摘要 淀粉样β(Aβ)诱导的氧化应激是阿尔茨海默病的主要病理特征。 氰胺是一种天然类黄酮化合物,对氧化损伤介导的变性具有神经保护作用。 然而,其分子机制尚不清楚。 在这里,我们研究了花青素预处理对Aβ诱导的PC12细胞神经毒性的影响,并探讨了潜在的机制。 氰胺预处理显著降低了Aβ诱导的PC12细胞死亡率和形态变化。 从机制上讲,花青素通过恢复线粒体膜电位有效阻止Aβ诱导的细胞凋亡 通过 Bcl-2蛋白表达上调。 此外,花青素通过阻断活性氧物种和超氧物积累,显著保护PC12细胞免受Aβ诱导的DNA损伤。 这些结果提供了证据表明,氰化物通过阻止活性氧化物介导的氧化损伤来抑制Aβ诱导的细胞毒性,而活性氧化物反过来又抑制线粒体凋亡。 我们的研究证明了氰化物在预防氧化应激介导的Aβ神经毒性方面的治疗潜力。
关键词: 神经再生 , 氰化物 , 淀粉样蛋白-β , 氧化损伤 , 活性氧化物种类 , 细胞凋亡 , 神经再生
介绍 阿尔茨海默病(AD)是一种与年龄相关的慢性进行性神经退行性疾病。 淀粉样蛋白β(Aβ)的沉积会导致老年斑块,使神经元受到氧化应激并破坏细胞内连接,导致神经元死亡。 这被认为是AD发病的主要机制(Selkoe,1998;Huang and Jiang,2009;Eckert et al.,2010;Pagani and Eckert,2011)。 活性氧(ROS)是一种小的活性分子,在引发人类疾病中起着重要作用。 β介导的氧化应激可攻击蛋白质、膜脂和DNA,诱导凋亡细胞死亡,并最终导致神经网络功能障碍(Acquaviva等人,2003年)。 因此,阻断Aβ诱导的氧化应激被认为是对抗AD的有效策略,有效的抗氧化剂在神经退行性疾病的治疗中显示出潜力。
氰化物是一种存在于红色浆果中的色素,属于类黄酮家族,因其抗氧化和生物特性而备受关注(Acquaviva等人,2003年;King等人,2013年)。 Acquaviva等人(2003年)报告称,氰化物对ROS介导的氧化损伤具有显著的保护潜力。 进一步的证据还表明,氰化物类似物可以消除过量生产的活性氧(Chen等人,2003年;Guerra等人,2005年)。 氰胺还被确定为一种抵抗氧化应激的神经保护成分(Kim等人,2005年)。 然而,保护效率和机制仍有待详细评估。 Ye等人(2010年)报告称,紫甘薯花色苷可以通过抑制氧化应激抑制Aβ诱导的PC12细胞的细胞毒性。 因此,在本研究中,我们使用PC12细胞系作为 在体外 神经元氧化损伤评估模型(Paavlica等人,2005),以研究氰化物保护PC12细胞免受Aβ诱导的神经毒性和凋亡的潜力。
材料和方法 细胞培养和药物治疗 PC12细胞(ATCC,MD,USA)保存在含有10%胎牛血清、100 U/mL青霉素和50 U/mL链霉素的高糖Dulbecco改良Eagle’s培养基中,浓度为5%CO 2 37°C时。 将细胞预先培养在96周的平板(1×10)中 4 每孔细胞数)24小时。 Aβ 25–35 (西格玛,加利福尼亚州旧金山,美国)在治疗前溶解在磷酸盐缓冲盐水(PBS)中,并在37°C下培养3天。 进行了一项剂量研究,将PC12细胞暴露于氰化物(5–80μM)24小时,然后再暴露于Aβ 25–35 (5、10、20、40和80µM)持续24小时。 为了进行保护实验,PC12细胞用氰化物(20、40、80µM;Sigma)预培养24小时,然后用Aβ 25–35 (10µM)持续24小时。
MTT分析 使用MTT分析细胞活力(Fan等人,2014)。 暴露于花青素和Aβ后 25–35 ,将20µL MTT溶液(Sigma;5 mg/mL)添加到每个孔中,在37°C下持续5小时。 取出培养基,向每个孔中添加100µL DMSO,并将平板培养15分钟。 使用SpectraMax M3(Molecular Devices,Sunnyvale,CA,USA)在570nm处测量颜色强度。 假设对照细胞的吸光度为100%,结果以MTT还原百分比表示。 细胞在显微镜下观察(尼康Eclipse 80i;尼康,日本东京)。
TUNEL/DAPI双重染色 如前所述,使用TUNEL-DAPI双染色试剂盒(Beyotime,中国北京)测量细胞凋亡(Fan等人,2014)。 将培养在培养箱载玻片中的细胞固定,用0.1%Triton X-100在PBS中渗透2分钟,用TUNEL反应混合物和末端脱氧核苷酸转移酶处理,用PBS洗涤,并在显微镜下观察(尼康)。
