摘要
谷氨酰胺的利用促进了癌细胞的生长。我们提出了一个新的癌症代谢概念图,其中线粒体NADH和NADPH在存在功能失调的电子转移链的情况下,促进谷氨酰胺的还原羧基化。我们还讨论了为什么需要烟酰胺核苷酸转氢酶(NNT)体内用于通过还原羧化来利用谷氨酰胺。此外,由戊糖磷酸途径和癌症特异性丝氨酸糖酵解转移产生的NADPH似乎可以维持谷氨酰胺在氨基酸合成、脂质合成和活性氧猝灭中的利用。NAD的供应+前体物降低肿瘤侵袭性提示了实验方法来阐明NADH驱动的氧化还原网络在癌症中的作用。
事实
大多数癌细胞的持续生长和存活依赖于代谢重组,其特征是糖酵解通量增强,并通过还原羧化刺激谷氨酰胺的利用。
线粒体功能障碍(例如复合物I活性降低)促进了癌症代谢重组的发生。
北美+/NADH和NADP+/NADPH参与癌症代谢重组途径的几种反应。
控制癌细胞中NAD(H)代谢影响其在异种移植中的肿瘤形成能力。
开放式问题
在以不同侵袭性为特征的各种癌症细胞类型中,NADPH的定量通量(来自戊糖磷酸途径或丝氨酸合成、一碳五代代谢和甘氨酸裂解途径)最终在NEAA、GSH和脂质中是什么?
癌症代谢重组中NADH和NADPH代谢的线粒体/细胞溶质分区是什么?
癌细胞中的细胞质苹果酸脱氢酶/苹果酸酶途径是否将NADH转化为NADPH?
两种代谢适应被认为是癌细胞的特征:(1)通过糖酵解途径增加葡萄糖利用以产生乳酸1和(2)通过还原羧化增加谷氨酰胺的消耗以维持合成代谢过程。2虽然癌细胞的代谢特征可能会受到转化组织和相关癌基因的影响,三许多不同类型的癌症都有代谢重组的迹象。因此,葡萄糖类似物2-氟脱氧葡萄糖(FDG)目前被用于正电子发射断层扫描(PET)的诊断目的。4
几种致癌信号通路通常依赖于癌基因的激活,如ras和myc,在几种人类癌症中发现突变,5,6,7通过诱导线粒体功能障碍(通常导致复合物I活性降低)和促进糖酵解和谷氨酰胺利用来表达其转化活性。6,8,9,10,11,12,13,14,15,16,17
抑制OXPHOS线粒体途径,特别是复合物I,能够促进谷氨酰胺的利用。18癌症代谢重塑的两种途径都需要支持癌细胞增殖。事实上,线粒体谷氨酰胺酶(GLS2)催化谷氨酰胺水解为谷氨酸,而乳酸脱氢酶A(丙酮酸转化为乳酸的酶)的抑制都能抑制肿瘤细胞增殖。19,20目前还没有关于糖酵解和通过还原羧基化利用谷氨酰胺之间联系的信息,尽管有报道称,除了少数例外,21,22只有谷氨酰胺,没有葡萄糖,癌细胞就不能生长。23综上所述,这些发现表明,不同类型癌症细胞的代谢差异可能会为寻找更有效的抗癌药物开辟新的靶点识别途径。24,25
