结果和讨论
衣康酸盐治疗在巨噬细胞激活过程中具有抗炎作用
衣康酸的产生是巨噬细胞在转录和代谢水平上活化的标志之一:Irg1是活化巨噬细胞中诱导程度最高的酶之一,衣康酸在细胞内大量积累(图1A). 活化骨髓源巨噬细胞(BMDM)培养上清液的代谢组学分析显示,如前所述,也分泌了衣康酸(斯特雷尔科等人,2011年) (图1B). 衣康酸的产量促使我们调查其潜在的调节作用。
图1。衣康酸盐对巨噬细胞活化具有抗炎作用。
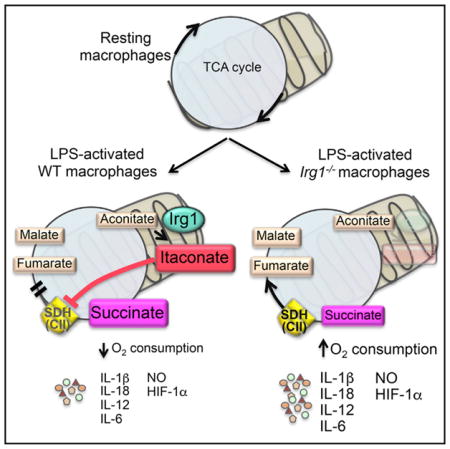
(A) 火山图显示了静息和激活BMDM(LPS+IFN-γ,24小时)之间差异表达的转录物(左)和代谢物(右)。
(B) 激活后指定时间点BMDM细胞内和分泌的衣康酸的相对表达(LPS+IFN-γ)。
(C) 通过流式细胞术测定未经治疗或注射DI-的BMDM(0.25 mM,12小时)中细胞内iNOS表达的直方图,然后用LPS+IFN-γ刺激(24小时)。
(D) (C)BMDM培养上清液中的IL-12水平。
(E) 未经治疗或DI-治疗的BMDM分泌IL-6和TNF-α,并用LPS刺激(24小时)。
(F) 所选炎症标记基因非刺激(Uns)、LPS刺激(LPS)、DI-去甲肾上腺素(DI+Uns)和DI-去乙肾上腺素以及LPS刺激的BMDM的热图(4小时)。
(G) 未经治疗的BMDM或DI-预处理的BMDM分泌的成熟IL-1β和IL-18,然后用LPS(4小时)和ATP(45分钟)刺激。
(H) 未经治疗的BMDM或DI-预处理的BMDM分泌的成熟IL-1β水平,并用LPS和尼葛霉素(Nig.)或尿酸单钠晶体(MSU)刺激。
(一) Western blot分析未经治疗或DI-pretrated和刺激的BMDM裂解液中的NLRP3、pro-IL-1β和ASC,如(G)所示。α-微管蛋白作为负荷对照。所示斑点是两个独立实验的代表。
对于(B)、(E)、(G)和(H),数据显示为平均值±SEM(n=3)。使用双尾Student t检验(B和E)或单向方差分析(ANOVA)计算p值,并与未经处理的刺激细胞(G和H)进行比较*p<0.05**p<0.01***p<0.001****p<0.0001。另请参见图S1.
我们首先测试了外源性添加衣康酸对LPS或LPS+IFN-γ刺激后诱导的炎症反应的影响。我们用生理相关剂量的衣康酸二甲酯(DI)治疗小鼠BMDM(图S1A)衣康酸的一种膜渗透性非离子形式。DI预处理抑制iNOS蛋白表达(图1C)IL-12p70和IL-6分泌(图1D和1E)从而干扰促炎巨噬细胞的激活。相反,TNF-α水平保持不变(图1E)表明DI-治疗的效果不是由于NF-κB依赖性基因表达的整体抑制。为了确定衣康酸影响的特定途径,我们通过RNA-seq对用DI或载体预处理的BMDM进行全局转录谱分析,然后用LPS刺激。差异基因表达证实DI-治疗导致炎症前转录物谱的下调(图1F和S1B级),包括2个,伊尔6、和伊利12b.
