脑室下区(SVZ)是发育过程中靠近端脑脑室区形成的重要生发层。 这一层在面向发育中的神经节隆起的侧脑室外侧壁中最为突出。 有趣的是,SVZ一直持续到成年( 艾伦,1912年 ; Smart,1961年 ; 奥尔特曼,1969年 ; Sturrock and Smart,1980年 )保持产生神经元和胶质细胞的能力( Lois和Alvarez-Buylla,1993年 ; Kirschenbaum和Goldman,1995年 )。 新生儿( 拉斯金,1993年 )和成年小鼠( Lois和Alvarez-Buylla,1994年 ),SVZ中出生的细胞沿着一条被称为吻侧迁移流(RMS)的受限路径迁移( 奥尔特曼,1969年 )到嗅球,在那里它们分化为颗粒神经元和肾小球周围神经元。 这些细胞以被称为链的细长细胞聚集体的形式迁移( Rousselot等人,1995年 ; Lois等人,1996年 )无需放射状胶质细胞或轴突引导。 最近,我们已经证明,不仅RMS,而且整个SVZ都包含链,这些链形成了一个从侧脑室的尾侧壁延伸到吻侧壁的广泛网络。 移植和显微标记实验表明,在该网络的不同唇腭水平产生的细胞向口腔迁移并加入RMS到达嗅球( Doetsch和Alvarez-Buylla,1996年 )。 神经元前体通过成年哺乳动物SVZ的广泛切向迁移表明,成年神经发生的祖细胞分布在整个细胞壁上。 SVZ的三维地形组织和产生迁移神经元前体的细胞尚未确定。
出生后的SVZ也是胶质生成的场所( Paterson等人,1973年 ; Levison和Goldman,1993年 )、和 在体外 实验表明,多能干细胞位于生发层中(有关综述,请参阅 Alvarez-Buylla和Lois,1995年 ; Calof,1995年 ; 盖奇等人,1995年b ; Weiss等人,1996b )。 与EGF或bFGF共同生长的SVZ保留了自我更新的能力,可以生成神经元和胶质细胞( Reynolds and Weiss,1992年 ; Gritti等人,1996年 ; Johe等人,1996年 )。 成年哺乳动物生发区中存在的神经干细胞增加了将SVZ用作移植前体和损伤或疾病后神经替换前体的可能性。
早期的研究将SVZ描述为未分化形态的深色和浅染细胞、胶质细胞和具有过渡形态的细胞的集合。 有人认为,暗细胞和亮细胞代表胶质细胞产生的不同阶段( Smart,1961年 ; 布莱克莫尔,1969年 ; Privat和Leblond,1972年 )或者它们对应于不同的胶质母细胞谱系( Sturrock and Smart,1980年 )。 然而,这些研究没有考虑到SVZ含有多能干细胞,并且SVZ是神经发生和广泛切向神经元迁移的场所。
在本研究中,我们建立了鉴定成年啮齿动物SVZ细胞类型的超微结构和免疫细胞化学标准。 鉴定出三种主要细胞类型:(1)神经母细胞或A型细胞,对应于先前描述的暗细胞; (2) 星形胶质细胞(B1和B2型细胞),对应于前面描述的光细胞; 和(3)未分化细胞C型细胞,可能与假定的前体细胞相对应。 连续切片重建显示,A型细胞形成由B1型和B2型细胞包裹的切向链,增殖的C型细胞簇与成神经细胞链有局部关联。 在此基础上,我们提出了一个整合SVZ中神经发生和切向迁移的模型。
材料和方法 成年雄性和雌性小鼠用戊巴比妥(Nembutal)进行深度麻醉,并用0.9%生理盐水经心灌注,然后用100ml Karnovsky固定剂(2%多聚甲醛和2.5%戊二醛)进行常规电子显微镜检查,或用100ml 4%多聚甲醛与0.1%戊二醛进行免疫细胞化学检查。 取下头部并在同一固定剂中固定过夜。 然后将大脑从颅骨中取出,并在0.1秒内清洗 米 磷酸盐缓冲液(PB)2小时。
电子显微镜。 在可控震源上切割横向或矢状100μm切片。 将切片在2%锇中后固定2小时,冲洗、脱水,并包埋在Araldite(Durcupan,Fluka BioChemika,Ronkonkoma,NY)中。 为了研究SVZ的整体组织,我们用玻璃刀切割了一系列1.5μm的半薄切片,并用1%甲苯胺蓝染色。 为了鉴定单个细胞类型,用金刚石刀切割超薄(0.05μm)切片,用柠檬酸铅染色,并在Jeol 100CX电子显微镜下进行检查。 所研究的横向半薄和超薄切片包括侧脑室SVZ的整个背腹范围。 为了确定不同类型细胞之间的关系,我们检查了每个SVZ位点50个连续超薄切片。 细胞类型的分类基于整个SVZ中的1800个细胞。 测定了不同细胞类型的横截面积,作为细胞大小的指示(表 1 )。 绘制了连续切片中单个细胞的最大横截面轮廓,并使用美国国立卫生研究院图像分析软件计算了其面积。
表1。
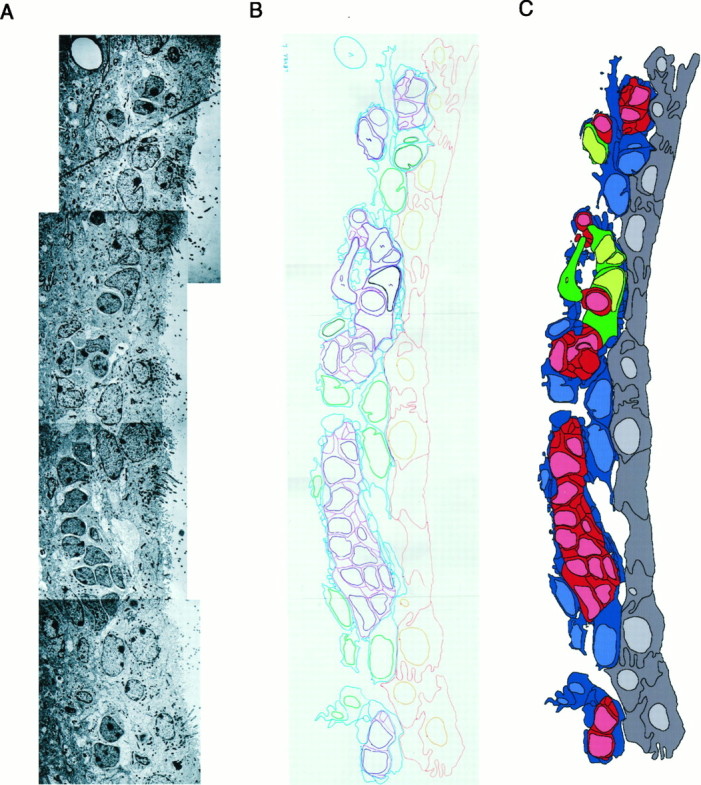
重建。 连续重建侧脑室壁的四个不同区域,并在电子显微镜下进行检查。 三个区域位于外侧壁[背外侧面(15层)、中间面(15级)和腹面(25层)],一个区域位于内侧壁[腹面面(25级)]。 用金刚石刀按以下顺序切割连续切片:一个1.5μm的半薄切片,然后是十个0.05μm的超薄切片。 每个2μm单位(1个半薄切片和10个超薄切片)代表重建中研究的15或25个水平中的一个。 图中显示了背外侧区15个水平中的10个 5 (见下文)。 半薄切片用1%甲苯胺蓝染色; 超薄切片放置在Formvar涂层的单槽格栅上,并用柠檬酸铅染色。 选择四个区域中的每一个区域并拍照(背侧壁,150×30×22μm;中间侧壁,100×30×2 2μm;腹侧壁,80×40×22μm;腹内侧壁,80 x 40×22µm)。 组装每一层的照片蒙太奇,用彩色标记在醋酸纤维板上描绘单个细胞的轮廓(见图。 4 )。 每种细胞类型都用不同的颜色进行追踪。 然后,对侧壁背外侧面研究的15个水平中的10个和侧壁中间面研究的10个水平进行了计算机数字化(数据未显示)。 重建(见图。 4 )通过将复合醋酸盐片从每一层扫描到计算机中来制作。 Adobe Photoshop会高亮显示级别内的单个单元格类型,并使用Adobe Streamline将其转换为矢量地图。 然后用Macromedia Freehand将矢量图组装成每一层的复合重建图。
图5。
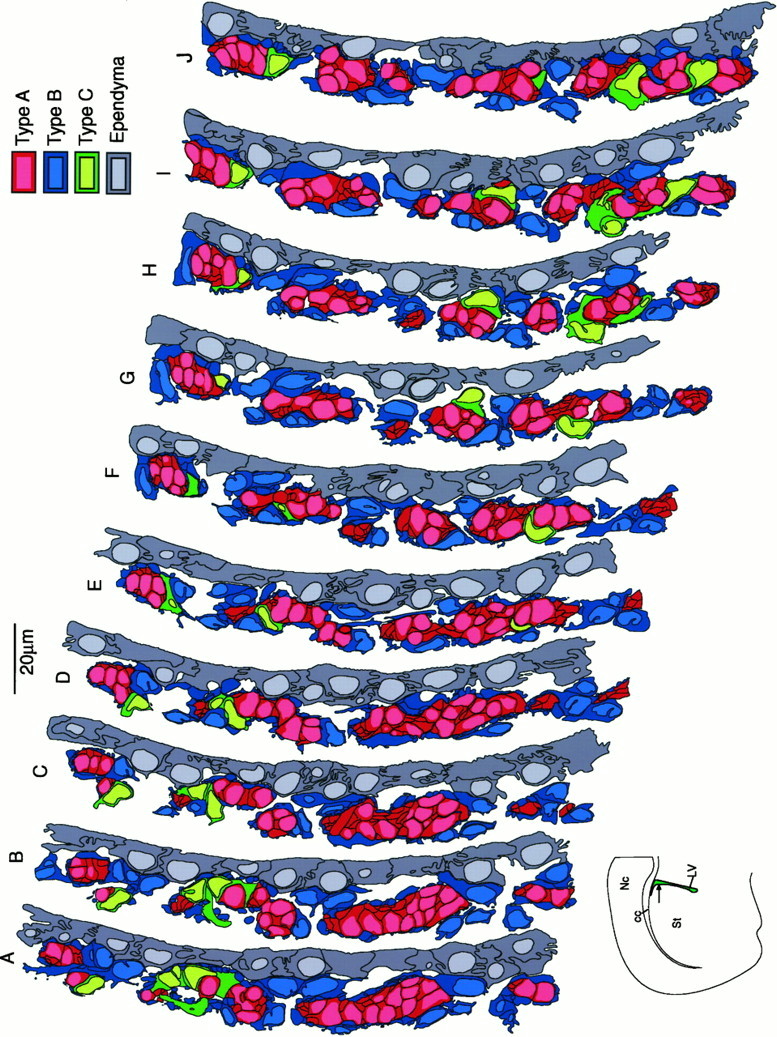