为了达到这个目的,当然有必要详细阐明癌症特异性代谢途径。在这篇论文中,我们重建了癌细胞中谷氨酰胺和葡萄糖的利用途径,以一种新的方式阐明了它们与以NAD为中心的红/氧化过程的调节联系+/NADH和NADP+/NADPH公司。这里没有涉及NAD(P)途径之间的多种复杂关系+利用及其对谷氨酰胺和葡萄糖代谢的影响。NAD(P)+-利用系统包括(i)单ADP-核糖基化反应,(ii)聚ADPR聚合酶(PARP),(iii)ADP-核糖基环化酶和(iv)西尔图因。
肿瘤细胞谷氨酰胺利用途径
哺乳动物细胞代谢的生化反应网络是众所周知的,人类代谢的全基因组重建(针对许多细胞类型)是可用的。26最近,信号通过影响转录水平或翻译水平和翻译后水平的酶活性来控制代谢的作用在许多生理/病理条件中被阐明。27代谢图可用于注释任何给定酶的基因编码、每种底物的动力学参数、所需的辅因子、变构调节因子、,但是,仅仅测量酶的表达和代谢物的水平不足以表明在任何特定的细胞和任何特定的时间利用了哪种代谢途径。基于稳定标记前体的代谢组分析和系统建模方法是正在开发的工具,用于描述细胞如何调节其全球代谢。8,28,29,30文献中报道了许多癌症代谢再布线(CMR)的图谱,31,32,33但他们并没有特别关注氧化还原平衡。我们展示的地图图1和2这样做。
图1。
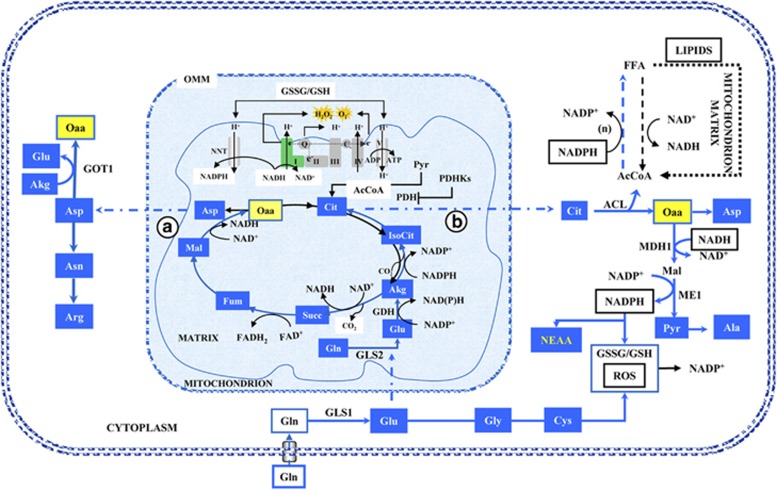
谷氨酰胺代谢重组示意图。进口谷氨酰胺通过Slc1a5进入一个复杂的代谢途径,如文中所述,因此其碳和氮都被用来促进癌细胞的生长和存活。乙酰辅酶A;ACL、ATP柠檬酸裂解酶;阿克,α-酮戊二酸;Ala,丙氨酸;精氨酸;天冬酰胺;天冬氨酸;Cit,柠檬酸盐;半胱氨酸;FFA,脂肪酸;Fum,富马酸盐;谷氨酸脱氢酶;谷氨酰胺;谷氨酸;甘氨酸;谷氨酸-草酰乙酸转氨酶;谷胱甘肽减少;GSSG,谷胱甘肽氧化;异柠檬酸,异柠檬酸;苹果酸;MDH1和2,苹果酸脱氢酶;ME,苹果酸酶;NEAA,非必需氨基酸;NNT,烟酰胺核苷酸转氢酶;Oaa,草酸盐;丙酮酸脱氢酶;PDHKs、丙酮酸脱氢酶激酶;丙酮酸;活性氧;琥珀酸
图2。
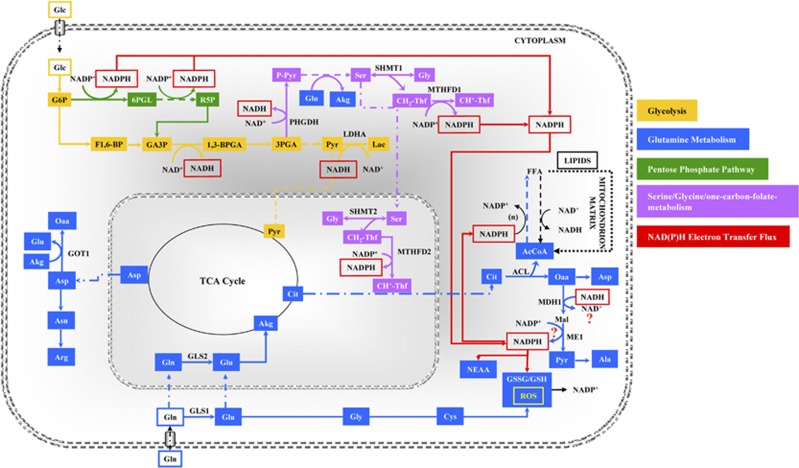
癌症代谢重组的概念图。维持谷氨酰胺还原羧化需要大量的糖酵解通量和复合物I功能障碍。黄色箭头表示糖酵解。蓝色箭头表示谷氨酰胺代谢,绿色箭头表示磷酸戊糖途径(PPP),明亮的紫丁香表示丝氨酸/甘氨酸/单碳-联体代谢,红色箭头表示NAD(P)H电子转移通量(ETF)。NAD(P)H ETF来源于丝氨酸转移途径,维持脂质合成、GSH对ROS的猝灭以及非必需氨基酸(NEAA)合成的还原步骤。