RNA-seq分析还显示,DI-预处理调节了参与炎症小体激活和功能的几个LPS调节基因的表达(图1F),包括伊尔1b,18岁,第2页第7页,案例1、和Pycard公司(ASC)。事实上,DI有效地抑制了在典型NLRP3激活条件下诱导的成熟IL-1β和IL-18的生成,即LPS驱动启动(信号1),然后是信号2诱导物ATP、尼日尔金和尿酸单钠晶体(图1G和1H),而它仅适度影响炎症体诱导的细胞毒性(图S1C). DI-治疗的BMDM在AIM-2依赖性炎症小体激活时也会降低IL-1β的产生(图S1D)这表明对炎症组有更广泛的调节作用。DI-处理细胞中IL-1β原、ASC和NLRP3的蛋白表达降低(图1I)表明衣康酸介导的炎症小体功能抑制主要是由于启动期缺陷所致。
鉴于衣康酸的这种抗炎作用,并鉴于其对细胞外细菌的杀灭能力,我们接下来确定了它在巨噬细胞感染期间对细胞内细菌的影响。为此,我们用鼠伤寒沙门菌是一种细胞内革兰氏阴性细菌,可触发TLR4信号和NLRP3炎症小体激活(Broz等人,2010年)有无DI预处理。与我们之前的研究结果一致,感染诱导的IL-1β、IL-6和一氧化氮(NO),而不是TNF-α,在DI-处理的细胞中被消除(图S1E)而DI-处理的BMDM和对照BMDM的细胞内细菌数量相当(图S1F)表明非细胞毒性浓度的衣康酸的抗炎作用不是直接杀菌作用的结果。相反,依他康酸可能具有调节作用,正如病毒感染背景下Irg1诱导所支持的那样(Cho等人,2013年) (图S1G),表明其具有非抗菌反应特有的调节功能。
外源衣康酸在体内外抑制琥珀酸脱氢酶
细胞代谢的扰动可能导致IL-1β生成和炎症小体激活的转录缺陷(Moon等人,2015年)我们假设衣康酸通过干扰细胞代谢发挥抗炎作用。使用转录数据的计算分析(Becker和Palsson,2008年),我们研究了在没有脂多糖的情况下,由衣康酸触发的代谢流量的可能重组。使用通量平衡分析框架,我们扩展了最初为RAW 264.7巨噬细胞系制定的代谢模型(Bordbar等人,2012年)通过包括原始模型中没有的几个反应和酶(例如Irg1和衣康酸,有关详细信息,请参阅实验程序)。我们在未经处理和经衣康酸处理的条件下寻找与RNA-seq数据最为一致的通量(见实验程序)。一个比较网络强调了三种类型的代谢流量变化对衣康酸治疗的反应(图2A和S2A公司):代谢通量降低(蓝色边缘),代谢通量增加(红色),以及对添加衣康酸盐不敏感的反应(灰色)。
图2。衣康酸抑制体内外Sdh活性并调节缺血再灌注损伤期间ROS介导的组织损伤。
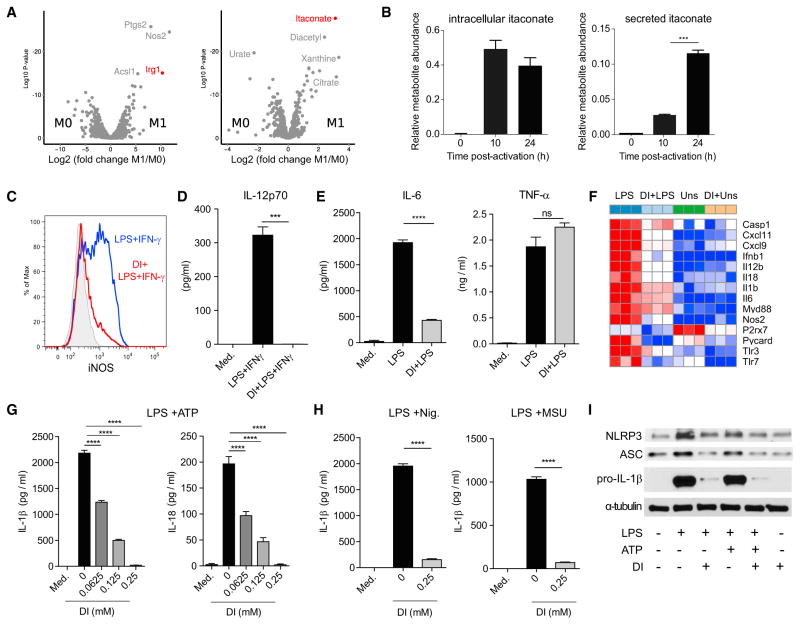
(A) 比较网络显示了使用和不使用依他康酸治疗的未刺激巨噬细胞之间预测流量大小的变化。
(B) 在DI-处理或未处理的BMDM中测量的细胞外酸化率(ECAR)。数据显示为两个代表性实验之一的10-15个重复的平均值±SEM。
(C) 琥珀酸、丙二酸和衣康酸的化学结构。
(D) 静息期BMDM治疗前后的相对琥珀酸水平。数据显示为平均值±SEM(n=3)。p值采用双尾Student t检验计算。
(E) Lineweaver-Burk图(左)、Dixon图(右)和计算的K米和K我值。所示数据为平均值±SD(L-B图,n=3)和平均值(Dixon图,n=3)。
(F) 经DI或生理盐水治疗后,IR损伤心脏的代表性Evans Blue和TTC染色切片。
(G和H)面积-风险(AAR)和梗死面积(IA)的定量,作为AAR(G)和左心室(LV)心肌(H)的百分比(生理盐水,n=8;DI,n=7)。p值采用双尾Student t检验计算。
(I) 在DI或稀释剂存在下缺氧24小时的新生大鼠心肌细胞(NRCM)中ROS的百分比变化(相对于各自的常氧对照)。
(J) 按(H)处理的NRCM中的细胞死亡百分比。对于(I)和(J),在单因素方差分析(n=8/条件)后,使用事后检验计算p值。
(K) 使用MitoSox染料检测BMDM中mROS表达的直方图,BMDM经DI预处理或未经DI处理,并用LPS激活(3小时)。
(五十) mROS平均荧光强度(相对于介质)的折叠变化,单位为(K)。
数据显示为平均值±SEM(n=3,每个重复)。p值采用双尾Student t检验计算*p<0.05**p<0.01***p<0.001。另请参见图S2.