侧脑室侧壁背外侧的连续切片(间隔2μm)重建,水平由 箭头 在冠状半切面上( 左下角 )。 根据文本中描述的特征,在照片蒙太奇中识别细胞类型。 绘制细胞的轮廓和过程,并将其转换为绘图程序,如材料和方法所述,如图所示 4 . 颜色 对应的不同单元类型在 钥匙 在心室的这部分,迁移细胞链(A型细胞)非常丰富。 A型细胞( 红色 )形成分支和收敛的连续链。这些链主要由B型细胞过程覆盖( 蓝色 )通常与C型细胞簇有关( 绿色 )。 空白区域是神经膜密集的区域。 编号 ,新皮质; 复写的副本 胼胝体; 标准 纹状体; 低压 ,侧脑室。 比例尺,20μm。
图4。
提取SVZ单元排列信息的步骤。 A类 ,SVZ的连续电子显微照片被组装成照片蒙太奇。 B类 用不同颜色追踪不同细胞类型的轮廓和细胞核。 C类 ,该信息被传输到计算机中,并按照材料和方法中的描述填充单元格。 这种SVZ表示法可以一目了然地识别细胞类型,并解释图中所示的连续切片重建 5 .
[ 三 H] 胸苷放射自显影术。 5只成年雄性和雌性CD-1小鼠接受1 mCi的50μl注射[ 三 H] 腹腔注射胸腺嘧啶核苷,1小时后处死; 大脑按照常规电子显微镜进行处理。 用玻璃刀切割一系列1.5μm厚的半薄切片,并将其安装在载玻片上,浸于自射线照相乳剂(Kodak NTB2)中,在4°C下暴露4周,在Kodak D-19中显影,并用1%甲苯胺蓝复染。 如果六个或更多的银颗粒覆盖在细胞核上,并且同一个细胞在三个相邻的部分被标记,则认为该细胞被标记。 七十九[ 三 H] 选择在半抗原切片中鉴定的胸苷标记的细胞进行电子显微镜检查。 将半薄切片粘在Araldite块上(Krazy胶),通过反复冷冻(在液氮中)和解冻从玻璃载玻片上分离。 带有扁平半薄切片的块安装在超微切片机中。 用金刚石刀切割超薄切片,并在Jeol 100CX电子显微镜下进行检查,以确定哪些细胞类型并入[ 三 H] 胸腺嘧啶。
细胞计数。 用两种方法估计侧脑室侧壁SVZ中不同细胞类型的比例。 (1) 在连续的超薄切片中鉴定单个细胞,并计算不同类型细胞的数量。 该方法基于连续切片重建,可准确测定细胞数量,不受切片厚度或细胞大小的影响。 (2) 在电子显微镜下计算SVZ(侧壁)背腹范围内不同细胞类型对应的剖面数量。 这项分析是在侧脑室的不同口直肠水平进行的(见表 三 )。 这些数字未经修正,可能会因单元尺寸、方向和截面厚度而产生偏差。 然而,如表所示 2 连续切片重建和单个切片轮廓计数计算出的不同细胞类型的比例非常相似。 这表明,考虑到侧脑室壁中不同细胞类型的大小、几何形状和方向,超薄电镜切片中暴露的轮廓数量是SVZ组成的合理表示。
表3。 侧脑室外侧壁和RMS中SVZ不同口-直肠水平的不同细胞类型的比例
表2。 侧脑室前角外侧壁中不同类型细胞的数量(前角前0–1 mm)
免疫细胞化学。 在振捣器上切割60μm厚的切片,并按照以下步骤进行预包埋免疫染色。
对于聚唾液酸神经黏附细胞分子(PSA–NCAM)免疫染色,振动切片在0.1 米 次碘酸钠 4 10分钟和1%NaBH 4 10分钟,然后在5%二甲基亚砜中培养30分钟。 切片清洗三次,持续10分钟。切片在10%的山羊血清中封闭1小时,在4°C下与1:2000抗脑膜炎球菌B(识别PSA–NCAM;法国爱克斯马赛大学II分校G.Rougon的礼物)孵育48小时,清洗,并在与过氧化物酶结合的二级抗IgM中孵育24小时。 清洗切片,用0.02%二氨基联苯胺(DAB)和0.01%H进行抗体染色 2 O(运行) 2 .
对于TuJ1免疫染色,切片在PBS中超声10秒,在10%的正常马血清中封闭,并在4°C的1:500抗TuJ1抗体(弗吉尼亚大学A.Frankfurter赠送)封闭溶液中孵育48小时。 对切片进行清洗,在1:200抗鼠生物素化二次培养皿(加利福尼亚州伯林盖姆Vector Laboratories,Burlingame,CA)中4°C培养24小时,清洗,在ABC(Vector)中培养4小时,并如上所述用DAB显示。 对于波形蛋白免疫染色,切片在3%BSA中封闭,在4°C下用1:1抗40E-C孵育48小时( Alvarez Builla等人,1987年 )并以上述生物素化二级产物显示。 对于GFAP免疫染色切片,用10%的正常马血清和0.2%的明胶封闭,在4°C的1:400抗GFAP抗体(西格玛,圣路易斯,密苏里州)中培养48小时,并用上述生物素化二级抗体显示。 为了进行巢蛋白免疫染色,将切片在10%马血清中封闭1小时,并在4°C下以1:1的比例在抗鼠401(马里兰州贝塞斯达国立卫生研究院R.McKay的礼物)或1:2000抗巢蛋白130(卡罗林斯卡研究所U.Lendahl的礼物)中孵育48小时。 切片清洗三次,并在1:200抗鼠(anti-401)或1:400抗兔生物素化二级品(anti-nestin 130)中孵育24小时,并如上所述用DAB显示。
免疫染色切片在马来酸缓冲液中清洗,并在0.1%锇溶液中固定 米 PB,脱水,嵌入Araldite(Durcupan,Fluka)。 用玻璃刀切割两微米厚的半薄切片,用1%甲苯胺蓝染色,如上所述重新包埋用于超薄切片,并在Jeol 100CX电子显微镜下检查。
结果 成年小鼠大脑的SVZ是一层不连续的暗细胞和亮细胞,靠近室管膜衬里。 这一层在面向纹状体的侧脑室侧壁最为明显( 米特罗和帕尔科维茨,1981年 )。 面向中隔的内侧壁基本上没有SVZ,但前角最前部除外,在那里可以发现类似于侧壁的暗细胞和亮细胞。 侧脑室顶部几乎没有SVZ。 这项研究的重点是面向纹状体的SVZ,涵盖了该侧壁在多个口-直肠水平的整个背腹范围。
成人SVZ中多种细胞类型共存 下面描述的不同细胞类型的超微结构特征在单个细胞的连续切片重建中测定。 并非所有功能都出现在单个部分中。 表 1 总结了不同细胞类型的超微结构特征。
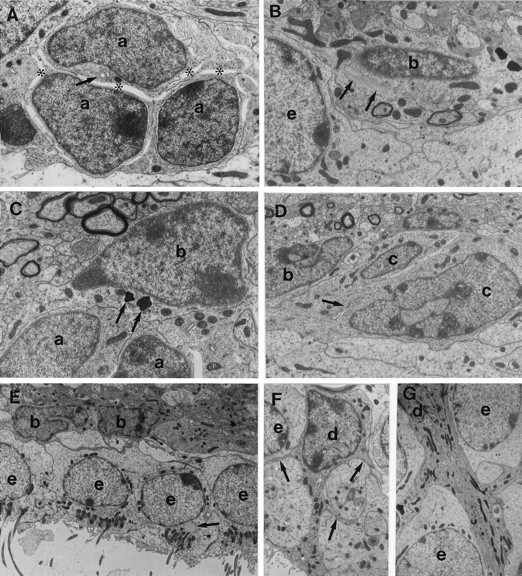
A型细胞对应于光学显微镜下可见的暗细胞。 这些细胞的形态(见图。 1 A类 , 三 )与RMS中描述的迁移神经元前体细胞(A型细胞)相似( Lois等人,1996年 )。 它们的主要特征是具有一个或两个突起的细长细胞体,丰富的疏松染色质,有两到四个小核仁,少而黑的细胞质含有许多游离核糖体,一些短的粗面内质网池(RER),一个小高尔基体,以及沿着细胞长轴方向的许多微管。 A型细胞的细胞核偶尔内陷。 A型细胞轮廓光滑,通过小连接复合体与其他A型细胞相连。 连续切片显示,这些连接复合体是圆形的,直径为0.5–1μm(图。 2 )和分布在细胞表面。 有趣的是,在连接复合体中经常观察到内吞小泡。 这些内吞小泡可能对应于细胞间信号交换的位点,也可能与连接复合体的清除有关( Privat,1974年 )。 在额叶部分,A型细胞的横截面积最小(表 1 ).