线粒体膜电位(Δψm)的测量 如前所述,使用JC-1染料评估Δψm(Fan等人,2014)。 简而言之,PC12细胞与80µM氰化物孵育1小时,与10µM Aβ共同孵育 25–35 2小时,然后用JC-1探针(10μg/mL)处理10分钟。 用PBS清洗细胞,并用荧光显微镜(200倍放大)观察。
半胱氨酸天冬氨酸蛋白酶-3活性 氰化物和Aβ之后 25–35 处理后,收集PC12细胞并在缓冲液中溶解,然后离心。 如前所述,测量蛋白质浓度,并检测caspase-3活性(Fan等人,2014)。 简单地说,将caspase-3底物Ac-Asp-Glu-Val-Asp-AMC(Ac-DEVD-AMC;Cell Signaling Technology,Boston,USA)和100µg总蛋白样品添加到96-well板的每个孔中。 将平板在37°C的黑暗中培养2小时,并分别在380 nm和440 nm的激发和发射波长下测量caspase-3活性。
活性氧和超氧物的检测 使用荧光法(二氯二氢荧光素双乙酸酯测定法(DCFH-DA;Beyotime)测定活性氧的相对水平。 由于自由基生成是一个早期事件,如之前报告的那样,ROS和超氧物在2小时内检测到(Wang等人,2015;Fu等人,2016)。 简单地说,细胞接种于1×10 4 细胞/孔置于96 well板中,然后用或不用20–80µM氰化物孵育1小时,并用10µM aβ共同孵育 25–35 持续2小时。 向细胞中添加DCFH-DA(10µM),并分别在488 nm和525 nm的激发和发射波长下监测ROS生成。 超氧化物由线粒体靶向荧光染料MitoSOX(Beyotime)检测。
蛋白质印迹分析 如前所述,通过western blot分析评估蛋白质表达(Wang等人,2015;Fu等人,2016)。 简单地说,收集细胞并进行裂解,使用BCA分析试剂盒对蛋白质进行定量。 将蛋白质与缓冲液混合,煮沸,并使用40µg蛋白质进行SDS-聚丙烯酰胺凝胶电泳(SDS-PAGE)。 将蛋白质转移到硝化纤维素膜上,然后用5%的非脂肪乳(Sigma)封闭2小时,并在4°C下与一级抗体(1:2000)孵育12小时。 主要抗体为caspase-3、Bcl-xl、Bcl-2、Bas、Bad、Ser15-p53、total-p53、Ser139-histone和β-actin(均来自细胞信号技术)。 然后将膜与二级抗体(1:2000)(细胞信号技术)在37°C下孵育2小时。 蛋白质在X射线胶片上可视化,并使用Quantity-One软件进行量化(Bio-Rad,Hercules,CA,USA)。 靶蛋白表达归一化为β-肌动蛋白(参考对照)。
统计分析 使用SPSS 17.0软件(SPSS Inc.,美国伊利诺伊州芝加哥)进行统计分析。 数据以平均值±标准差表示。通过双尾Student’s分析两组之间的差异 t吨 -测试。 通过单因素方差分析和多重比较分析三组或更多组之间的差异。 对 <0.05被认为具有统计学意义。
结果 氰胺保护PC12细胞免受Aβ诱导的细胞毒性 细胞生长首先在存在氰化物和Aβ的情况下进行评估。 MTT分析表明,在浓度为5–80µM的氰化物处理后,PC12细胞活力没有变化( 图1A ). 单独暴露于Aβ(5–80μM)会以浓度依赖的方式降低细胞活力,80μM时毒性最大( 图1B ). 因此,我们选择了10µM aβ的浓度进行后续实验,该浓度可诱导约40%的细胞死亡。 暴露于10µM Aβ后,细胞活力为60.2%( 图1C ). 然而,花青素预处理有效地阻止了Aβ诱导的PC12细胞活性下降。 在40和80µM时,氰化物将细胞存活率从60.2%分别提高到79.2%和94.4%。 结果表明,花青素预处理能有效阻断Aβ诱导的细胞毒性。 此外,与对照细胞相比,暴露于Aβ的细胞出现凋亡小体,数量较少。 正如预期的那样,花青素预处理显著改善了Aβ诱导的形态变化( 图1D ). 未观察到凋亡小体,且氰化物预处理组的细胞数量明显大于单独暴露于Aβ的组。 这些细胞形态和数量的变化进一步验证了氰化物的保护潜力( 图1D ).