计算分析的两个特征显而易见。首先,预计添加衣康酸会增加乳酸脱氢酶(Ldh)的生成。我们使用Seahorse技术对未经模拟的BMDM进行实验验证,包括DI治疗和未经DI治疗。我们观察到,细胞外酸化率(ECAR)在衣康酸的存在下增加,这是培养基中乳酸积累的结果(图2B). 第二个预测是,添加衣康酸会降低通过Sdh的代谢流量。这表明,衣康酸可能具有竞争性地抑制Sdh,这可能是因为衣康酸、琥珀酸和丙二酸之间的结构相似,丙二酸是已知的Sdh抑制剂(图2C). 事实上,67年前,基于丙二酸盐和衣康酸盐对线粒体功能的类似作用,人们推测衣康酸对脱氢酶有抑制作用(阿克曼和波特,1949年). 为了评估这一假设,我们比较了纯化的Sdh在有或无依他康酸的情况下的活性。值得注意的是,当琥珀酸的使用浓度接近生理浓度1 mM时,衣康酸剂量依赖性地阻断了Sdh的活性(Bennett等人,2009年) (图S2B)和依他康酸治疗静息期BMDM导致琥珀酸水平升高(图2D). 动力学分析证实了衣康酸盐抑制Sdh的竞争模式(图2E,左),Dixon图分析显示K我对于衣康酸盐0.22 mM(图2E,右侧)。计算的K米琥珀酸的值为0.29±0.8 mM。这些结果与24小时LPS激活的巨噬细胞内琥珀酸和衣康酸的摩尔量一致(图S2C). 这些数据表明,衣康酸对BMDM的抗炎作用可能是由于抑制Sdh。支持这一观点,丙二酸二甲酯预处理也抑制LPS+ATP刺激后IL-1β的生成(图S2D).
接下来,我们评估了衣康酸是否在体内抑制Sdh活性。最近的一项研究(Chouchani等人,2014年)强调了琥珀酸氧化在心脏缺血再灌注损伤中的促炎作用。在IR损伤期间,线粒体中可能会产生mROS,这是逆向电子传递(RET)的结果,即缺血期间积累的琥珀酸为Sdh提供燃料。因此,丙二酸二甲酯对Sdh的抑制限制了IR损伤和mROS水平(Chouchani等人,2014年). 我们使用这个模型来测试衣康酸盐是否会在体内抑制Sdh介导的氧化。缺血期间静脉输注DI可显著缩小心肌梗死面积(图2F和2G). 虽然两组的面积风险相似(图2H),DI治疗后梗死面积减少42%与丙二酸二甲酯治疗后的梗死面积减少相当(Chouchani等人,2014年),表明了一种共同的作用机制。为了评估这一假设,我们使用了一种体外试验,该试验通过缺氧损伤新生心肌细胞来模拟心肌梗死损伤(Ma等人,2012年). DI预处理减弱了低氧诱导的ROS生成增加(图2I)并对低氧诱导的细胞死亡提供剂量依赖性保护(图2J).
因此,我们评估了衣康酸介导的Sdh抑制是否影响mROS的生成。DI预处理降低BMDM上调mROS对LPS的反应能力(图2K和2L). 因为干扰mROS会影响炎症小体启动(Bauernfeind等人,2011年),钝化的mROS反应在机制上与Sdh抑制和衣康酸对IL-1β和IL-18生成的抗炎作用有关(图1G). 此外,这表明琥珀酸处理而不是积累对巨噬细胞的炎症重组很重要。
内源性衣康酸调节炎症巨噬细胞的代谢重塑、琥珀酸水平和线粒体呼吸
为了测试内源性衣康酸的生理相关性,我们用靶向性破坏红外线1基因(图S3). 作为来自的BMDM爱尔兰1−/−小鼠不能产生或分泌衣康酸(图3A)在用LPS和IFN-γ刺激后,我们得出结论,Irg1是在这些条件下进行衣康酸合成的唯一酶。LPS激活的WT巨噬细胞显示,由于天冬氨酸-精氨琥珀酸分流术的活性,富马酸和苹果酸的浓度增加(Jha等人,2015年)以及琥珀酸的积累,可能是由于衣康酸的抑制。为了证实这一点,我们对爱尔兰1−/−BMDM和观察到的明显变化表明Sdh活性改变。缺乏Irg1表达导致琥珀酸积累消失,而富马酸和苹果酸浓度仍在增加(图3B). 因此,在缺乏内源性衣康酸的情况下,Sdh保持活性,并将琥珀酸氧化为富马酸,富马酸迅速转化为苹果酸(图3C).
