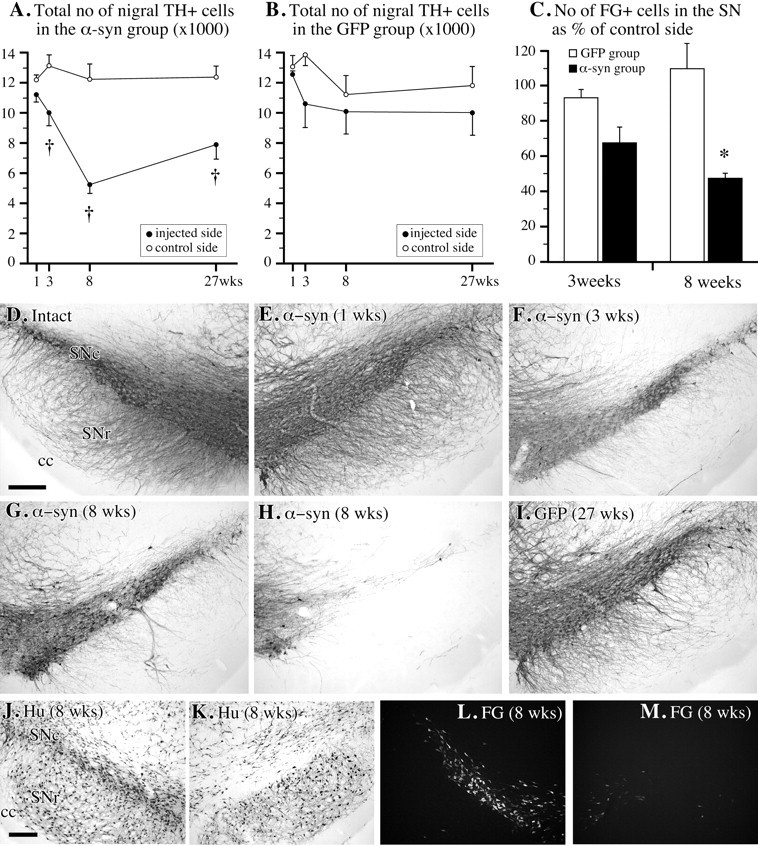
序号中的单元格丢失。A类,B,根据rAAV-α-突触核蛋白体视学测定的SNc中TH阳性细胞总数的变化(A类;n个=每个时间点10–11)和rAAV-GFP注射大鼠(B;n个=每个时间点5);C在8周时,rAAV-α-突触核蛋白转基因动物中标记FG的SNc细胞数量减少了约50%,但在GFP转基因对照组中保持不变(M(M),注入侧;L(左),对侧完好);D–H型,TH免疫组织化学显示,随着时间的推移,SNc中TH阳性细胞的逐渐丢失,变化范围约为30-40%(G公司)至70–80%(H(H));我,在rAAV-GFP处理的动物中未观察到TH阳性细胞损失;J型,K(K)注射SNc的神经元变性(K(K))通过使用神经元特异性抗体Hu进行确认。比例尺:天,J中,250μm(适用于D–I型和J–M,分别)。† 对与完好侧相比<0.0125*对与rAAV-GFP组相比,<0.05。
图5。
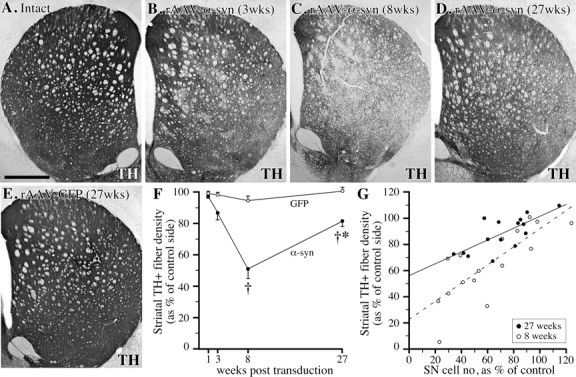
纹状体TH阳性神经支配缺失。A–ETH-免疫组化切片显示TH阳性纹状体神经支配在3(B), 8 (C)、和27周(天)黑质内注射rAAV-α-synuclein后,与正常对照侧比较(A类)rAAV-GFP注射动物的神经支配未受影响(E类).F类,密度测定显示,注射后8周和27周,α-突触核蛋白转导的动物TH阳性纹状体神经支配显著减少(†与注射后1周的值和GFP转导的对照组相比有显著差异对< 0.0125). α-synuclein组在8周至27周之间恢复显著(*对< 0.0125);G公司进一步分析黑质TH阳性细胞数与纹状体TH阳性纤维密度(对照侧的%)之间的相关性表明,通过简单的线性回归分析获得了最佳拟合。8周时,回归线(虚线)斜率为0.695年-截距22.905(对= 0.0003). 然而,在27周时,线路的坡度(固体)降至0.478,并且年-轴截距增加到55.167(对=0.0016),表明黑质纹状体投射神经元的丢失部分由剩余细胞增加的神经支配所补偿。比例尺:A类,1 mm(适用于A–E).