缩写:1,3BPGA,双磷酸甘油酯;3PGA,3-磷酸甘油酯;6PGL,6-磷酸葡萄糖醇内酯;乙酰辅酶A;ACL、ATP柠檬酸裂解酶;阿克,α-酮戊二酸;Ala,丙氨酸;精氨酸;天冬酰胺;天冬氨酸;中国+-Thf,5-亚甲基四氢叶酸;中国2-Thf,5,10-亚甲基四氢叶酸;Cit,柠檬酸盐;半胱氨酸;F1,6BP,1,6二磷酸果糖;FFA,脂肪酸;G6P,6-磷酸葡萄糖;GA3P,3-磷酸甘油醛;Glc,葡萄糖;谷氨酰胺;谷氨酸;甘氨酸;谷氨酸-草酰乙酸转氨酶;谷胱甘肽减少;GSSG,谷胱甘肽氧化;乳酸;乳酸脱氢酶;苹果酸脱氢酶;ME,苹果酸酶;MTHFD1和2,亚甲基四氢叶酸脱氢酶(NADP+依赖);NEAA,非必需氨基酸;Oaa,草酸盐;磷酸戊糖途径;对丙酮酸,3-磷酸羟基丙酮酸;丙酮酸;R5P,5-磷酸核糖;活性氧;丝氨酸;SHMT1和2,丝氨酸羟甲基转移酶
如果我们只考虑碳通量,那么细胞通过转运蛋白(Slc1a5)摄取谷氨酰胺(Gln),该转运蛋白的表达受到多种致癌信号通路的刺激。11,34然后Gln通过细胞质GLS1脱氨基生成谷氨酸(Glu),并通过Slc25a11线粒体载体转移到线粒体基质中。否则,Gln可能被转运到线粒体中,在线粒体中GLS2将其脱氨基生成Glu。有趣的是,Myc控制GLS1的表达,14而p5335,36p53家族成员控制GLS2。37,38,39反过来,谷氨酸脱氢酶(GDH)将谷氨酸转化为α-酮戊二酸(Akg)。这种还原羧基化反应是通过异柠檬酸脱氢酶(IDH)2在线粒体中进行的,但也可能通过IDH1在细胞质中进行。已经描述了癌症细胞中Akg利用的两种途径,一种是逆时针(途径B),另一种是顺时针(途径A)。途径B中(图1),Akg羧基化为异柠檬酸盐(IsoCit),后者转化为柠檬酸盐,然后输出到细胞质中。细胞溶质酶ATP-柠檬酸-赖氨酸酶(ACL)将其分解为草酸盐(Oaa)和乙酰辅酶A(AcCoA)。AcCoA随后被用作脂质生物合成的构建块。40,41在途径A中,Akg遵循正常的TCA循环步骤,直到Oaa,然后Oaa被天冬氨酸转氨酶(GOT2)转化为天冬氨酸酯(Asp)并输出到细胞质中。反过来,天冬氨酸可以转化为天冬酰胺(Asn)和精氨酸(Arg),用于蛋白质合成。
NADH和NADPH参与CMR
途径A和B的许多反应(图1)是红/氧化反应,受两种特定辅酶NAD(H)和NADP(H)的可用性的强烈影响。关于线粒体基质中发生的反应,如前所述,复合物I功能障碍是癌细胞中相当常见的事件,42,43但TCA循环酶富马酸水合酶(FH)和琥珀酸脱氢酶(SDH)中也可能发生其他突变,44,45,46或在电子转移链(ETC)的其他络合物中,如络合物III。47所有这些改变都表明通过还原羧基化促进谷氨酰胺的利用。41,47,48安在体外对该反应的分析表明,为了产生足够的柠檬酸盐,通过异柠檬酸脱氢酶(IDH)催化的还原羧基化,线粒体NADPH/NADP较高+比率是必需的。49从这个角度来看,虽然途径B似乎是氧化还原平衡的,涉及两个反应,第一个反应(GDH)产生NADPH,另一个反应消耗NADPH。但提高谷氨酰胺的利用率需要,体内,烟酰胺核苷酸转氢酶(NNT)的活性,这是一种将电子从NADH转移到NADPH的线粒体酶。