图3。内源性衣康酸控制TCA循环重塑和琥珀酸水平。
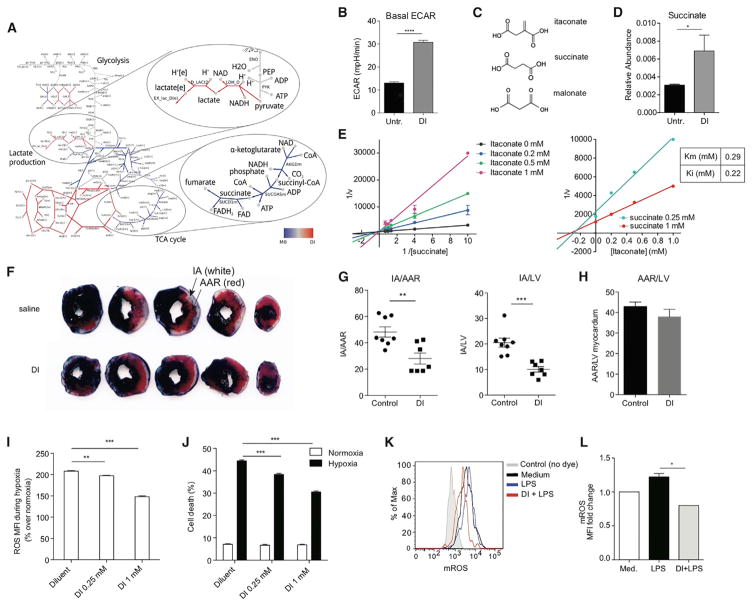
(A) 野生型和细胞内衣康酸盐分泌和细胞内的相对表达爱尔兰1−/−用LPS+IFN-γ激活后的BMDM。
(B) 琥珀酸、富马酸和苹果酸在(A)细胞提取物中的相对表达。
(C) 显示衣康酸如何调节LPS激活的巨噬细胞中TCA流量的方案。
(D) 通过休息(中等)和LPS激活的BMDM(24小时)从WT和爱尔兰1−/−老鼠。
数据显示为平均值±SEM(A和B,n=3;D,n=2总技术重复次数13-55)。p值采用双尾Student t检验计算*p<0.05***p<0.001****p<0.0001。另请参见图S3.
除了在TCA循环中的作用外,Sdh也是线粒体电子传输系统的一部分(作为复合物II)。在LPS激活的巨噬细胞中,线粒体呼吸显著降低(Kelly和O'Neill,2015年). 因此,我们通过测量耗氧率(OCR)来测试衣康酸介导的Sdh抑制是否影响线粒体功能。值得注意的是,与WT电池相反,红外线1−/−BMDM在激活后24小时显示OCR增加(图3D)这表明,通过抑制Sdh,内源性衣康酸调节线粒体呼吸,正如最近提出的外源性衣康酸酯(Németh等人,2016年).
内源性衣康酸盐调节巨噬细胞炎症反应
接下来,我们评估了在缺乏内源性衣康酸的情况下巨噬细胞的活化。我们使用RNA-seq分析LPS-激活野生型(WT)和爱尔兰1−/−BMDM。值得注意的是爱尔兰1−/−细胞与衣康酸处理的WT细胞基本呈负相关:基因在爱尔兰1−/−衣康酸处理的WT BMDM中细胞下调(图4A)提供了额外的证据,证明内源性衣康酸的功能类似于外源性添加的DI。与这些观察结果一致,LPS激活爱尔兰1−/−与WT细胞相比,BMDM产生更多的IL-12、NO和IL-6,但TNF-α的数量相似(图4B和4C).爱尔兰1−/−在刺激NLPR3的条件下,BMDM还维持了成熟IL-1β和IL-18的高表达(图4D). 与已知的HIF-1α促进IL-1β的作用一致(Tannahill等人,2013年),我们观察到HIF-1αmRNA和蛋白水平在爱尔兰1−/−DI-治疗的BMDM中HIF-1α表达的抑制(图4E–4G). 因此,HIF-1α和IL-1β生成的变化与琥珀酸盐被Sdh氧化的效率相关。这些观察结果增加了以下可能性:HIF-1α-IL-1β轴与活化巨噬细胞中电子传递链的效率和方向性有关,而不是通过琥珀酸积累直接传递信号。
图4。LPS激活的Irg1−/−缺乏衣康酸的巨噬细胞表现出增强的炎症反应。
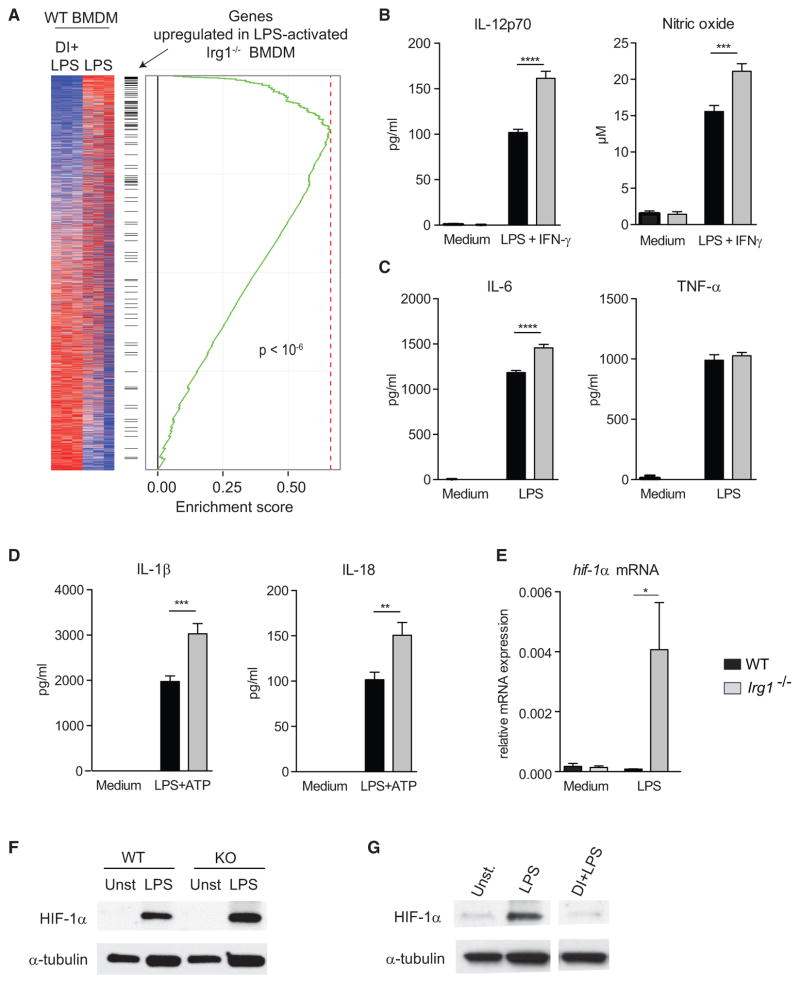
(A) 激活的转录签名爱尔兰1−/−和DI-处理激活的WT-BMDM呈负相关。
(B) WT和WT上清液中的IL-12和NO水平红外线1−/−用LPS+IFN-γ刺激BMDM(24小时)。
(C) LPS-活化(24小时)WT和爱尔兰1−/−BMDM公司。
(D) WT和WT的上清液中成熟的IL-1β和IL-18水平爱尔兰1−/−用LPS(4小时)和ATP刺激BMDM。
(E) 相对hif1a病毒LPS激活的WT和爱尔兰1−/−BMDM(4小时)。