为了确定SN中TH阳性细胞数量的减少是否归因于细胞的实际损失或所用细胞标记物(即TH酶和VMAT蛋白)的下调,我们使用了三种独立的方法来标记黑质神经元。首先,在注射rAAV-α-synuclein载体前5天,我们使用逆行示踪剂FG双侧注射纹状体,在损伤前标记SNc中的细胞。3周时,SNc中存活的FG阳性细胞数量减少了32%,8周时减少了52%(图。4C、 L、M). wt组和突变α-synuclein组的细胞丢失相似(对> 0.05). 其次,我们使用神经元特异性蛋白Hu的抗体作为存活SNc神经元的独立标记。与FG数据一致,Hu标记显示rAAV-α-突触核蛋白注射侧的SNc细胞明显丢失,在8周时完全发育(图。4J、 K). 第三,在注射α-突触核蛋白载体之前,我们使用rAAV-GFP载体对SNc神经元进行预标记。这些动物在单侧rAAV-α-突触核蛋白注射前5天接受双侧rAAV-GFP注射。在α-突触核蛋白转导侧,SNc中GFP标记的细胞数量(注射后8周评估)显著减少(数据未显示)。
在长期存活的动物中,在载体注射6个月后进行分析,细胞体和轴突病理学的迹象大大减少(图。三K(K))这表明受损的黑质细胞体和轴突要么退化(并消失),要么从最初的损伤中恢复。然而,6个月后,存活的黑质纹状体神经元的细胞体和轴突中的α-突触核蛋白表达仍保持在高水平(图。三K(K)). 在TH免疫染色切片中也观察到恢复的迹象:纹状体中TH阳性纤维的密度增加,从8周时正常的约50%增加到6个月时正常的~80%(对<0.0125)(图。5C、 D、F). VMAT-2染色切片中也观察到类似程度的恢复(数据未显示)。进一步分析表明,纹状体TH阳性纤维的恢复,至少部分归因于纹状体轴突终末TH表达的恢复,该终末在8周时呈弱TH免疫反应。尽管SNc中TH阳性神经元的数量在8到27周之间呈现增加趋势(从正常的46%增加到65%)(图。4A类),这并没有达到意义。因此,纹状体TH阳性神经支配的恢复与SNc中TH阳性神经元的类似恢复不匹配,这表明轴突萌芽可能有助于长期存活动物纹状体TH阳性神经的恢复(图。5G公司).
黑质纹状体多巴胺系统功能下降
在纹状体和SN组织样本中,通过神经化学方法评估了α-突触核蛋白过度表达对DA合成、储存和转换的影响。纹状体DA水平(图。6A类)纹状体TH酶活性(图。6B)在3周时减少了40–50%,并且在α-synuclein转导后的8周和27周时,这些减少保持不变,wt和突变的α-synuglein的情况类似(对> 0.05). 注射rAAV-GFP的对照大鼠未发现明显变化(图。6A、 B). rAAV-α-synuclein动物在3周时SN中的DA水平降低了27.3±7.9%,8周时降低了41.8±7.0%,27周时下降了28.2±10.1%,而其他前脑区域(伏隔核和前额叶皮层)的DA水平没有变化(数据未显示)。纹状体DA和TH活性水平的降低伴随着DA转换的显著增加,以DOPAC/DA比率进行测量(图。6C). 在rAAV-GFP治疗的动物中,这种增加只是暂时的,但在注射α-突触核蛋白治疗的动物后8周和27周,这种增加维持在正常水平的~160%。伏隔核或前额叶皮层的DA转换没有变化。