图1。
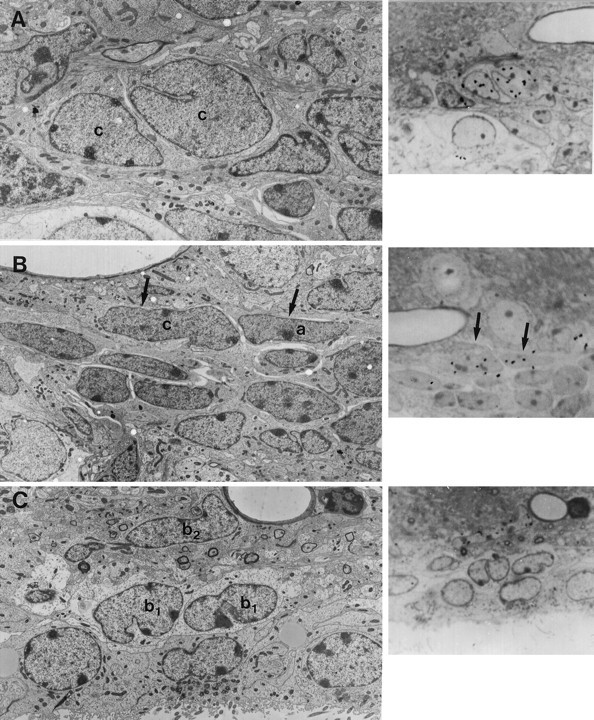
成年小鼠SVZ中的细胞类型。 不同细胞类型的鉴定是在连续重建中进行的,并非所有的超微结构特征都出现在每个显微照片中。 A类 ,三个A类电池( 一 )显示出稀少的黑色细胞质和许多游离核糖体,这是一个小高尔基体( 箭头 )和致密异染色质。 A型电池的表面相对光滑。 在这个横截面上看不到它们的细长形状。 A型细胞经常被开放的细胞外空间分隔开( 星号 )并由特殊的连接复合体连接(见图。 2 )。 放大倍率,10500×。 B类 ,B1型( b条 )细胞的细胞质较轻,胞体致密,中间丝束较粗( 箭头 )以及穿透周围细胞间隙的不规则轮廓。 B1型细胞位于室管膜细胞之间的界面( e(电子) )和覆盖的组织。 放大倍数,13000×。 C类 ,B2型细胞胞质较轻,轮廓不规则,胞质内有致密小体( 箭头 )。 B2型细胞的细胞质中含有少量游离核糖体。 B2型电池( b条 )位于纹状体实质和A型细胞的交界处( 一 )。 注意B2型细胞与纹状体实质有髓和无髓轴突的接触。 放大倍率,8000×。 D类 ,C型电池( c(c) )细胞核不规则,内陷深,染色质疏松,核仁大而网状。 细胞质比B型细胞的电子密度高( b条 )包含一个清晰的高尔基体( 箭头 )。 放大倍数,6500×。 E类 ,室管膜细胞( e(电子) )排列心室,经常有纤毛。 它们严重交叉指状,并含有顶端连接复合体。 细胞核呈圆形,染色质未包被。 它们的细胞质很轻,含有许多基础线粒体和一些游离核糖体。 脂滴( 箭头 )是室管膜细胞所特有的。 注意两个B型电池( b条 )。 放大倍率,3800×。 F类 , G公司 ,坦尼字节( d日 )是与心室接触的黑色无钩细胞。 它们有不规则的细胞核和含有许多线粒体和大型高尔基体的电子致密细胞质。 它们有侧枝( 箭头 )与室管膜细胞交叉的( e(电子) )和B型电池。 放大倍数:6000×in F类 ; 4500×英寸 G公司 .
图3。
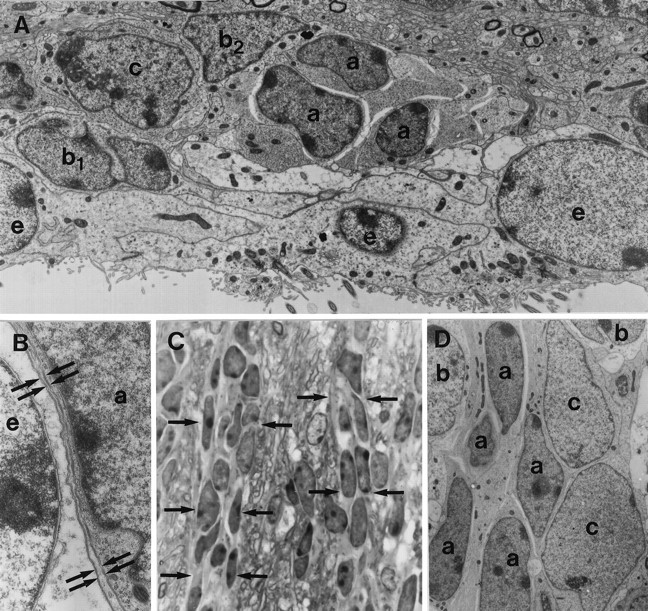
SVZ中主要细胞类型之间的关系。 用连续超薄切片鉴定单个细胞类型。 A类 ,通过侧壁SVZ的正面截面,显示了不同细胞类型之间的地形关系。 A型横向切割链条( 一 )细胞与室管膜细胞分离( e(电子) )通过B1型细胞的电子透明横向扩张( b条 1 )位于室管膜层附近。 相反,B2型细胞的过程( b条 2 )位于纹状体实质附近的基底部,将A型细胞链与周围神经膜隔离。 链内从未发现B型细胞。 注意B2型细胞典型的成簇染色质和B1型细胞的未成簇染色素。 A C型电池( c(c) )其典型的网状核仁位于A型细胞链附近。 放大倍数,6700×。 B类 ,薄片( 箭头 )由B1型细胞的单个或多个过程形成,分离A型细胞( 一 )来自室管膜细胞( e(电子) )。 放大倍数,16800×。 C类 ,通过SVZ的切向切片,在光学显微镜下显示了A型细胞在半薄切片中的链式组织。 这个 箭头 表示存在对应于A型细胞的两条暗细胞链。 放大倍数,360×。 D类 通过SVZ的切向截面显示了该平面上A型细胞的拉长形状及其与C型细胞的紧密联系。 这里可以清楚地看到C型细胞的较大尺寸,以及A型、B型和C型细胞细胞质电子密度的差异。 一 ,A型电池; b条 ,B型电池; c(c) ,C型电池。 放大倍数3500倍。
图2。
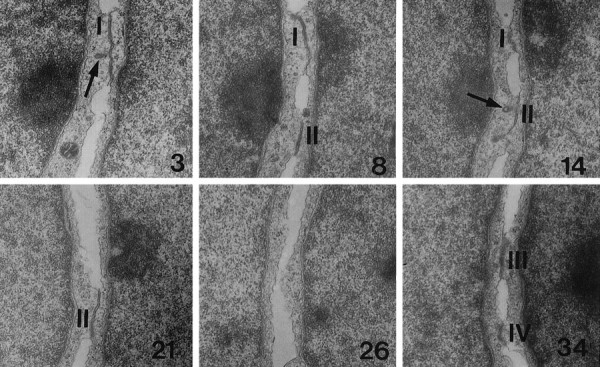
a型细胞之间的小附着带样接触的连续重建的选定切片。 在39个超薄连续切片上拍摄了两个A型细胞之间的连接; 截面号 显示在 右下角 .四个路口( 一、 二、三、四 )出现在这个重建中。 接合处不连续,但呈圆盘状,直径为~0.5–1μm。 内生小泡( 箭头 )通常与这些连接有关。 放大倍数,12500倍。
B型电池(图。 1 B、 C类 )具有不规则的轮廓,大量填充相邻细胞之间的空间。 这些细胞的细胞核不规则,经常有内陷(图。 1 E类 )。 B型细胞的细胞质较轻,含有少量游离核糖体。 B型细胞的显著特征是其丰富的中间丝(图。 1 B类 )细胞质中有致密体(图。 1 C类 )。 B型细胞中有两种亚型,B1型和B2型。 B1型星形胶质细胞较B2型细胞轻,细胞质多,体积大(图。 8 C类 ,表 1 )。 此外,B2型星形胶质细胞细胞核中的染色质呈团块状,而B1型细胞的染色质相对分散(图。 三 A类 )。 B1型细胞的超微结构与RMS中之前描述的B型细胞相似( Lois等人,1996年 )。 B1型细胞最常见于室管膜细胞附近,它们延伸许多突起,形成覆盖室管膜层的薄层。 相反,B2型细胞最常见于纹状体薄壁组织的界面。 尽管这两种细胞类型之间存在差异,但都具有超微结构( Peters等人,1991年 )星形胶质细胞的免疫细胞化学(见下文)特征。