(F) WT和WT裂解物中HIF-1α的Western blot分析爱尔兰1−/−BMDM激活,如(E)所示;α-微管蛋白作为负荷对照。所示斑点是两个独立实验的代表。
(G) Western blot分析未经处理或经DI-处理并用LPS激活的WT-BMDM裂解液中的HIF-1α(24小时)。所示斑点是两个独立实验的代表。
(B)–(E)中的数据显示为平均值±SEM(n=3/组,每个重复2-3次)。p值采用双尾Student t检验进行计算*p<0.05**p<0.01***p<0.001****p<0.0001。
总之,我们的工作将衣康酸确定为转录后机制的一部分,该机制通过抑制Sdh和调节琥珀酸水平来控制TCA循环重塑和巨噬细胞激活。我们的结果扩展了衣康酸的生理作用,使其超出了直接的抗菌作用,包括调节TLR介导的炎症细胞因子的产生,并提供了一种生理调节机制来控制电子传递链流量、琥珀酸水平、ROS的产生和组织炎症。
实验程序
老鼠
爱尔兰1−/−(MGI:103206)小鼠是在华盛顿大学接受胚胎干细胞(ESCs)(Irg1)后产生的tm1a(KOMP)Wtsi公司)来自敲除小鼠项目库(KOMP,加利福尼亚大学戴维斯分校),包含外显子3和4之间的插入盒。该盒阻止下游外显子4和5的转录和成熟蛋白的产生。爱尔兰1−/−将C57BL/6N ESCs微量注射到(Cg)-提尔c-2J/J白化受体雌性C57BL/6小鼠囊胚。选择黑毛嵌合小鼠,将其繁殖为野生型C57BL/6N小鼠。纯合子爱尔兰1−/−杂合动物杂交产生小鼠,并经PCR证实。爱尔兰1−/−小鼠有生育能力,表现出正常的孟德尔频率,并使用了来自两性的BMDM。
使用来自Charles River Laboratories的C57BL/6N WT小鼠作为年龄匹配的对照。根据华盛顿大学动物研究委员会批准的联邦和大学指南和协议,小鼠在华盛顿大学特定的无病条件下饲养。
巨噬细胞的分化和活化
BMDM由6-8周龄小鼠制备,如前所述(Jha等人,2015年)10岁时播种5在RMPI-1640培养基(ThermoFisher)中加入10%FBS、2 mM L-谷氨酰胺(Thermo Fisher Scientific)和100 U/mL青霉素-链霉素(Thermo-Fisher科学),96周组织培养板中的细胞/孔。用DI(0.25 mM,12小时;Sigma)处理或不处理细胞,并用LPS激活(大肠杆菌0111:B4;100纳克/毫升;西格玛)±干扰素γ(50 ng/mL;肽基)。对于炎症小体的激活,在添加ATP(3 mM,45 min;Sigma)、尼葛霉素(5 mM;1 hr;Sigma-)或尿酸单钠晶体(250μg/mL;4 hr;InvivoGen)之前,用LPS(100 ng/mL,4 hr)刺激细胞。
流式细胞术
用抗CD16/32(克隆93,Biolegend)培养细胞,对F4/80(克隆BM8,eBioscience)和CD11b(克隆M1/70;BD PharMinen)进行表面染色,并用LIVE/DEAD染料(Invitrogen)进行染色,然后用抗NOS2(克隆C-11;Santa Cruz Biotechnology)和FITC-结合IgG(克隆A85-1;BD PHARMinen)对细胞进行细胞内iNOS染色使用BD细胞修复/细胞周期试剂盒(BD Biosciences)。在Canto II流式细胞仪(BD Biosciences)上采集细胞,并使用FlowJo v.9.5.2软件(Tree Star)分析数据。