图6。
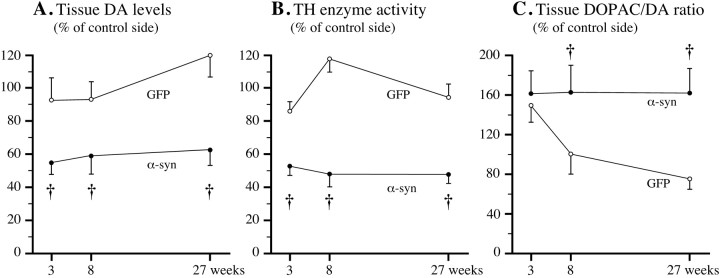
纹状体DA、DOPAC和TH活性水平随时间降低。A类,B、,纹状体DA含量(A类)和TH活性(B)在所有时间点减少了约40–50%;C根据DOPAC/DA比率评估,α-突触核蛋白转基因动物的纹状体DA转换在所有时间点都增加了~60%(实心圆圈)但GFP组仅短暂增加(开放的圆圈). † 对<0.05(组效应,双向重复测量方差分析)。
α-突触核蛋白转基因动物纹状体DA和TH活性水平的长期降低与观察到的SNc细胞丢失一致。然而,纹状体DA水平和TH活性在3周时已经最大程度降低,即在SNc中TH阳性细胞仅部分丢失(~20%)的时候,这一事实表明α-突触核蛋白(wt和突变体)细胞质水平升高可能对DA的合成和储存产生抑制作用,独立于其神经退行性作用。这种对神经递质功能的抑制作用也可以解释为什么纹状体DA和TH酶活性在6个月时仍然较低(−40至−50%),尽管此时纹状体中TH阳性纤维密度已恢复至正常值的~75%。
运动行为障碍
在一系列药物诱导和自发行为测试中监测到的运动行为,在所有时间点,α-突触核蛋白治疗的动物的运动行为总体上只受到轻微影响。这表明,观察到的DA神经元细胞丢失和纹状体DA耗竭的程度不足以诱发容易检测到的行为损伤。这与之前的研究一致,之前的研究表明,药物诱导和自发运动行为的显著损伤只会出现在黑质DA神经元和纹状体DA水平降低>50-60%的动物身上(Lee等人,1996年;Kirik等人,1998年). 然而,对个体动物的仔细检查显示,α-突触核蛋白治疗组动物的行为表现存在显著差异(图。7A、 B). 因此,与GFP对照组相比,在阿朴吗啡旋转试验和爪子疼痛试验中,约25%的α-突触核蛋白转基因动物显著受损(图。7A、 B,实心圆). 这种程度的损伤(与rAAV-GFP对照组中的30±4粒相比,足爪伸直试验中回收的10-20粒)与60-80%的黑质DA神经元的丢失相一致(Lee等人,1996年;Kirik等人,1998年).
图7。
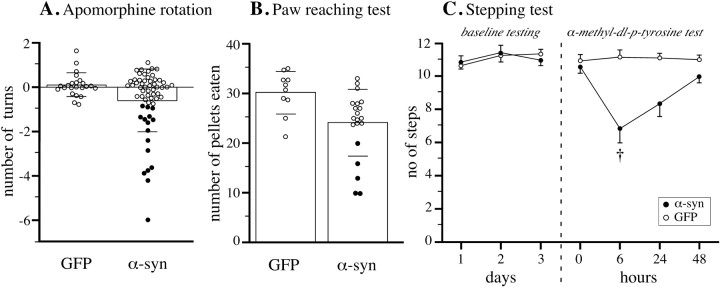
运动行为的变化。在所有三项测试中,α-突触核蛋白转基因动物的总体表现与GFP组没有差异(对>所有对比度均为0.05)。然而,在阿朴吗啡旋转试验中(A类; 来自8周试验的数据)和爪子伸直试验(B; 在24周时进行)25%的α-突触核蛋白转基因动物明显受损,因为它们的得分低于GFP对照组中的任何动物(实心圆圈). 条形图英寸A类和B给出平均值±1 SD。C,单次低剂量DA合成抑制剂α-甲基-数字图书馆-对-在测试的第四天给予酪氨酸,在步进测试中产生了明显的损伤。†与药物前试验和GFP组相比对< 0.0125.