图1。
氰胺(Cya)抑制淀粉样β(Aβ)诱导的PC12细胞的细胞毒性。
(A–C)Cya对PC12细胞活力的影响(MTT法)。 将接种在96-well板中的PC12细胞用Cya(5-80μM)处理48小时(a),用aβ(5-80µM)处理24小时(B),或两者都用(Cya 5–80µM+aβ10μM;C)。 Cya保护细胞免受Aβ诱导的细胞毒性。 MTT法测定细胞活力。 单独Aβ治疗(5-80μM)剂量依赖性降低细胞活力( 对 <0.05),而Cya剂量依赖性地增加了20–80μM时的细胞活力( 对 < 0.05). 采用单向方差分析进行多重比较。 数据以四个独立实验的平均值±标准差表示。 (D) 含有Cya和Aβ的PC12细胞的形态学变化。 箭头,凋亡小体。
氰胺阻断Aβ诱导的PC12细胞凋亡 首次用TUNEL和DAPI染色检测了氰胺对Aβ诱导的细胞凋亡的预防作用。 凋亡细胞产生许多3′-OH末端,当受到TUNEL反应时,这些末端发出绿色荧光。 因此,绿色荧光代表凋亡细胞。 单独暴露于Aβ的PC12细胞表现出显著的凋亡变化,如染色质凝聚和核碎裂( 图2A ). 此外,还检测了caspase-3的裂解和活性。 Aβ处理后Caspase-3活性和裂解显著升高( 图2B , C类 ; 对 < 0.05). 然而,花青素预处理保护PC12细胞免受Aβ诱导的凋亡,表现为荧光强度降低,并显著减弱caspase-3活性和分裂( 对 < 0.05). 总的来说,结果证实了花青素预处理通过抑制Aβ诱导的涉及caspase-3激活的细胞凋亡,显著抑制了Aβ诱导细胞毒性。
图2。
氰胺(Cya)可阻止淀粉样βAβ诱导的PC12细胞凋亡。
(A) TUNEL/DAPI双重染色。 细胞暴露于80µM Cya 24小时和/或10µM Aβ24小时,TUNEL/DAPI双重染色检测细胞凋亡。 箭头表示TUNEL阳性细胞。 (B) 荧光法测定Cya阻断Aβ诱导的caspase-3激活* 对 < 0.05, 与 .控制# 对 < 0.05, 与 Aβ治疗。 采用单向方差分析进行多重比较。 (C) 半胱氨酸天冬氨酸蛋白酶-3表达(蛋白质印迹)。 检测了全长caspase-3(35kDa)和caspase-3的裂解分数(CF)。 使用Quantity-One软件测量蛋白质表达,并根据β-肌动蛋白标准化所有测量值。 所有实验都进行了三次。
氰胺通过调节Bcl-2抑制Aβ诱导的Δψm损失 鉴于线粒体在药物诱导凋亡中的重要性,采用JC-1探针检测了Δψm。 JC-1探针能穿过细胞线粒体并显示出明显的红色荧光。 但是,作为对凋亡刺激的响应,探针可以转移到细胞质中并显示绿色荧光。 因此,荧光变化可以间接反映Δψm的损失。 单独Aβ暴露后,PC12细胞的Δψm显著降低,表现为增强的绿色荧光( 图3A ). 仅用氰化物处理的细胞没有引起Δψm的变化。 用花青素预处理细胞可完全逆转Aβ诱导的Δψm耗竭。 