细胞因子和一氧化氮的定量
使用用于IL-1β/IL-1F2、TNF-α和IL-6的DuoSet试剂盒(所有研发系统)、IL-12 ELISA MAX Deluxe试剂盒(BioLegend)和IL-18 ELISA试剂盒(Medical&Biological Laboratories)对细胞因子进行定量。使用Griess试剂系统(Promega)检测一氧化氮。
西方印迹法
细胞在RIPA裂解缓冲液(Santa Cruz)中进行裂解,并在还原样品缓冲液中进行热变性(Thermo Fisher Scientific)。蛋白质在4%–20%聚丙烯酰胺梯度凝胶(BioRad)上分离并转移到PVDF膜上。用5%脱脂牛奶阻断非特异性结合,用IL-1β(1:1000;12507S,细胞信号)、Nlrp3(1:500;NBP2-12446,Novus)、HIF-1α(1:500,NB100-449,Novus)、α-微管蛋白(1:2000;2125S,细胞信号)和ASC(1:1000,sc-22514-R,Santa Cruz)的一级抗体探测细胞膜,然后用抗兔HRP(1:10000;sc-2030,Santa Cruz)和Clarity western ECL底物(Bio-Rad)培养。
Sdh活性测定
Sdh从BV2巨噬细胞系中纯化,并根据制造商的协议,使用复合II酶活性微孔板检测试剂盒(Abcam)在衣康酸存在下测定其活性。将底物在活性缓冲液中稀释,并在添加活性溶液前5分钟添加到磷脂混合物中。对于动力学分析,如活性缓冲液中所示,使用六水合琥珀酸钠。
GC-MS分析代谢产物
从每个样品中等量的细胞中提取细胞代谢物,并按所述用GC-MS进行分析(Vincent等人,2015年). 简单地说,用800μl 80%甲醇抑制细胞内代谢。为了分析分泌的代谢物,将20μl上清液添加到800μl 80%甲醇中。D-肉豆蔻酸(750纳克/样品)用作内标。用真空离心机干燥提取物,并将颗粒重新悬浮在含有10 mg/mL甲氧基胺盐酸盐的30μl吡啶中,然后使用N衍生-(叔丁基-丁基二甲基硅基)-N-甲基三氟乙酰胺。代谢物丰度表示为相对于内标物。
心肌缺血再灌注模型
体内缺血再灌注建模如下所述(Ma等人,2012年). 将8至10周大的小鼠麻醉,并进行可逆的左前降支结扎的开胸手术30分钟,随后再灌注2小时。在缺血前和缺血期间10分钟静脉输注盐水或DI(4mg/kg/min)。通过主动脉原位逆行注射心脏停搏液,然后再注射Evans Blue(LAD再次闭合后)。然后将左心室切成五片,在37°C(30分钟)的TTC(氯化三苯基四氮唑)中培养,并用ImageJ(NIH)分析图像。
新生大鼠心肌细胞缺氧模型的建立
按照描述分离和培养新生大鼠心肌细胞(Ma等人,2012年). 细胞在氧气控制柜(Coy Laboratories)中缺氧,氧气控制柜安装在培养箱内,配有氧气控制器和传感器,用于连续监测氧气水平。95%氮气和5%CO的混合物2用于制造缺氧,监测舱内氧气水平并保持在<1%。用哺乳动物细胞(Invitrogen)活细胞毒性活性试剂盒评估细胞死亡,用荧光羧基-H2DCFDA流式细胞术检测ROS(在10μmol/L中孵育30分钟后)(Ma等人,2012年).