这些数据表明,在接受α-突触核蛋白治疗的大鼠中,TH阳性黑质细胞丢失超过了~60%的临界阈值,从而导致行为障碍。众所周知,黑质纹状体DA投射的部分损害可能被残余传入中DA转换的增加部分补偿(Zigmond等人,1990年)这解释了为什么在纹状体DA神经支配丧失50-60%的动物中可以维持正常的纹状体功能。然而,这种代偿性动物对DA合成的阻滞剂很敏感,这种阻滞剂在剂量水平下会导致运动障碍,而对正常动物没有影响(Heffner等人,1977年;马歇尔,1979年). 因此,这种方法可以用来揭示神经化学补偿部分受损动物的潜在功能缺陷。
在注射wtrAAV-α-synuclein 8周后,研究低剂量DA合成阻断剂对大鼠的影响(n个=10),突变rAAV-α-synuclein(n个=10)或rAAV-GFP(n个=10),使用前肢步进测试(图。7C). 在用药前3d试验中,所有组均在正常水平下进行(~11步)。第四天,给大鼠单次低剂量TH酶抑制剂α-甲基-对-酪氨酸。6小时后,当TH抑制达到最大值时,α-突触核蛋白转导的动物相对于rAAV-GFP处理的对照组和它们自己的用药前基线都显著受损(对< 0.01). 两个α-突触核蛋白组的效果相似(对>0.05)(组合在图中。7C). 在随后的2天内,正常运动性能逐渐恢复。
讨论
这些结果表明,wt或突变型α-突触核蛋白的过度表达可导致黑质纹状体DA神经元发生进行性神经退行性病变,其特征是α-突触体阳性细胞质和轴突内含物、营养不良和分裂的神经突以及细胞死亡。这些退行性变化是黑质DA神经元特有的,在rAAV-α-突触核蛋白载体有效转导的三个中脑非多巴胺能神经元系统中没有发现。α-突触核蛋白表达的影响似乎是双重的:首先,当α-突触核蛋白呈弥散细胞质分布时(注射后3周),TH酶活性和纹状体DA水平受到抑制(约40–50%),随后黑质DA神经元减少30–80%,与细胞质内含物和营养不良性神经突起的出现以及DA神经元细胞丢失超过50-60%临界阈值的动物出现明显运动障碍相一致。SNc中90%以上的TH阳性神经元由rAAV载体转导。与之前使用该载体的研究一致(Mandel等人,1998年;Bjorklund等人,2000年;Kirik等人,2000年)α-突触核蛋白和GFP转基因在黑质纹状体神经元中长期稳定表达,载体注射27周后仍保持高表达。因为rAAV是一种单链DNA病毒,所以有可能两个cDNA互补的病毒必须感染一个细胞才能成功转导(Ferrari等人,1996年). 这开启了一种可能性,即不同数量的基因组拷贝被插入到给定的细胞中。因此,最终的表达水平可能会因转导细胞而异。这可能很容易解释为什么只有一些转导的SNc神经元退化,以及为什么不同动物的细胞丢失不同。
上一个在体外研究表明,α-synuclein的过度表达会对细胞产生毒性,该蛋白可以与氧化应激和线粒体损伤相互作用,发挥其细胞毒性作用(Ostrerova等人,1999年;Hsu等人,2000年;神田等人,2000年;Tabrizi等人,2000年). 因此,氧化应激以及细胞α-突触核蛋白水平的增加可能会促进α-突触核蛋白的聚集(Giasson等人,2000b;Hsu等人,2000年)从而可能导致线粒体损伤和有毒自由基的生成增加(Ostrerova等人,1999年;Hsu等人,2000年). 此外,Giasson等人(2000b)有报道称,α-synuclein是氧化损伤的靶点,α-sunuclein聚集成有毒内含物可能是由活性氧和氮物种硝化蛋白质引起的。我们的研究结果以及果蝇属(费尼和本德,2000年)表明DA神经元可能特别容易受到α-synuclein过度表达的影响。这可能是因为这些类型的细胞中DA和铁的含量都很高,所以自由基的产生量很高。类似地,氧化应激与线粒体复合物I和α-突触核蛋白聚集水平降低相结合被认为是人类PD中黑质DA神经元选择性死亡的基础(Hsu等人,2000年;大不里士等人,2000年).
这种“双重打击”模型可能解释了以下事实:本研究中α-突触核蛋白过度表达导致了一些但不是所有黑质DA神经元的病理变化和细胞死亡。尽管wt或突变α-突触核蛋白的细胞表达维持在高水平,但持续病理学的迹象,包括α-突触核蛋白阳性内含物和营养不良神经炎,随着时间的推移而消退。这表明,那些在α-突触核蛋白过度表达的最初影响下存活下来的神经元即使在转基因产物的细胞水平保持不变、增加的情况下也能存活和发挥功能。因此,只要细胞不暴露于过度氧化应激或其他可能损害线粒体功能的事件,α-突触核蛋白本身可能对细胞无毒。然而,神经化学数据表明,即使在没有任何明显的病理迹象的情况下,α-突触核蛋白也可能对DA神经元功能产生负面影响。因此,在长期存活的动物中,在载体注射6个月后,纹状体DA水平和纹状体TH酶活性平均降低了40–50%,而纹状体TH阳性神经支配在这个时间点显示出部分恢复(至正常的约75%)。与短期存活动物的观察结果一致,即载体注射后3周,这些数据表明,细胞内α-突触核蛋白水平增加可能对DA合成和储存产生直接抑制作用。