结果表明,花青素通过调节Δψm来阻断Aβ诱导的线粒体功能障碍。 Bcl-2家族在调节Δψm和启动线粒体介导的凋亡中起着关键作用。 用western blot检测Bcl-2家族成员(包括促凋亡和促生存成员)( 图3B ). Aβ治疗显著上调Bax和Bad的表达,但下调Bcl-XL和Bcl-2的表达,表明Aβ通过调节Bcl-2家族诱导PC12细胞凋亡。 然而,花青素预处理显著阻断了Aβ诱导的Bcl-2家族失衡。 总之,这些结果表明,矢车菊素通过调节Bcl-2家族来抑制Aβ诱导的细胞凋亡和Δψm的损失。
图3。
氰胺(Cya)通过调节Bcl-2家族蛋白抑制淀粉样β(Aβ)诱导的线粒体膜电位(Δψm)损失。
(A) Cya抑制Aβ介导的Δψm耗竭。 JC-1探头用于检测Δψm。 红色,线粒体荧光; 绿色,细胞质荧光。 荧光从红色变为绿色表明Δψm的变化。 (B) Cya阻止了Aβ诱导的Bcl-2家族蛋白表达下降(western blot分析)。 所有实验都进行了三次。
氰胺阻断活性氧介导的Aβ暴露细胞DNA损伤 采用DCFH-DA法测定ROS累积。 用10µM Aβ治疗1小时后,细胞内ROS显著增加至对照组的226.3%( 图4 ). 然而,用花青素(20、40和80µM)预处理导致暴露于aβ的细胞的ROS显著下降(分别下降到198.3%、153.8%和118.1%),且呈剂量依赖性。 此外,还通过线粒体靶向红色染料MitoSOX检测活细胞中的超氧化物。 Aβ处理后,PC12细胞线粒体中出现显著的超氧物生成,但氰化物预处理显著阻止了这一现象( 图4B )红色荧光微弱就证明了这一点。 这些结果表明,花青素保护PC12细胞免受Aβ诱导的细胞毒性 通过 抑制活性氧和超氧物。
图4。
氰胺(Cya)通过抑制活性氧(ROS)的积累来防止淀粉样β(Aβ)诱导的DNA损伤。
(A) Cya阻止了Aβ诱导的PC12细胞中ROS的积累。 用Cya(20–80µM)预处理细胞1小时,然后在有或无10µM Aβ的情况下培养2小时。 使用DCFH-DA染料监测ROS累积。 20–80µM剂量依赖性的Cya治疗可降低Aβ诱导的活性氧升高( 对 < 0.05). 采用单向方差分析进行多重比较。 (B) Cya抑制Aβ诱导的PC12细胞产生超氧物。 使用线粒体靶向染料MitoSOX观察超氧化物。 DAPI染色细胞核。 这里显示的所有图像都是三个独立实验的代表。 红色荧光,线粒体超氧化物。 (C) Cya对Aβ诱导的DNA损伤的保护(western blot分析)。 实验进行了三次。
活性氧的积累会破坏DNA,甚至导致凋亡细胞死亡。 因此,我们检测了氰化物对Ser15-p53和Ser139-H2A磷酸化的影响,这两种都是DNA损伤标记物。 在暴露于Aβ的PC12细胞中,磷酸化的Ser139-H2A和Ser15-p53显著上调( 对 < 0.05). 正如预期的那样,用氰化物预处理完全抑制了Ser139-H2A和Ser15-p53的磷酸化,达到了在对照细胞中观察到的水平( 对 <0.05),表明氰化物预处理可防止Aβ诱导的DNA损伤。 总之,这些结果提供了证据,证明矢车菊素预处理通过抑制ROS的积累显著减轻了Aβ诱导的DNA损伤。