图8。
成年小鼠SVZ中包含[ 三 H] 胸腺嘧啶。 A类 ,一对C型电池( c(c) )注射后1小时标记[ 三 H] 胸腺嘧啶。 这两个细胞细胞核上的银颗粒显示在 正确的 .放大倍数,4500×。 B类 ,两个标记的单元格( 箭头 在半薄截面中 正确的 并在电子显微照片中 左边 )1小时后[ 三 H] 胸腺嘧啶注射对应于C型细胞( c(c) )和a型电池( 一 )。 放大倍数,3400×。 C类 ,B2型细胞也被标记为[ 三 H] 注射后1小时胸腺嘧啶核苷。 在这张电子显微照片中,是一个细长的B2型细胞( b条 2 , 箭头 )用银颗粒标记( 正确的 , 箭头 )。 室管膜细胞( e(电子) )和B1型电池( b条 1 )未观察到用[ 三 H] 胸腺嘧啶。 放大倍数3500倍。
我们区分了第三种细胞类型,我们称之为C型(图。 1 D类 , 三 A、 D类 , 8 A类 )与A型细胞(假定的迁移细胞)和B型细胞(星形胶质细胞)不同。 与A型细胞相比,C型细胞更大、更球形(较少拉长)、更透光,但比B型细胞电子密度更高。 它们的细胞核内陷很深(图。 1 D类 )大多数染色质疏松,尽管有时染色质结块。 C型细胞有一个非典型的大网状核仁(图。 三 A类 )。 它们的细胞质含有一个巨大的高尔基体(图。 1 D类 )与A型细胞相比,核糖体更少,并且没有B型细胞典型的中间丝束。 C型细胞轮廓光滑,常与A型细胞接触。 偶尔,小的连接复合体,类似于A型细胞之间所描述的那些(图。 2 )在C型和A型细胞之间观察到。 C型细胞的大小与B1型细胞相似(表 1 )但流程较少。
单核细胞(D型细胞)在研究区域中很少出现。 这些无钩细胞被夹在室管膜细胞之间,与心室接触,其管腔表面含有微绒毛。 这些细胞在室管膜细胞之间有大量的侧向延伸(图。 1 F、 G公司 )。 细胞核形状不规则,含有深色染色质聚集体。 这些细胞的细胞质电子密度高,细胞器丰富,含有许多线粒体、溶酶体、丰富的RER和独特的高尔基体。
巨大的室管膜(E型)细胞形成了一层上皮单层,将SVZ与心室腔隔开(图。 1 E类 )。 许多研究人员之前已经描述过这些细胞的结构(有关综述,请参阅 Peters等人,1991年 )。 它们的主要区别特征如下:相邻室管膜细胞的外侧突起高度交叉(图。 1 E类 , 三 A类 )包含顶端连接复合体。 暴露于心室的表面含有微绒毛,并且经常有纤毛。 细胞质是电子透明的,含有许多线粒体和位于顶端细胞质中的基体。 它们的细胞核是球形的,染色质没有结块。
此外,SVZ中偶尔可见小胶质细胞、少量固缩细胞、有丝分裂和大神经元。
三维重建揭示了不同细胞类型的地形组织 为了确定不同细胞类型的数量及其相互关系,我们在电子显微镜下重建了SVZ。 侧脑室侧壁在三个背腹位置重建。 第四次重建是内侧壁。 如图所示,追踪并数字化单个细胞的轮廓 4 只显示了一个重建(图。 5 )对应于SVZ背外侧。
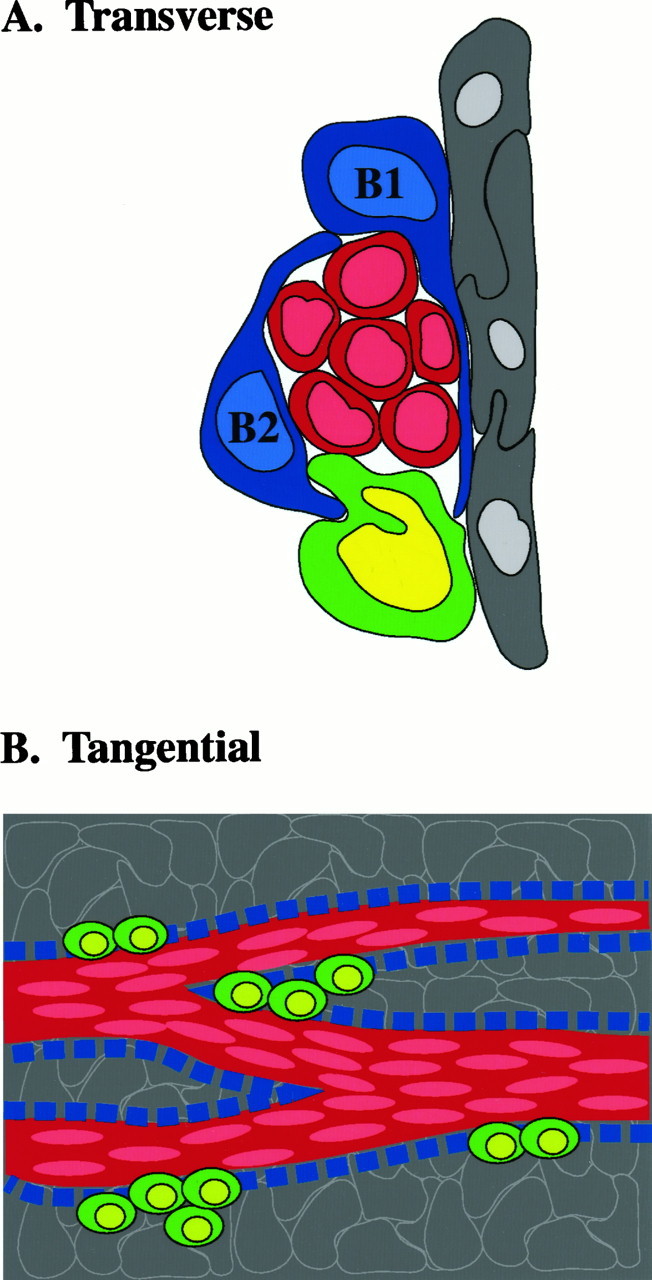
室管膜细胞形成覆盖脑室的连续上皮细胞片(图中深灰色细胞和浅灰色细胞核)。 5 )。 在SVZ的单个超薄横切面上,A型细胞(图中的暗红色细胞和淡红色细胞核)。 5 )出现在1至4个细胞宽(内侧)和2至30个细胞长(背腹)的簇中(图。 三 A、 C类 , 5 )。 然而,当对其进行连续重建检查时,观察到这些A型细胞簇与延伸到多个切片上的细胞链相对应。 这些切线方向的链(平行于侧脑室壁)在穿过SVZ的前后水平时都分裂成较小的链或连接其他链(图。 5 ).
通过B型细胞突起和胞体形成的毡网从纹状体和室管膜层分离出A型细胞链(图中为深蓝色细胞,细胞核为浅蓝色)。 5 )。 B1型细胞膨胀形成的薄层(图。 三 B类 )从室管膜层分离出A型细胞链,而B2型细胞的扩增从上覆的纹状体分离出A型细胞链。 因此,B1型和B2型细胞共同包裹着迁移细胞链。 偶尔,B2型细胞在A型细胞和纹状体之间的鞘中留下小间隙。 相反,A型细胞和室管膜层之间的胶质覆盖层(B1)没有出现间隙。
C型细胞(图中带淡绿色细胞核的绿色细胞。 5 )最常见于小簇中,偶尔个别细胞与A型细胞链接触(图。 三 A、 D类 )。 然而,与A型细胞不同,B型细胞并没有从周围的薄壁组织中分离出C型细胞。 虽然有些细胞部分被B型突起覆盖,但C型细胞同时接触室管膜细胞和纹状体薄壁组织。
对上述重建的一个预测(基于连续的额叶切片)是,在平行于室管膜层的切片中应该可以看到A型细胞链。 在通过SVZ的切向半薄切片中,观察到暗细胞链(图。 三 C类 )。 在电子显微镜下,这些暗细胞对应于A型细胞,可以观察到C型细胞与A型细胞链的紧密联系(图。 三 D类 ).