虽然长期以来,氧化应激被认为与PD的发病机制有关,但泛素蛋白体处理系统导致的蛋白质降解功能障碍只是最近才被认为是主要的发病机制。对这种蛋白质降解机制的关注源于多种形式家族性PD的鉴定,其中突变基因:α-突触核蛋白、parkin和泛素C末端水解酶L1(UCHL1)都与蛋白质体降解系统有关(Shimura等人,2000年). 因此,在本研究达到的水平上,黑质DA神经元中α-突触核蛋白的过度表达可能会压倒蛋白质体途径成功处理和清除细胞中多余α-突触核蛋白的能力。此外,α-突触核蛋白的过度表达可能导致细胞内22kDa形式的α-突触核蛋白水平增加(Shimura等人,2001年). 虽然这种糖基化α-突触核蛋白亚型尚未得到很好的表征,但它可能对某些神经元有毒,但对其他神经元则没有毒性(Shimura等人,2000年). 根据该模型,如果所有形式的α-突触核蛋白(wt或突变体)在足够高的水平上表达,或者如果它们受到氧化损伤(例如,通过黑质神经元中DA的氧化代谢),则可能有毒,或者是因为α-突触核蛋白的细胞处理受损(parkin基因突变的患者可能会出现这种情况;Shimura等人,2001年). 这一论点进一步表明,黑质外的非多巴胺能神经元,或黑质内的多巴胺抵抗神经元,可能通过更有效地处理过量的α-突触核蛋白而得到保护。这里看到的α-突触核阳性包涵体没有泛素染色,这一事实与α-突触核蛋白本身可能不是泛素化的底物这一事实相一致(Tofaris等人,2001年). 此外,神经退行性变发生的速度(在本文研究的过度表达模型中为3-8周,而在PD患者中为数月或数年),至少可以部分解释为细胞内α-突触核蛋白的过度表达水平以及细胞处理该蛋白降解的能力。
使用rAAV载体在黑质纹状体系统中靶向α-突触核蛋白过度表达的技术提供了一种新的PD模型,该模型再现了人类疾病的一些主要病理、神经化学和行为特征,因此,这为阐明家族性或特发性PD神经变性的发病机制提供了一个有趣的工具。使用病毒载体过度表达假定的毒性蛋白,如α-突触核蛋白,可能比标准转基因小鼠具有明显优势。首先,在动物生命周期内的任何时候,都可以将病毒载体注射到大脑中的任何特定解剖位置。第二,病毒载体可以在一个半球投与,让另一个半球作为对照,在这里使用的模型系统中,允许使用依赖于行为不对称的功能测试。第三,病毒载体转导的优点是可以在大鼠身上使用,也就是说,在一种比小鼠更适用于行为研究的物种中。最后,病毒载体策略的使用也适用于其他物种,因此也可以研究灵长类动物中α-突触核蛋白过度表达的影响。事实上,使用病毒载体策略的α-突触核蛋白过度表达导致黑质纹状体DA系统的神经病理学和细胞死亡比迄今为止使用转基因技术在小鼠中实现的更为显著。因此,病毒载体战略将增加一个重要的研究工具,以补充其他众所周知的,体内基因修饰策略。
脚注
这项工作得到了瑞典医学研究委员会拨款04X-3874和99-XG-13285、NsGene A/S(丹麦)和国立卫生研究院拨款PO1 NS36302(新墨西哥州)的支持。我们感谢Kerstin Fogelström和Ulla Jarl提供专家技术援助,感谢鲍威尔基因治疗中心载体核心实验室生产载体,感谢Virginia m.Lee博士慷慨提供α-突触核蛋白抗体。
通信地址:瑞典隆德S-221 84 BMC A11沃伦堡神经科学中心Anders Björklund。电子邮件:安德斯.Bjorklund@mphy.lu.se.
参考文献
-
1Arima K、Ueda K、Sunohara N、Hirai S、Izumiyama Y、Tonozuka-Uehara H、Kawai M。帕金森病和路易体痴呆患者路易体丝状成分上NACP/α-突触核蛋白表位的免疫电镜显示。1998年《大脑研究》;808:93–100。doi:10.1016/s0006-8993(98)00734-3。[内政部] [公共医学] [谷歌学者]
-
2Baba M、Nakajo S、Tu PH、Tomita T、Nakaya K、Lee VM、Trojanowski JQ、Iwatsubo T。散发性帕金森病和痴呆患者路易体中α-突触核蛋白的聚集。《美国病理学杂志》。1998;152:879–884.[PMC免费文章] [公共医学] [谷歌学者]
-
三。Bjorklund A、Kirik D、Rosenblad C、Georgievska B、Lundberg C、Mandel RJ。帕金森病的神经保护基因治疗:在大鼠帕金森模型中使用腺病毒、AAV和慢病毒载体将GDNF基因转移到黑质纹状体系统。脑研究,2000年;886:82–98. doi:10.1016/s0006-8993(00)02915-2。[内政部] [公共医学] [谷歌学者]
-
4Braak H,Braak E.帕金森病的病理解剖学。神经学杂志【增刊2】2000;247:3–10. doi:10.1007/PL00007758。[内政部] [公共医学] [谷歌学者]
-
5Conway JE、Zolotukhin S、Muzyczka N、Hayward GS、Byrne BJ。表达Rep和Cap的单纯疱疹病毒1型扩增子完全支持重组腺相关病毒2型的复制和包装。《维罗尔杂志》。1997;71:8780–8789. doi:10.1128/jvi.71.11.8780-8789.1997。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
6Duda JE、Lee VM、Trojanowski JQ。同核蛋白聚集体的神经病理学。神经科学研究杂志2000;61:121–127. doi:10.1002/1097-4547(20000715)61:2<121::AID-JNR1>3.0.CO;2-4.[内政部] [公共医学] [谷歌学者]
-
7Feany MB,本德WW。帕金森氏病果蝇模型。自然。2000;404:394–398. doi:10.1038/35006074。[内政部] [公共医学] [谷歌学者]
-
8Ferrari FK、Samulski T、Shenk T、Samulsbi RJ。二级合成是通过重组腺相关病毒载体进行有效转导的一个速率控制步骤。《维罗尔杂志》。1996;70:3227–3234. doi:10.1128/jvi.70.5.3227-3234.1996。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