讨论 β诱导的氧化应激和氧化还原失调有助于神经退行性疾病的病因和病理学(Selkoe,1998;Huang和Jiang,2009;Eckert等人,2010;Pagani和Eckert,2011)。 氧化应激可能损伤神经细胞,被认为是神经毒性的主要原因,在人类神经退行性疾病的发生和发展中起着重要作用(Crispo等人,2010;Tapias等人,2014)。 活性氧很容易穿过细胞质膜,结合DNA,导致DNA功能紊乱(Gao等人,2013)。 抗氧化治疗有助于控制这些疾病(Arai等人,2007年)。 因此,在流行病学研究中,具有抗氧化活性的药物通常被接受为化学预防剂,以调节氧化应激介导的疾病的进展。 越来越多的证据表明,花青素是一种广泛使用的天然产物,具有新颖的抗氧化活性,可以直接清除自由基。 然而,其机制尚不清楚。
半胱氨酸蛋白酶是在启动细胞凋亡中起积极作用的半胱氨酸蛋白酶。 半胱氨酸天冬氨酸蛋白酶-3可以被切割并激活以响应细胞凋亡(Kong等人,2003;Zhang等人,2008)。 我们的结果表明,花青素阻止了Aβ诱导的PC12细胞激活,并降低了caspase-3的表达水平。 半胱氨酸天冬氨酸蛋白酶-3活性和表达的变化都证实了花青素具有抑制Aβ诱导的细胞凋亡的潜力。 活性氧是调节和诱导细胞凋亡的重要小分子。 ROS积累和细胞防御系统之间的平衡最终决定细胞的存活或死亡(Pabla等人,2008;Fan等人,2014)。 氰化氢可以通过减少ROS的产生来保护神经元细胞免受氧化损伤诱导的神经毒性的影响(Li等人,2013;Mokni等人,2013)。 在本研究中,Aβ显著增加PC12细胞中ROS的积累并激活DNA损伤。 然而,花青素有效地终止了Aβ诱导的ROS生成和DNA损伤。 研究结果表明,花青素具有清除自由基的潜力,从而防止PC12细胞受到ROS介导的氧化应激和凋亡。
抗氧化酶系统是内源性自由基清除剂(Mokni等人,2013)。 这些酶可以转移H 2 O(运行) 2 阻断体内过氧化脂质的过程,使过氧化脂质还原为无害的多元醇(Kalpana等人,2007)。 因此,我们推测,花青素的加入可能会影响内源性抗氧化酶系统,消除Aβ诱导的ROS,最终抑制Aβ诱导的PC12细胞凋亡,为开发治疗神经系统疾病的抗氧化药物提供了新的思路。
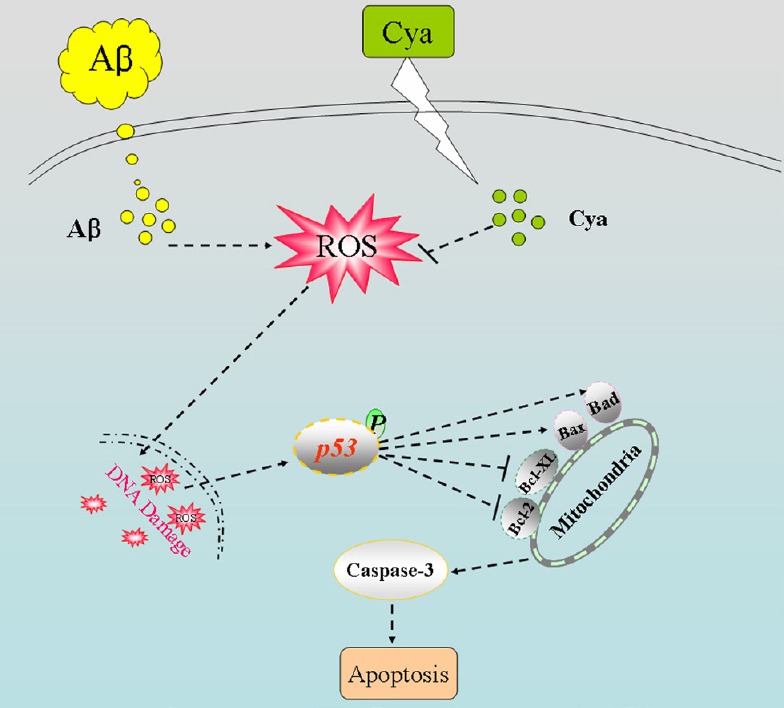
总之,我们评估了花青素对Aβ诱导的PC12细胞神经毒性的保护潜力,并探讨了其潜在机制。 我们的结果表明,花青素通过抑制ROS介导的氧化损伤和线粒体介导的凋亡来抑制Aβ诱导的细胞毒性( 图5 ).