mROS测量
用DI(0.25 mM,12小时)或载体处理细胞,在37°C下在添加0.1%BSA的HBSS中加载5μM MitoSOX(Invitrogen)30分钟,并用温培养基冲洗,然后用LPS刺激1小时(3小时),并在Canto II流式细胞仪(Becton Dickinson)上进行分析。
统计分析
在GraphPad Prism 6软件中使用每个实验的统计测试进行统计分析。
致谢
美国国立卫生研究院(NIH)拨款R01 AI104972(M.S.D.)支持这项研究。S.N.得到了DFG奖学金的支持。这个红外线1−/−靶向等位基因和ESCs是由小鼠生物学计划创造的(https://www.mousebiology.org/)加州大学戴维斯分校。ESC由trans-NIH敲除鼠标项目(KOMP)生成,并从KOMP知识库获得(https://www.komp.org/). NIH向Regeneron Inc.的Velocigene(U01HG004085)和CSD Consortium(U01HG004080)拨款,资助KOMP计划中8500多个基因的基因靶向载体和ESC的生成,并由加州大学戴维斯分校和CHORI的KOMP知识库存档和分发(U42RR024244)。我们感谢麦吉尔大学GCRC代谢组学核心设施提供的技术援助。E.E.V.和R.G.J.得到了加拿大卫生研究院的资助(CIHR MOP-142259 to R.G.J)。A.D.得到了NIH(HL107594)和退伍军人事务部(1I01BX001969)的资助。A.S.得到了俄罗斯联邦政府(074-U01)的资助。
脚注
作者贡献
V.L.和M.N.A.构思并设计了这项研究。V.L.进行了许多免疫学实验、流式细胞术、海马实验和数据分析。A.S.对RNA-seq数据进行了计算分析,并进行了通量平衡分析。M.B.对巨噬细胞进行了western blots、sdh活性分析和mROS测量。E.L.进行了基因组学分析并帮助进行了实验。S.N.和M.S.D.生成了爱尔兰1−/−老鼠。E.E.V.、T.G.和R.G.J.进行代谢组学分析。提供L.C.-B鼠伤寒沙门菌文化。C.J.W.、X.M.和V.L.在A.D.S.C.-C.H.的监督下进行了缺血再灌注模型实验和数据分析,帮助进行了海马实验。E.J.D、S.K.、G.J.R.和M.S.D.对手稿的研究设计和批判性编辑做出了贡献。V.L.和M.N.A.撰写了手稿的初稿。
参考文献
-
阿克曼WW,波特VR。与化疗有关的酶抑制。1949年《Proc Soc Exp Biol Med.》;72:1–9. doi:10.3181/00379727-72-17313。[内政部] [公共医学] [谷歌学者]
-
Bauernfeind F、Bartok E、Rieger A、Franchi L、Núñez G、Hornung V。前沿:活性氧抑制剂阻断NLRP3炎症小体的启动,但不能激活。免疫学杂志。2011;187:613–617. doi:10.4049/jimmunol.1100613。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Becker SA,Palsson BO。上下文特定的代谢网络与实验一致。公共科学图书馆计算生物学。2008;4:e1000082。doi:10.1371/journal.pcbi.1000082。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Bennett BD、Kimball EH、Gao M、Osterhout R、Van Dien SJ、Rabinowitz JD。大肠杆菌中的绝对代谢物浓度和隐含酶活性位点占有率。自然化学生物。2009;5:593–599. doi:10.1038/nchembio.186。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Bordbar A、Mo ML、Nakayasu ES、Schrimpe-Rutledge AC、Kim YM、Metz TO、Jones MB、Frank BC、Smith RD、Peterson SN等。模型驱动的多组分数据分析阐明了巨噬细胞活化的代谢免疫调节剂。分子系统生物学。2012;8:558. doi:10.1038/msb2012.21。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Broz P、Newton K、Lamkanfi M、Mariathasan S、Dixit VM、Monack DM。炎症小体受体NLRP3和NLRC4在宿主防御沙门氏菌中的冗余作用。《实验医学杂志》2010;207:1745–1755. doi:10.1084/jem.20100257。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Cho H,Proll SC,Szretter KJ,Katze MG,Gale M,Jr,Diamond MS。神经元亚型的不同先天免疫反应程序决定了阳性标记RNA病毒对大脑感染的敏感性。《国家医学》2013;19:458–464. doi:10.1038/nm.3108。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Chouchani ET、Pell VR、Gaude E、AksentijevićD、Sundier SY、Robb EL、Logan A、Nadtochiy SM、Ord EN、Smith AC等。琥珀酸的缺血积累通过线粒体ROS控制再灌注损伤。自然。2014;515:431–435. doi:10.1038/nature13909。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Degrandi D,Hoffmann R,Beuter-Gunia C,Pfeffer K。促炎细胞因子诱导的IRG1蛋白与线粒体相关。干扰素细胞因子研究杂志,2009;29:55–67. doi:10.1089/jir.2008.013。[内政部] [公共医学] [谷歌学者]
-