9Giasson BI、Jakes R、Goedert M、Duda JE、Leight S、Trojanowski JQ、Lee VM。一组表位特异性抗体检测帕金森氏病路易体中分布在人类α-同核蛋白中的蛋白质结构域。神经科学研究杂志2000a;59:528–533. doi:10.1002/(SICI)1097-4547(20000215)59:4<528::AID-JNR8>3.0.CO;2-0.[内政部] [公共医学] [谷歌学者]
-
10Giasson BI、Duda JE、Murray IV、Chen Q、Souza JM、Hurtig HI、Ischiropoulos H、Trojanowski JQ、Lee VM。在同核蛋白病损害中,通过选择性α-同核蛋白硝化与神经变性相关的氧化损伤。科学。2000亿;290:985–989. doi:10.1126/science.290.5493.985。[内政部] [公共医学] [谷歌学者]
-
11Hayashida K,Oyanagi S,Mizutani Y,Yokochi M。路易体成熟前的早期细胞质变化:一例青少年帕金森病尸检病例中黑质的超微结构研究。神经病理学学报。1993;85:445–448。doi:10.1007/BF00334457。[内政部] [公共医学] [谷歌学者]
-
12Heffner TG,Zigmond MJ,Stricker EM.多巴胺能激动剂和拮抗剂对完整和6-羟多巴胺治疗大鼠喂养的影响。药理学实验与治疗杂志。1977;201:386–399。[公共医学] [谷歌学者]
-
13Hsu LJ、Sagara Y、Arroyo A、Rockenstein E、Sisk A、Mallory M、Wong J、Takenouchi T、Hashimoto M、Masliah E.α-合核蛋白促进线粒体缺陷、氧化应激。《美国病理学杂志》。2000;157:401–410. doi:10.1016/s0002-9440(10)64553-1。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
14Irizarry MC、Growdon W、Gomez-Isla T、Newell K、George JM、Clayton DF、Hyman BT。帕金森病和皮质路易体病中的黑质和皮质路易体以及营养不良的黑质神经突含有α-同核蛋白免疫反应。神经病理学实验神经学杂志。1998;57:334–337. doi:10.1097/00005072-199804000-00005。[内政部] [公共医学] [谷歌学者]
-
15Kahle PJ、Neumann M、Ozmen L、Muller V、Jacobsen H、Schindzielorz A、Okochi M、Leimer U、van Der Putten H、Probst A、Kremmer E、Kretzschmar HA、Haass C。人类和转基因小鼠脑中野生型和帕金森病相关突变体α-同核蛋白的亚细胞定位。神经科学杂志。2000;20:6365–6373. doi:10.1523/JNEUROSCI.20-17-06365.2000。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
16Kanda S,Bishop JF,Eglitis MA,Yang Y,Mouradan MM。α-同核蛋白突变和C末端截断增强了氧化应激的易感性。神经科学。2000;97:279–284. doi:10.1016/s0306-4522(00)00077-4。[内政部] [公共医学] [谷歌学者]
-
17Kirik D,Rosenblad C,Bjorklund A.纹状体内6-羟基多巴胺诱导大鼠黑质纹状体多巴胺系统部分损伤后行为和神经退行性改变的表征。实验神经学。1998;152:259–277. doi:10.1006/exnr.1998.6848。[内政部] [公共医学] [谷歌学者]
-
18Kirik D,Rosenblad C,Bjorklund A,Mandel RJ.帕金森病大鼠模型中GDNF的长期rAAV介导基因转移:纹状体内而非黑质内转导促进受损黑质纹状体系统的功能再生。神经科学杂志。2000;20:4686–4700. doi:10.1523/JNEUROSCI.20-12-04686.2000。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
19Klein RL、Meyer EM、Peel AL、Zolotukhin S、Meyers C、Muzyczka N、King MA。重组腺相关病毒载体在大鼠隔海马或黑质纹状体通路中的神经元特异性转导。实验神经学。1998;150:183–194. doi:10.1006/exnr.1997.6736。[内政部] [公共医学] [谷歌学者]
-
20Kruger R、Kuhn W、Muller T、Woitalla D、Graeber M、Kosel S、Przuntek H、Epplen JT、Schols L、Riess O.Ala30Pro帕金森病α-同核蛋白编码基因突变。自然遗传学。1998;18:106–108. doi:10.1038/ng0298-106。[内政部] [公共医学] [谷歌学者]
-