在SVZ中发现C型细胞,但在RMS中未发现 为了确定SVZ的组成,我们在电子显微镜下计算了前角侧脑室外侧壁超薄切片(角前0、0.5和1 mm)中不同细胞的数量。 此外,我们计算了三次重建中不同细胞类型的数量。 两种方法计算的不同细胞类型的比例非常相似(表 2 )。 A型细胞是最常见的细胞类型,其次是室管膜细胞和B型细胞。 C型细胞占该壁细胞的~10%。 A:B:C型细胞的比例约为3:2:1。
在侧脑室的前角进行上述量化。 因为侧脑室侧壁的SVZ进一步向尾侧延伸到下角( Smart,1961年 ; Doetsch和Alvarez-Buylla,1996年 )在超薄切片中,我们测定了前角尾侧0.5、1.0、1.5和2mm处不同细胞类型的数量。 在研究的所有口腔粘膜水平上均发现A、B和C型细胞。 SVZ尾部的细胞排列与上述类似,A型细胞形成链,被B型细胞包裹(数据未显示)。 如表所示 三 前角尾侧0.5和1mm处的细胞比例与前角相似。 A型和C型细胞被发现位于距前角−2 mm的尾部。 然而,随着B型细胞相对数量的增加,角尾侧−1.5 mm和2.0 mm处的a型和C型细胞数量显著减少。 相反,RMS由A型和B型细胞组成( Jankovski和Sotelo,1996年 ; Lois等人,1996年 ); 在RMS中采样的249个细胞中,没有一个对应于C型细胞(表 三 ).
SVZ不同细胞类型的免疫细胞化学特征 为了进一步确定成年小鼠侧脑室壁的不同细胞类型,我们在光镜和电镜水平上确定了哪些细胞表达PSA–NCAM,这是一种在神经可塑性部位表达的蛋白质( Theodosis等人,1991年 )通过迁移RMS和SVZ中的成神经细胞( Rousselot等人,1995年 ; Doetsch和Alvarez-Buylla,1996年 ); TuJ1,识别年轻神经元的β-微管蛋白标记物( Easter等人,1993年 ; 穆迪等人,1996年 ); 星形胶质细胞标志物GFAP( 比格纳米和达尔,1974年 ); 波形蛋白,一种由前体细胞表达的中间丝( Cocchard和Paulin,1984年 ; Alvarez-Buylla等人,1987年 , 1990 ; Sancho-Tello等人,1995年 ),胶质细胞( Schiffer等人,1986年 ; Cohen等人,1994年 )和一些神经元( Schwob等人,1986年 ); 神经上皮干细胞标记物nestin( Lendahl等人,1990年 )。 表 4 总结了不同细胞类型的免疫染色特征。
表4。
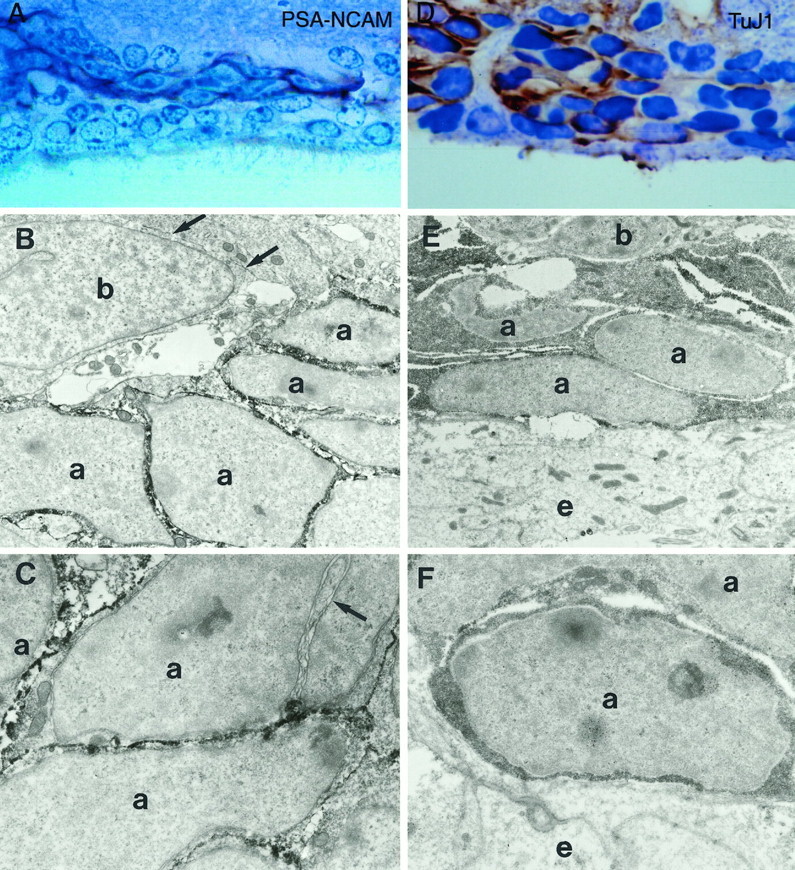
PSA–NCAM和TuJ1免疫染色 A型细胞被抗PSA–NCAM和TuJ1抗体深染(图。 6 A–C )。 PSA–NCAM染色在A型细胞的整个质膜上是连续的,无论细胞面对的是另一个A型细胞还是B型或C型细胞。 TuJ1抗体染色了A型细胞的细胞质(图。 6 D–F )。 PSA–NCAM和TuJ1免疫染色均为阴性的B型和C型细胞以及室管膜细胞。
图6。
SVZ中不同细胞类型的免疫细胞化学特征:PSA–NCAM和TuJ1。 A型细胞链对PSA–NCAM和TuJ1均呈免疫阳性。 其他细胞类型对这些标记物呈免疫阴性。 A类 甲苯胺蓝染色冠状半薄切片中的PSA–NCAM染色显示免疫阳性细胞链两侧为免疫阴性细胞。 放大倍数,400×。 B类 , C类 ,超微结构水平的PSA–NCAM免疫染色。 A型细胞对PSA-NCAM免疫阳性,而B型细胞免疫阴性( 箭头 在里面 B类 )。 PSA–NCAM染色沿A型细胞的质膜连续( 一 ); A型细胞的核膜未染色( 箭头 在里面 C类 )。 放大倍率:8000×in B类 ; 11500×英寸 C类 . D类 甲苯胺蓝染色冠状半薄切片TuJ1免疫染色。 放大倍数,500×。 E类 , F类 ,TuJ1在超微结构水平上的免疫染色。 抗TuJ1抗体使A型细胞的细胞质呈深染。 B型电池( b条 )和室管膜细胞( e(电子) )对这个标记物是免疫阴性的。 放大倍率:6800×英寸 E类 ; 12000×英寸 F类 .
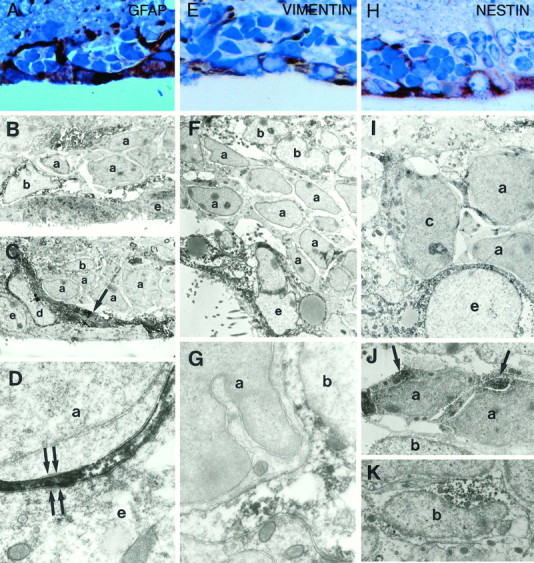
GFAP、波形蛋白和巢蛋白免疫染色 B型细胞被抗GFAP抗体深染(图。 7 A–D )并被波形蛋白抗体轻微染色(图。 7 E–G公司 )。 B1型细胞从室管膜细胞中分离A型细胞链的过程用抗GFAP抗体进行了清晰染色(图。 7 D类 )。 这种染色模式支持我们将这些细胞作为星形胶质细胞的超微结构特征。 室管膜细胞被抗波形蛋白抗体深染(图。 7 E、 F类 )并被抗GFAP抗体轻微染色(图。 7 A、 B、D )。 用抗GFAP强染色的单核细胞(D型细胞)(图。 7 C类 )。 A型和C型细胞对GFAP和波形蛋白均呈阴性。
图7。
SVZ中不同细胞类型的免疫细胞化学特征:GFAP、波形蛋白和巢蛋白。 A型细胞链对GFAP和波形蛋白免疫阴性,但对巢蛋白免疫阳性。 包裹A型细胞链的B型细胞对GFAP、波形蛋白和巢蛋白呈免疫阳性。 室管膜细胞被巢蛋白和波形蛋白抗体染色很强,但被GFAP抗体染色较弱。 图示为GFAP免疫染色( A类 ),波形蛋白( E类 )和嵌套( H(H) )甲苯胺蓝染色的SVZ冠状半薄切片。 A类 ,成串的细胞被GFAP免疫阳性的突起包围。 室管膜细胞也被GFAP抗体染色。 E类 室管膜细胞被抗波形蛋白抗体强烈染色。 星形胶质细胞的突起也呈波形蛋白阳性。 H(H) ,上皮细胞被巢蛋白抗体染色非常暗。 星形胶质细胞和A型细胞(细胞核较深)也可见染色。 放大倍数:300×in A类 ; 400×英寸 E类 ; 400×英寸 H(H) . B类 ,在电子显微镜下,B型细胞的突起和胞体( b条 )被GFAP抗体染色。 这些免疫阳性过程围绕着a型细胞链( 一 )GFAP免疫阴性。 室管膜细胞( e(电子) )被轻微污染。 放大倍数,3000×。 C类 、D型细胞(单核细胞, 箭头 )被GFAP抗体染得很深。 放大倍率,2800×。 D类 、薄层过程( 箭头 ; 另请参见图。 2 E类 )分离A型电池的B1型电池( 一 )来自室管膜细胞( e(电子) )GFAP阳性。 放大倍数,26000×。 F类 ,室管膜细胞( e(电子) )对波形蛋白呈强免疫阳性。 B型细胞对波形蛋白也呈阳性,但染色不象室管膜细胞那样深。 放大倍数,3000×。 G公司 波形蛋白免疫阴性的A型细胞与波形蛋白阳性的B型细胞相邻,放大倍数更高( b条 )细胞质中有深色免疫反应沉淀物。 放大倍数,24500×。 我 室管膜细胞中的巢蛋白免疫反应( e(电子) ),A型电池( 一 )和C型单元格( c(c) )。 nestin抗体未对纹状体神经膜和神经元进行染色。 室管膜细胞的细胞质染色均匀,而A型和C型细胞的染色呈团块状。 放大倍率,5000×。 J型 、A型细胞中巢蛋白免疫反应团( 一 )集中于核周细胞质( 箭头 ; 另请参见 我 )。 放大倍数6000×。 K(K) ,B型电池( b条 )在它们的细胞质中也含有巢蛋白免疫反应物质。 放大倍率,8500×。
两种不同的抗巢蛋白抗体显示出相同的染色模式; 两者都标记为多个单元格类型。 室管膜细胞染色最强烈(图。 7 H、 我 )。 A、B和C型细胞也呈巢式阳性(图。 7 I–K型 )。 A型和C型细胞中的巢蛋白染色在细胞质中呈团块状出现(图。 7 一、 J型 ).