图5。
说明潜在信号通路的方案。
淀粉样β(Aβ)直接引起活性氧(ROS)积累,引发DNA损伤,诱导PC12细胞凋亡。 然而,花青素通过调节Bcl-2家族蛋白的表达和恢复Δψm,抑制活性氧的积累,减轻DNA损伤,最终逆转Aβ诱导的PC12细胞凋亡。
脚注
基金: 本研究得到了中国山东省自然科学基金(No.ZR2014HM046)(致ZCZ)的资助 .
剽窃检查: 这篇论文在发表前用交叉检查法进行了两次筛选,以验证其原创性 .
同行评审: 这篇论文是由国际专家评审员进行的双盲严格评审 .
由Slone-Murphy J、Raye W、Li CH、Song LP、Zhao M编辑
工具书类
Acquaviva R、Russo A、Galvano F、Galvan O G、Barcellona ML、Li Volti G.氰化物和氰化物3-O-β-D-葡萄糖苷作为DNA裂解保护剂和抗氧化剂。 细胞生物毒性。 2003; 19:243–252. doi:10.1023/b:cbto.0000003974.27349.4e。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Arai M、Shibata Y、Pugdee K、Abiko Y、Ogata Y。活性氧(ROS)对MC3T3-E1细胞抗氧化系统和成骨分化的影响。 IUBMB寿命。 2007; 59:27–33. doi:10.1080/15216540601156188。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Chun OK,Kim DO,Lee CY。新鲜李子中主要多酚的超氧自由基清除活性。 农业食品化学杂志。 2003; 51:8067–8072. doi:10.1021/jf034740d。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Crispo JA,Piche M,Ansell DR,Eibl JK,Tai IT,Kumar A.没食子酸甲酯对H2O2诱导PC12细胞凋亡的保护作用。 生物化学与生物物理研究委员会。 2010; 393:773–778. doi:10.1016/j.bbrc.2010.02.079。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Eckert A,Schulz KL,Rhein V,Gotz J.体内线粒体淀粉样β和tau病理学的会聚。 摩尔神经生物学。 2010; 41:107–114. doi:10.1007/s12035-010-8109-5。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Fan C,Zheng W,Fu X,Li X,Wong YS,Chen T.通过硒代半胱氨酸(一种调节ROS介导信号的协同剂)增强阿霉素对人肝细胞癌的治疗效果的策略。 Oncotarget公司。 2014; 5:2853–2863. doi:10.18632/目标1854。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
傅XY,杨MF,曹MZ,李德伟,杨XY,孙JY。 姜黄素抑制PC12细胞氧化损伤诱导的神经毒性的策略,ROS介导的DNA损伤以及MAPK和AKT通路的作用。 摩尔神经生物学。 2016; 53:369–378. doi:10.1007/s12035-014-9021-1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
高S、陈涛、崔美美、梁毅、薛杰、王毅。 氰胺通过抑制ROS介导的DNA损伤和调节ERK和AKT通路,逆转顺铂诱导的HK-2近端肾小管细胞凋亡。 癌症快报。 2013; 333:36–46. doi:10.1016/j.canlet.2012.12.029。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Guerra MC、Galvano F、Bonsi L、Speroni E、Costa S、Renzulli C.氰胺-3-O-β-吡喃葡萄糖苷,一种天然自由基清除剂,可对抗黄曲霉毒素B1和赭曲霉毒素a在人类肝癌细胞系(Hep G2)和人类结肠腺癌细胞系(CaCo-2)Br J Nutr中诱导的细胞损伤。 2005; 94:211–220. doi:10.1079/bjn20051425。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
黄慧聪、蒋采发。 累积淀粉样β肽和高磷酸化tau蛋白:阿尔茨海默病中的关系和联系。 阿尔茨海默病杂志。 2009; 16:15–27. doi:10.3233/JAD-2009-0960。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Kalpana C、Sudheer AR、Rajasekharan KN、Menon副总裁。 姜黄素及其合成类似物在烟碱诱导毒性期间对组织脂质过氧化和抗氧化状态的比较作用。 新加坡医学杂志2007; 48:124–130. [ 公共医学 ] [ 谷歌学者 ]