21Lee CS,Sauer H,Bjorklund A.纹状体内6-羟基多巴胺损伤轴突末端后大鼠多巴胺能神经元变性和运动损伤。神经科学。1996;72:641–653. doi:10.1016/0306-4522(95)00571-4。[内政部] [公共医学] [谷歌学者]
-
22Leroy E、Boyer R、Auburger G、Leube B、Ulm G、Mezey E、Harta G、Brownstein MJ、Jonnalagada S、Chernova T、Dehejia A、Lavedan C、Gasser T、Steinbach PJ、Wilkinson KD、Polymeropoulos MH。帕金森病中的泛素途径。自然。1998;395:451–452. doi:10.1038/26652。[内政部] [公共医学] [谷歌学者]
-
23Mandel RJ,Rendahl KG,Spratt SK,Snyder RO,Cohen LK,Leff SE。帕金森病大鼠模型中人酪氨酸羟化酶和人GTP-环水解酶I纹状体内重组腺相关病毒介导基因转移的特征。神经科学杂志。1998;18:4271–4284。doi:10.1523/JNEUROSCI.18-11-04271.1998。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
24Marshall JF。脑内注射多巴胺耗尽的6-OHDA后的体感失神:自发恢复和药物控制。脑研究,1979年;177:311–324. doi:10.1016/0006-8993(79)90782-0。[内政部] [公共医学] [谷歌学者]
-
25Masliah E、Rockenstein E、Veinbergs I、Mallory M、Hashimoto M、Takeda A、Sagara Y、Sisk A、Mucke L.α-同核蛋白小鼠多巴胺能缺失和包涵体形成:神经退行性疾病的意义。科学。2000;287:1265–1269. doi:10.1126/science.287.5456.1265。[内政部] [公共医学] [谷歌学者]
-
26Matsuoka Y、Vila M、Lincoln S、McCormack A、Picciano M、LaFrancois J、Yu X、Dickson D、Langston WJ、McGowan E、Farrer M、Hardy J、Duff K、Przedborski S、Di Monte DA。在酪氨酸羟化酶启动子驱动的表达人类α-突触核蛋白的转基因小鼠中缺乏黑色素病理学。神经生物学疾病。2001;8:535–539. doi:10.1006/nbdi.2001.0392。[内政部] [公共医学] [谷歌学者]
-
27McLaughlin SK、Collis P、Hermona PL、Muzyczka N.腺相关病毒一般转导载体:前病毒结构分析。《维罗尔杂志》。1988;62:1963–1973. doi:10.1128/jvi.62.6.1963-1973.1988。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
28McNaught KS,Olanow CW,Halliwell B,Isacson O,Jenner P.帕金森病中泛素蛋白酶体系统的失败。Nat Rev神经科学。2001;2:589–594. doi:10.1038/35086067。[内政部] [公共医学] [谷歌学者]
-
29Muzyczka N.将腺相关病毒用作哺乳动物细胞的通用转导载体。当前顶级微生物免疫学。1992;158:97–129. doi:10.1007/978-3-642-75608-5_5。[内政部] [公共医学] [谷歌学者]
-
30Ostrerova N、Petrucelli L、Farrer M、Mehta N、Choi P、Hardy J、Wolozin B.α-同核蛋白与14-3-3蛋白具有物理和功能同源性。神经科学杂志。1999年;19:5782–5791. doi:10.1523/JNEUROSCI.19-14-05782.1999。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
31Polymeropoulos MH、Lavedan C、Leroy E、Ide SE、Dehejia A、Dutra A、Pike B、Root H、Rubenstein J、Boyer R、Stenroos ES、Chandrasekharappa S、Athanassiadou A、Pappetropoulos T、Johnson WG、Lazzarini AM、Duvoisin RC、Di Iorio G、Golbe LI、Nussbaum RL。帕金森病家族中发现的α-突触核蛋白基因突变。科学。1997;276:2045–2047. doi:10.1126/science.276.5321.2045。[内政部] [公共医学] [谷歌学者]
-
32Rathke-Hartlieb S、Kahle PJ、Neumann M、Ozmen L、Haid S、Okochi M、Haass C、Schulz JB。帕金森氏病相关突变α-同核蛋白转基因小鼠对MPTP的敏感性没有增加。神经化学杂志。2001;77:1181–1184. doi:10.1046/j.1471-4159.2001.00366.x。[内政部] [公共医学] [谷歌学者]
-