多个单元类型合并[ 三 H] 成年小鼠SVZ中的胸腺嘧啶 细胞在有丝分裂中的位置和超微结构特征表明,SVZ中存在多种细胞类型。 这样我们就可以确定哪些细胞在分裂[ 三 H] 注射胸腺嘧啶核苷,1小时后处死动物。 合并的A、B和C型电池[ 三 H] 胸苷(图。 8 A–C )。 第79页[ 三 H] 电镜观察胸腺嘧啶标记细胞,41(52%)对应C型细胞,12(15%)对应A型细胞,10(12%)对应B型细胞。 16个标记细胞(20%)无法识别。 有趣的是,所有[ 三 H] 胸腺嘧啶标记的B型细胞对应于B2亚类; 没有发现B1星形胶质细胞合并[ 三 H] 胸腺嘧啶。 未观察到标记的室管膜细胞。 [ 三 H] 胸腺嘧啶标记的B型和C型细胞出现在侧脑室侧壁的所有背腹水平。 相比之下[ 三 H] 胸腺嘧啶标记的A型细胞主要位于SVZ的背外侧和腹侧,但不位于中央区域。
讨论 我们的结果显示了SVZ中的多种细胞类型是如何排列的。 主要发现如下:(1)SVZ是围绕成神经细胞链(A型细胞)组织的,这些成神经细胞与侧脑室壁相切;(2)成神经细胞链条是由两种类型的胶质细胞(B1型和B2型)从室管膜细胞和纹状体中分离出来的;(3)增殖前体的热点(C型) 细胞仅与SVZ中的迁移神经母细胞相关,而与RMS中的迁移细胞无关。
A型细胞对应于增殖、迁移的神经元前体( Lois等人,1996年 )。 它们具有与RMS中迁移细胞相同的超微结构和免疫细胞化学特征( 梅内泽斯和卢斯金,1994年 ; Rousselot等人,1995年 ; Jan-kovski和Sotelo,1996年; Lois等人,1996年 )。 本研究中的三维重建表明,A型细胞簇对应于神经元前体链的横截面轮廓,这些前体链是贯穿SVZ的广泛切向链网络的一部分( Doetsch和Alvarez Builla,1996年 )。 对几种哺乳动物SVZ的超微结构研究表明,前额部有黑细胞巢( 布莱克莫尔,1969年 ; 布莱克莫尔和乔利,1972年 ; Privat和Leblond,1972年 )与我们在这里观察到的A型细胞簇非常相似(例如,图。 1 A类 , 三 A类 )。 这表明在其他脊椎动物的SVZ中可能存在类似的迁移前体链网络。
B型细胞的超微结构和免疫细胞化学染色特征表明它们是星形胶质细胞。 除了波形蛋白和GFAP外,B型细胞也表达巢蛋白。 这种中间丝在反应性星形胶质细胞和胶质瘤中有描述( Tohyama等人,1992年 ; 克拉克等人,1994年 )。 与RMS中相同( Lois等人,1996年 )SVZ中的B型细胞形成管状小梁,包裹着a型细胞链,将其与周围的薄壁组织隔离开来。 与RMS不同,SVZ中可以区分两种类型的星形胶质细胞。 B1型星形胶质细胞将A型细胞链从室管膜层中分离出来,而位于基底的B2型星形胶质胶质细胞将A型细胞链与周围的纹状体实质分离出来。 围绕链的星形细胞鞘的功能,包括与室管膜细胞的复杂隔离,尚不清楚。 神经胶质细胞可以沿着某些路径进行物理迁移,并可能阻止细胞接触心室。 鉴于A型细胞在不同的局部环境中穿越的距离很长,胶质鞘也可以为迁移的细胞提供营养支持,并有助于将这些细胞与周围薄壁组织的电气和化学影响隔离开来。
在SVZ中发现了A型细胞链,表明链迁移是成人SVZ的主要组成部分。 在放射状胶质细胞或轴突纤维链的三维重建中,我们没有发现任何证据,这表明,与RMS一样,这些神经元前体正在紧密相连地迁移。 神经元前体不断从SVZ转移到RMS,这表明这些细胞必须被SVZ中常驻前体产生的新细胞所取代。 已从成人SVZ中分离出具有干细胞样特性的细胞。 这些细胞在表皮生长因子(EGF)的存在下生长( Morshead等人,1994年 )或碱性成纤维细胞生长因子(bFGF)( Gritti等人,1995年 )产生球状的细胞簇,称为神经球。 神经层细胞可以分化为神经元、胶质细胞和少突胶质细胞( Reynolds and Weiss,1992年 ),表明它们来自多能性前体,可能与 体内 祖先。 将EGF输注到侧脑室也会导致SVZ中增殖细胞的急剧膨胀( Craig等人,1996年 )。 细胞 体内 对应于 在体外 干细胞尚未鉴定。 确定我们在这里描述的不同细胞类型中,哪些会产生神经球,并且在注入EGF后会扩张,这将是一件有趣的事情。 神经球对巢蛋白免疫阳性,巢蛋白是胚胎中神经上皮干细胞的标志物( Lendahl等人,1990年 )。 尽管在成人SVZ中观察到巢蛋白阳性细胞( Morshead等人,1994年 ; 盖茨等人,1995年 ; Craig等人,1996年 )使用光学显微镜,分辨率不足以确定哪些细胞类型表达这种分子 体内 我们在这里表明,巢蛋白在各种细胞类型中表达,包括室管膜细胞和星形胶质细胞(图。 8 H–K )这表明,在成人大脑中,仅nestin不能作为神经干细胞的唯一标记物。
具有未成熟特征的C型细胞( 罗丹,1974年 )与A型和B型细胞明显不同,在侧脑室外侧壁均有发现,但在RMS中未发现。C型细胞与胶质细胞不对应; 它们轮廓光滑,细胞质未分化,缺乏大束中间丝和精细的突起,GFAP和波形蛋白免疫阴性。 此外,这些大细胞对PSA–NCAM和TuJ1呈免疫阴性,表明C型细胞不同于迁移的成神经细胞。 C型细胞是SVZ中增殖最活跃的细胞(50% 三 [H] 胸腺嘧啶标记细胞为C型细胞),表达中间丝巢蛋白( Lendahl等人,1990年 ),含有一个特征性网状核仁,类似于在其他前体细胞中发现的核仁( 埃尔南德斯·维尔顿,1986年 )。 C型细胞通常以集群的形式出现,与A型细胞的链并列(图。 5 )偶尔与他们形成专业联系。 这些特征表明,C型细胞对应于链中成神经细胞的前体。 胶质细胞也出现在SVZ中( 利维森和高盛,1993年 ; 拉斯金,1993年 )但这些细胞的前体尚不清楚。 虽然新生成的胶质细胞可能来自增殖的B2型细胞,但C型细胞可能同时生成胶质细胞和神经元,因此,与假定的多能性前体细胞相对应。 与A型和B型细胞接触的C型细胞在链外围的位置与这种解释一致。 此外,C型细胞未被B型细胞包裹,其子代可能迁移至脑实质。 虽然我们的结果表明C型细胞对应于未成熟细胞,但C型细胞的起源和命运仍有待确定。
一个相对静止的细胞,位于SVZ内或靠近SVZ,被认为会产生神经球( Morshead等人,1994年 )。 这表明C型细胞不是神经圈的前体细胞,因为它们正在非常积极地分裂。 相反,C型细胞可能对应于相对静止的假定干细胞和迁移的成神经细胞之间的中间增殖群体。 A型和B2型细胞也被标记为[ 三 H] 胸腺嘧啶,将其作为相对静止的SVZ干细胞的候选细胞丢弃。 其余细胞类型——室管膜细胞、B1型细胞和单核细胞——未观察到合并[ 三 H] 但这些细胞具有高度分化的表型,不太可能成为假定的干细胞。 然而,尽管室管膜细胞被认为是终末分化的( Bruni等人,1985年 ),它们表达了最高水平的巢蛋白,如果一些巢蛋白很少分裂,它们就会被一个单独的巢蛋白检测不到[ 三 H] 胸腺嘧啶注射液。 因此,我们的工作并没有排除某些或所有室管膜细胞可能具有分裂能力并表现为干细胞的可能性 体内 或者,有可能产生神经元和神经胶质的细胞 体内 与在EGF或bFGF存在下诱导增殖的细胞不同。 在bFGF存在的情况下,从成年纹状体、海马体和其他不包括SVZ的大脑区域分离出多能细胞( Richards等人,1992年 ; 盖奇等人,1995年 ; Palmer等人,1995年 )或EGF和bFGF( Weiss等人,1996a ).