Everts B、Amiel E、Huang SC、Smith AM、Chang CH、Lam WY、Redmann V、Freitas TC、Blagih J、van der Windt GJ等。通过激酶TBK1-IKKεTLR驱动的早期糖酵解重编程支持树突状细胞激活的合成代谢需求。自然免疫学。2014;15:323–332. doi:10.1038/ni.2833。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Huang SC、Everts B、Ivanova Y、O'Sullivan D、Nascimento M、Smith AM、Beatty W、Love-Gregory L、Lam WY、O’Neill CM等。细胞内溶酶体脂解对巨噬细胞的替代激活至关重要。自然免疫学。2014;15:846–855。doi:10.1038/ni.2956。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
宾夕法尼亚州詹森,日本帕宾。代谢网络模型和表达数据的功能集成,无需任意阈值。生物信息学。2011;27:541–547. doi:10.1093/bioinformatics/btq702。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Jha AK、Huang SC、Sergushishev A、Lampropoulou V、Ivanova Y、Loginicheva E、Chmielewski K、Stewart KM、Ashall J、Everts B等。并行代谢和转录数据的网络整合揭示了调节巨噬细胞极化的代谢模块。豁免权。2015;42:419–430. doi:10.1016/j.immuni.2015.02.005。[内政部] [公共医学] [谷歌学者]
-
Kelly B,O'Neill LA。先天免疫中巨噬细胞和树突状细胞的代谢重编程。细胞研究2015;25:771–784. doi:10.1038/cr.2015.68。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Ma X,Liu H,Foyil SR,Godar RJ,Weinheimer CJ,Hill JA,Diwan A.自噬体清除受损导致心肌细胞在缺血/再灌注损伤中死亡。循环。2012;125:3170–3181. doi:10.1161/CIRCULATIONAHA.111.041814。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Michelucci A、Cordes T、Ghelfi J、Pailot A、Reiling N、Goldmann O、Binz T、Wegner A、Tallam A、Rausell A等。免疫应答基因1蛋白通过催化衣康酸的产生将代谢与免疫联系起来。美国国家科学院院刊2013;110:7820–7825. doi:10.1073/pnas.1218599110。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Moon JS、Hisata S、Park MA、DeNicola GM、Ryter SW、Nakahira K、Choi AM.mTORC1-诱导的HK1-依赖性糖酵解调节NLRP3炎症激活。细胞报告2015;12:102–115。doi:10.1016/j.celrep.2015.05.046。[内政部] [PMC免费文章] [公共医学] [谷歌学者] [已缩回]
-
Naujoks J、Tabeling C、Dill BD、Hoffmann C、Brown AS、Kunze M、Kempa S、Peter A、Mollenkopf HJ、Dorhoi A等。IFNs修改含军团菌液泡的蛋白质组并通过IRG1-衍生衣康酸限制感染。《公共科学图书馆·病理学》。2016;12:e1005408。doi:10.1371/journal.ppat.1005408。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Németh B、Doczi J、Csete D、Kacso G、Ravasz D、Adams D、Kiss G、Nagy AM、Horvath G、Tretter L等。LPS诱导小鼠巨噬细胞系细胞中Irg1表达产生的衣康酸对线粒体底物水平磷酸化的消除。FASEB J.2016;30:286–300. doi:10.1096/fj.15-279398。[内政部] [公共医学] [谷歌学者]
-
Strelko CL、Lu W、Dufort FJ、Seyfried TN、Chiles TC、Rabinowitz JD、Roberts MF。衣康酸是巨噬细胞激活期间诱导的哺乳动物代谢物。美国化学学会杂志2011;133:16386–16389. doi:10.1021/ja2070889。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Tannahill GM、Curtis AM、Adamik J、Palsson-McDermott EM、McGettrick AF、Goel G、Frezza C、Bernard NJ、Kelly B、Foley NH等。琥珀酸是一种炎症信号,通过HIF-1α诱导IL-1β。自然。2013;496:238–242. doi:10.1038/nature11986。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Vincent EE、Sergushichev A、Griss T、Gingras MC、Samborska B、Ntimbane T、Coelho PP、Blagih J、Raissi TC、Choinière L等。线粒体磷酸丙酮酸羧激酶调节代谢适应并促进葡萄糖非依赖性肿瘤生长。摩尔细胞。2015;60:195–207. doi:10.1016/j.molcel.2015.08.013。[内政部] [公共医学] [谷歌学者]
关联数据
本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。