Kim DO,Heo HJ,Kim YJ,Yang HS,Lee CY。甜樱桃酚及其对神经细胞的保护作用。 农业食品化学杂志。 2005; 53:9921–9927. doi:10.1021/jf0518599。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
国王AE、Southam KA、迪特曼J、维克斯JC。 紫杉醇抑制兴奋毒素诱导的caspase-3激活和轴突微管解体。 神经病理学会学报。 2013; 1:59. doi:10.1186/2051-5960-1-59。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
孔JM,Chia LS,Goh NK,Chia TF,Brouillard R.花色苷的分析和生物活性。 植物化学。 2003; 64:923–923. doi:10.1016/s0031-9422(03)00438-2。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Li S,Shen C,Guo W,Zhang X,Liu S,Liang F.木糖衍生物在帕金森病模型中的合成和神经保护作用。 Mar药物。 2013; 11:5159–5189. doi:10.3390/md11125159。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Mokni M、Hamlaoui S、Karkouch I、Amri M、Marzouki L、Limam F。白藜芦醇通过调节抗氧化酶活性,在缺血/再灌注损伤后提供心脏保护。 伊朗医药研究杂志2013; 12:867–875. [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Pabla N,Dong Z.顺铂肾毒性:机制和肾保护策略。 《肾脏国际》2008; 73:994–1007. doi:10.1038/sj.ki.5002786。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Pagani L,Eckert A.淀粉样β与线粒体的相互作用。国际老年痴呆症杂志。 2011年doi:10.40601/2011/925050。 doi:10.4061/2011/925050。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Pavlica S,Gebhardt R.鞣花酸和绿原酸对PC12细胞氧化应激的保护作用。 自由基研究2005; 39:1377–1390. doi:10.1080/09670260500197660。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Selkoe DJ。阿尔茨海默病中β-淀粉样前体蛋白和早老蛋白的细胞生物学。 细胞生物学趋势。 1998; 8:447–453. doi:10.1016/s0962-8924(98)01363-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
塔皮亚斯五世、坎农JR、格林纳米尔JT。 石榴汁加剧了帕金森氏病患者的氧化应激和黑质纹状体变性。 神经生物老化。 2014; 35:1162–1176. doi:10.1016/j.neurobiolaging.2013.10.077。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Wang K,Fu XY,Fu XT,Hou YJ,Fang J,Zhang S.DSePA拮抗高葡萄糖诱导的神经毒性:dna损伤介导的p53磷酸化、mapks和akt通路的证据。 摩尔神经生物学。 2015年doi:10.1007/s12035-015-9373-1。 DOI:10.1007/s12035-015-9373-1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
叶JL、孟晓杰、闫CL、王CB。 紫甘薯花色苷通过抑制氧化应激对β-淀粉样蛋白介导的PC-12细胞死亡的影响。 神经化学研究2010; 35:357–365. doi:10.1007/s11064-009-0063-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Zhang YB,Lu GW,Yang MF,Niu JZ,Sun BL.低氧预适应小鼠皮层Bcl-2和Caspase-3表达的变化。 《胜利学报》。 2008; 60:249–253. [ 公共医学 ] [ 谷歌学者 ]
神经再生研究的文章由以下人员提供 Wolters Kluwer——Medknow出版物