切向迁移是成人SVZ的主要功能。 在开发早期,SVZ也已证明存在广泛的切向偏移( Rakic和Sidman,1969年 ; 韩礼德和塞普科,1992年 ; O'Rourke等人,1995年 ),但其相应机制尚不清楚。 此处揭示的成人大脑SVZ的神经发生和迁移组织也可能适用于胚胎。 根据A型、B型和C型细胞的排列,我们认为C型细胞团是前体细胞增殖的热点,从而产生链中的成神经细胞(图。 9 )。 这些病灶是子代被喂入链网络的位置,可能指示干细胞分裂的区域。 请注意,C型细胞所在的胶质鞘是开放的,这强烈表明这些是SVZ链网络的入口点。
图9。
成人SVZ的组织简图。 A类 ,迁移神经母细胞链的横截面示意图( 红色 )被两种类型的胶质细胞(B1、B2、, 蓝色 )将迁移细胞与纹状体分开( 左边 )和室管膜细胞( 灰色 )。 C型电池( 绿色 (推测的前体)不被胶质细胞包裹,与迁移的成神经细胞链密切相关。 B类 ,示意图 面对面 从纹状体观察SVZ。 这个 红色通道 代表迁移的成神经细胞链(A型细胞),细胞核呈切线延长( 浅红色 )。 这个 蓝色方块 代表包被胶质细胞(B1和B2型)。 这些细胞形成隧道状结构,A型细胞通过隧道状结构迁移。 推测的前体细胞(C型细胞, 绿色 )与迁移的成神经细胞密切相关,并在小簇中有斑点。 下面的室管膜细胞( 灰色 )在心室表面形成一层薄片。
SVZ中不同细胞类型之间的功能相互作用、它们的谱系关系以及成年SVZ中神经发生的分子决定因素尚待揭示。 在这里,我们提供了成人SVZ中不同细胞类型的超微结构和免疫细胞化学特征,并描述了它们的三维地形组织。 该组织提出了一个迁移和细胞生产如何在SVZ中功能整合的模型,并将有助于定义该区域发生的细胞-细胞相互作用类型。
脚注
F.D.是贝克研究员。 这项工作得到了美国国立卫生研究院(National Institutes of Health)的支持:美国国立儿童健康与人类发展研究院(NICHD-NS32116)和美国国立神经疾病与中风研究所(NINDS-NS28478)。 我们感谢A.Frankfurter博士、G.Rougon博士、U.Lendahl博士和R.McKay博士捐赠的抗体。 我们感谢E.Font、C.Lois和C.Scharff对这份手稿的有益评论。
F.D.和J.M.G.-V.对这项工作做出了同等贡献。
信件应寄至洛克菲勒大学Arturo Alvarez-Buylla博士,地址:纽约州纽约市约克大道1230号,邮编:10021。
参考文献
1 Allen E.白化大鼠中枢神经系统有丝分裂的停止。 《计算机神经学杂志》。 1912; 22:547–568. [ 谷歌学者 ]
2 Altman J.出生后神经发生的放射自显影和组织学研究。 四、 前脑前部的细胞增殖和迁移,特别是嗅球的持续神经发生。 《计算机神经学杂志》。 1969; 137:433–458. doi:10.1002/cne.901370404。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
三。 Alvarez-Buylla A,Lois C.成年脊椎动物大脑中的神经干细胞。 干细胞。 1995; 13:263–272. doi:10.1002/stem.55301307。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
4 Alvarez-Buylla A、Buskirk DR、Nottebohm F。单克隆抗体显示成年鸟类大脑中存在放射状胶质细胞。 《计算机神经学杂志》。 1987; 264:159–170. doi:10.1002/cne.902640203。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
5 Alvarez Builla A,Theelen M,Nottebohm F。成年禽心室区的增殖“热点”揭示了放射状细胞分裂。 神经元。 1990; 5:101–109. doi:10.1016/0896-6273(90)90038-h。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
6 Bignami A,Dahl D.星形胶质细胞特异性蛋白和神经胶质分化。 胶质纤维酸性蛋白抗体的免疫荧光研究。 《计算机神经学杂志》。 1974; 153:27–38. doi:10.1002/cne.901530104。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
7 Blakemore WF。大鼠室管膜下板的超微结构。 J Anat杂志。 1969; 104:423–433. [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
8 Blakemore WF,Jolly DR。狗的室管膜下板和相关室管膜。 超微结构研究。 神经细胞药理学杂志。 1972; 1:69–84. doi:10.1007/BF01098647。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
9 Bruni JE,Del Bigio MR,Clattenburg RE。室管膜:正常和病理。 文献综述。 《大脑研究评论》1985; 9:1–19. doi:10.1016/0165-0173(85)90016-5。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
10 调节脊椎动物神经发生的内在和外在因子。 神经生物电流。 1995; 5:19–27. doi:10.1016/0959-4388(95)80082-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
11 Clarke SR、Shetty AK、Bradley JL、Turner DA。反应性星形胶质细胞表达中间神经丝巢蛋白。 神经报告。 1994; 5:1885–1888. doi:10.1097/00001756-199410000-00011。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
12 Cochard P,Paulin D。体内小鼠胚胎中枢和外周神经系统中神经丝和波形蛋白的初始表达。 神经科学杂志。 1984; 4:2080–2094. doi:10.1523/JNEUROSCI.04-08-02080.1984。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
13 Cohen I、Sivron T、Lavie V、Blaugrund E、Schwartz M.鱼视神经中的波形蛋白免疫反应胶质细胞:再生的意义。 格利亚。 1994; 10:16–29. doi:10.1002/glia.440100104。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
14 Craig CG、Tropepe V、Morshead CM、Reynolds BA、Weiss S、Van der Kooy D。成年小鼠脑内内源性室管膜下神经前体细胞群的体内生长因子扩增。 神经科学杂志。 1996; 16:2649–2658. doi:10.1523/JNEUROSCI.16-08-02649.1996。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
15 Doetsch F,Alvarez-Buylla A.成年哺乳动物大脑中神经元迁移切线路径网络。 美国国家科学院院刊,1996年; 93:14895–14900. doi:10.1073/pnas.93.25.14895。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
16 Easter SS,Ross LS,Frankfurter A.小鼠大脑中的初始轨迹形成。 神经科学杂志。 1993; 13:285–299. doi:10.1523/JNEUROSCI.13-01-00285.1993。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
17 Gage FH、Coates PW、Palmer TD、Kuhn HG、Fisher LJ、Suhonen JO、Peterson DA、Suhr ST、Ray J.成年祖细胞移植到成人大脑的存活和分化。 美国国家科学院院刊,1995a; 92:11879–11883. doi:10.1073/pnas.92.25.11879。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
18 Gage FH、Ray J、Fisher LJ。 从中枢神经系统中分离、鉴定和使用干细胞。 《神经科学年鉴》。 1995年b; 18:159–192. doi:10.1146/annurev.ne.18.030195.001111。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
19 Gates MA、Thomas LB、Howard EM、Laywell ED、Sajin B、Faissner A、Gotz B、Silver J、Steindler DA。大脑半球发育中和成年小鼠室下区的细胞和分子分析。 《计算机神经学杂志》。 1995; 361:249–266. doi:10.1002/cne.903610205。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
20 Gritti A、Cova L、Parati EA、Galli R、Vescovi AL。碱性成纤维细胞生长因子支持成年小鼠中枢神经系统表皮生长因子生成的神经元前体细胞的增殖。 神经科学快报。 1995; 185:151–154. doi:10.1016/0304-3940(95)11247-t。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
21 Gritti A、Parati EA、Cova L、Frolichsthal P、Galii R、Wanke E、Faravelli L、Morassutti DJ、Roisen F、Nickel DD、Vescovi AL。来自成年小鼠大脑的多潜能干细胞在碱性成纤维细胞生长因子的作用下增殖和自我更新。 神经科学杂志。 1996; 16:1091–1100. doi:10.1523/JNEUROSCI.16-03-01091.1996。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
22 Halliday AL,Cepko CL.发育中纹状体细胞的生成和迁移。 神经元。 1992; 9:15–26. doi:10.1016/0896-6273(92)90216-z。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
23 Hernandez-Verdun D.哺乳动物细胞中核仁的结构组织。 收件人:Jasmin G,Simard R,编辑。 核亚显微检查。 卡格尔; 巴塞尔:1986年。 第26-62页。 [ 公共医学 ] [ 谷歌学者 ]
24 Jankovski A,Sotelo C.成年小鼠脑室下区-嗅球迁移途径:通过异慢性和异位移植确定的细胞组成和特异性。 《计算机神经学杂志》。 1996; 371:376–396. doi:10.1002/(SICI)1096-9861(19960729)371:3<376::AID-CNE3>3.0.CO; 2-#. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