33Reinhard JF,Jr,Smith GK,Nichol CA。一种基于3H2O释放和[3H]-酪氨酸被木炭吸附的快速、灵敏的酪氨酸-3-单加氧酶测定方法。生命科学。1986;39:2185–2189. doi:10.1016/0024-3205(86)90395-4。[内政部] [公共医学] [谷歌学者]
-
34Schmidt RH,Ingvar M,Lindvall O,Stenevi U,Björklund A.黑质移植物重新神经化纹状体的功能活性:神经递质代谢和[14C]2-脱氧-d-葡萄糖放射自显影术。神经化学杂志。1982;第38:737–748页。doi:10.1111/j.1471-4159.1982.tb08693.x。[内政部] [公共医学] [谷歌学者]
-
35Shimura H、Hattori N、Kubo S、Mizuno Y、Asakawa S、Minoshima S、Shimizu N、Iwai K、Chiba T、Tanaka K、Suzuki T。家族性帕金森病基因产物parkin是一种泛素蛋白连接酶。自然遗传学。2000;25:302–305. doi:10.1038/77060。[内政部] [公共医学] [谷歌学者]
-
36Shimura H、Schlossmacher MG、Hattori N、Frosch MP、Trockenbacher A、Schneider R、Mizuno Y、Kosik KS、Selkoe DJ。parkin从人脑中泛滥成灾一种新型α-突触核蛋白:对帕金森病的影响。科学。2001;293:263–269. doi:10.1126/science.1060627。[内政部] [公共医学] [谷歌学者]
-
37Spilantini MG、Schmidt ML、Lee VM、Trojanowski JQ、Jakes R、Goedert M.路易体中的Alpha-synuclein。自然。1997;388:839–840. doi:10.1038/42166。[内政部] [公共医学] [谷歌学者]
-
38Spilantini MG、Crowther RA、Jakes R、Hasegawa M、Goedert M.帕金森病和路易体痴呆患者路易体丝状内含物中的α-Synuclein。美国国家科学院院刊1998;95:6469–6473. doi:10.1073/pnas.95.11.6469。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
39Tabrizi SJ、Orth M、Wilkinson JM、Taanman JW、Warner TT、Cooper JM、Schapira AH。突变体α-同核蛋白的表达增加了多巴胺毒性的敏感性。人类分子遗传学。2000;9:2683–2689. doi:10.1093/hmg/9.18.2683。[内政部] [公共医学] [谷歌学者]
-
40武田A、Mallory M、Sundsmo M、Honer W、Hansen L、Masliah E。神经退行性疾病中NACP/α-同核蛋白的异常积累。《美国病理学杂志》。1998;152:367–372.[PMC免费文章] [公共医学] [谷歌学者]
-
41Tofaris GK,Layfield R,Spillantini MG.α-同核蛋白代谢,聚集与蛋白酶体的泛素非依赖性降解有关。FEBS信函。2001;509:22–26. doi:10.1016/s0014-5793(01)03115-5。[内政部] [公共医学] [谷歌学者]
-
42van der Putten H、Wiederhold KH、Probst A、Barbieri S、Mistl C、Danner S、Kauffmann S、Hofele K、Spooren WP、Ruegg MA、Lin S、Caroni P、Sommer B、Tolnay M、Bilbe G。表达人α-同核蛋白小鼠的神经病理学。神经科学杂志。2000;20:6021–6029. doi:10.1523/JNEUROSCI.20-16-06021.2000。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
43Xu L,Daly T,Gao C,Flotte TR,Song S,Byrne BJ,Sands MS,Ponder KP。CMV-β-actin启动子比巨细胞病毒或延伸因子1alpha启动子在肝脏中从腺相关病毒载体中引导更高的表达,并导致小鼠中人类因子X的治疗水平。人类基因治疗。2001;12:563–573. doi:10.1089/104303401300042500。[内政部] [公共医学] [谷歌学者]
-
44Zigmond MJ、Abercrombie ED、Berger TW、Grace AA、Stricker EM。中枢多巴胺能神经元损伤后的补偿:一些临床和基本意义。《神经科学趋势》。1990;13:290–296. doi:10.1016/0166-2236(90)90112-n。[内政部] [公共医学] [谷歌学者]
-
45Zolotukhin S、Byrne BJ、Mason E、Zolotukin I、Potter M、Chesnut K、Summerford C、Samulski RJ、Muzyczka N。使用新方法纯化重组腺相关病毒可提高感染滴度和产量。基因疗法。1999年;6:973–985. doi:10.1038/sj.gt.3300938。[内政部] [公共医学] [谷歌学者]