25 Johe KK、Hazel TG、Muller T、Dugich-Djordjevic MM、McKay RDG。 单一因素指导胚胎和成人中枢神经系统干细胞的分化。 基因开发1996; 10:3129–3140. doi:10.1101/gad.10.24.3129。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
26 Kirschenbaum B,Goldman SA。脑源性神经营养因子促进成年大鼠前脑室管膜下区神经元的存活。 美国国家科学院院刊,1995年; 92:210–214. doi:10.1073/pnas.92.1.210。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
27 Lendahl U、Zimmerman LB、McKay RDG。 CNS干细胞表达一类新的中间丝蛋白。 单元格。 1990; 60:585–595. doi:10.1016/0092-8674(90)90662-x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
28 Levison SW,Goldman JE。少突胶质细胞和星形胶质细胞均由出生后大鼠前脑脑室下区的祖细胞发育而来。 神经元。 1993; 10:201–212. doi:10.1016/0896-6273(93)90311-e。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
29 Lois C,Alvarez-Buylla A.成年哺乳动物前脑脑室下区的增殖细胞可以分化为神经元和胶质细胞。 美国国家科学院院刊,1993年; 90:2074–2077. doi:10.1073/pnas.90.5.2074。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
30 Lois C,Alvarez-Buylla A.成年哺乳动物大脑中的长距离神经元迁移。 科学。 1994; 264:1145–1148. doi:10.1126/science.8178174。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
31 Lois C,Garcia-Verdugo JM,Alvarez-Buylla A.神经元前体的链迁移。 科学。 1996; 271:978–981。 doi:10.1126/science.271.5251.978。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
32 拉斯金·MB。 来自前脑脑室下区的出生后生成神经元的增殖和迁移受限。 神经元。 1993; 11:173–189. doi:10.1016/0896-6273(93)90281-u。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
33 Menezes JRL,Luskin MB。 神经元特异性微管蛋白的表达定义了发育中端脑增殖层中的一个新群体。 神经科学杂志。 1994; 14:5399–5416. doi:10.1523/JNEUROSCI.14-09-05399.1994。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
34 Mitro A,Palkovits M.大鼠脑室、室管膜和室周结构的形态学。 作者:Lierse W,编辑。 解剖图书馆,第21期。 卡格尔; 巴塞尔:1981年。 [ 公共医学 ] [ 谷歌学者 ]
35 Moody SA,Miller V,Spanos A,Frankfurter A.青蛙(非洲爪蟾)中神经元特异性β-微管蛋白的发育表达:胚胎期轴突生长的标志。 《计算机神经学杂志》。 1996; 364:219–230. doi:10.1002/(SICI)1096-9861(19960108)364:2<219::AID-CNE3>3.0.CO; 2-8. [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
36 Morshead CM、Reynolds BA、Craig CG、McBurney MW、Staines WA、Morassutti D、Weiss S、Van der Kooy D。成年哺乳动物前脑中的神经干细胞:室管膜下细胞的相对静止亚群。 神经元。 1994; 13:1071–1082. doi:10.1016/0896-6273(94)90046-9。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
37 O'Rourke NA,Sullivan DP,Kaznowski CE,Jacobs AA,McConnell SK。发育中大脑皮层神经元的切向迁移。 1995年开发(Camb); 121:2165–2176. doi:10.1242/dev.121.7.2165。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
38 Palmer TD、Ray J、Gage FH。 FGF-2反应性神经元祖细胞位于成年啮齿动物大脑的增殖区和静止区。 分子细胞神经科学。 1995; 6:474–486. doi:10.1006/mcne.1995.1035。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
39 Paterson JA,Privat A,Ling EA,Leblond CP。半薄切片中胶质细胞的研究。 三、 年轻大鼠侧脑室注射3H-胸腺嘧啶核苷后,放射自显影显示室管膜下细胞转化为胶质细胞。 《计算机神经学杂志》。 1973; 149:83–102. doi:10.1002/cne.91490106。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
40 Peters E、Palay SL、Webster H de F。神经系统的精细结构:神经元及其支持细胞。 牛津大学UP; 纽约:1991年。 [ 谷歌学者 ]
41 Privat A.生长中大鼠脑内附着板再吸收的一种可能机制。 《大脑研究》1974; 69:125–129. doi:10.1016/0006-8993(74)90376-x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
42 Privat A,Leblond CP。幼年大鼠大脑的室管膜下层和邻近区域。 《计算机神经学杂志》。 1972; 146:277–302. doi:10.1002/cne.901460302。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
43 Rakic P,Sidman RL。胎儿人脑中枕叶神经元的端脑起源。 Z Anat Entwicklungsgesch公司。 1969; 129:53–82. doi:10.1007/BF00521955。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
44 Reynolds B,Weiss S.从成年哺乳动物中枢神经系统的分离细胞中生成神经元和星形胶质细胞。 科学。 1992; 255:1707–1710. doi:10.1126/science.1553558。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
45 罗丹JAG。 组织学。 牛津大学UP; 纽约:1974年。 [ 谷歌学者 ]
46 Richards LJ、Kilpatrick TJ、Bartlett PF。成年小鼠大脑神经元细胞的从头生成。 美国国家科学院院刊1992; 89:8591–8595. doi:10.1073/pnas.89.18.8591。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
47 Rousselot P、Lois C、Alvarez-Buylla A.胚胎(PSA)N-CAM揭示了成年小鼠侧脑室和嗅球之间迁移的成神经细胞链。 《计算机神经学杂志》。 1995; 351:51–61。 doi:10.1002/cne.90351006。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
48 Sancho-Tello M,Vallés s,Montoliu C,Renau-Piqueras J,Guerri C。大鼠脑和放射状胶质细胞培养中GFAP和波形蛋白基因表达的发育模式。 格利亚。 1995; 15:157–166. doi:10.1002/glia.440150208。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
49 Schiffer D、Giordana MT、Migheli A、Giaccone G、Pezzotta S、Mauro A.大鼠脑实验性胶质反应中的胶质纤维酸性蛋白和波形蛋白。 1986年《大脑研究》; 374:110–118. doi:10.1016/0006-8993(86)90399-9。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
50 Schwob JE、Farber NB、Gottlieb DI。 成年大鼠嗅上皮的神经元含有波形蛋白。 神经科学杂志。 1986; 6:208–217。 doi:10.1523/JNEUROSCI.06-01-00208.1986。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
51 Smart I.注射胸腺嘧啶-H3后放射自显影显示的小鼠大脑室管膜下层及其细胞生成。 《计算机神经学杂志》。 1961; 116:325–348. [ 谷歌学者 ]
52 Sturrock RR,智能IHM。 小鼠胚胎至老年室管膜下层的形态学研究。 J Anat杂志。 1980; 130:391–415. [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
53 Theodosis DT,Rougon G,Poulain DA。具有可塑性的成年神经元系统对胚胎特征的保留:下丘脑神经垂体系统中的多唾液酸化神经细胞粘附分子。 美国国家科学院院刊1991; 88:5494–5498. doi:10.1073/pnas.88.13.5494。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
54 Tohyama T、Lee VM-Y、Rorke LB、Marvin M、McKay RDG、Trojanowski JQ。 巢蛋白在胚胎人神经上皮和人神经上皮肿瘤细胞中的表达。 实验室投资。 1992; 66:303–313. [ 公共医学 ] [ 谷歌学者 ]
55 Weiss S、Dunne C、Hewson J、Wohl C、Wheatley M、Peterson AC、Reynolds BA。成年哺乳动物脊髓和心室神经轴中存在多功能CNS干细胞。 神经科学杂志。 1996年a; 16:7599–7609. doi:10.1523/JNEUROSCI.16-23-07599.1996。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
56 Weiss S、Reynolds BA、Vescovi AL、Morshead C、Craig CG、Van der Kooy D。哺乳动物前脑中有神经干细胞吗? 《神经科学趋势》。 1996年b; 19:387–393. doi:10.1016/s0166-2236(96)10035-7。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]