50此外,在FH缺陷或复合物III功能障碍的癌细胞中,TCA循环中Akg的氧化(路径A图1)需要维持同时还原羧化为异柠檬酸盐,从而表明用于将Akg转化为异柠檬酸盐的还原当量溢出。48虽然琥珀酸在FH缺陷癌细胞中积累,但在含有缺陷复合物I的癌细胞中却不会积累。48在后一种细胞中,Akg的每个分子都转化为Oaa,产生两个NADH分子和一个FADH分子2如前所述,NADH可以通过NNT转化为NADPH,为还原羧基化反应提供动力,或者它可以供给功能失调的ETC以产生ATP和消耗氧气。由于复合物I活性降低和几个OXPHOS基因表达水平不同,K-ras转化成纤维细胞中ETC的剩余活性约为正常细胞的50%。10与这些数据一致,胡等9已经证明了在体外激活K-rasG12伏导致复合物I的破坏。具体而言,使用基于SILAC(细胞培养中氨基酸的稳定同位素标记)的质谱法,作者观察到几种复合物I成分(NDUFA2、NDUFA4、NDUVA5、NDUFA11、NDU5A12、NDUSA13、NDUFB4、NDUFB6和NDUFB7)的水平降低。9迄今为止,一些有趣的研究表明致癌K-ras与癌细胞线粒体功能障碍之间的联系。51,52,53,54此外,这种联系可能导致TCA循环中谷氨酰胺利用的代谢重编程,以维持细胞氧化还原动态平衡。8,54,55总之,ETC或TCA循环成分的失活(大多数癌细胞的特征)设定了能够通过还原羧基化促进谷氨酰胺利用的条件。事实上,它导致NADH/NAD的增加+在线粒体基质中,通过NNT的作用,决定NADPH/NADP的增加+维持Akg的还原羧基化所需的。
虽然有许多关于癌症细胞中谷氨酰胺利用所引发的生物反应的研究结果,但谷氨酰胺代谢产物柠檬酸盐和天冬氨酸盐的命运,即从线粒体输出到细胞质,部分尚未确定。我们在下面提出了一个新的解释,支持这一概念,即糖酵解维持了谷氨酰胺完全利用的还原当量。
在癌细胞中,ACL的表达强烈上调56IDH1和IDH2催化的还原性谷氨酰胺代谢导致脂质生物合成增强。41与此相关的是,脂肪酸合成酶的过度表达给癌细胞带来生长和生存优势,57相反,抑制ACL抑制肿瘤细胞生长。58综上所述,这些发现表明,从谷氨酰胺衍生的AcCoA中持续产生脂质,这对癌细胞的生长和生存是必要的。
将AcCoA转化为脂质所需的NADPH的代谢来源是什么?在活跃增殖的细胞中,氧化戊糖磷酸途径(PPP)一直被认为是NADPH的主要来源,苹果酸酶在某些细胞类型中也很重要。11,59最近,有报道称,四氢叶酸(THF)依赖途径也有类似贡献。60这一途径源于丝氨酸,其从糖酵解中间体3-磷酸甘油酸(3Pga)产生需要磷酸甘油酸脱氢酶(PHGDH),这是一种产生NADH的酶,其功能在许多癌细胞中受到强烈刺激(图2).61,62,63,64在从丝氨酸转化为甘氨酸(在癌细胞中受刺激)的过程中,形成亚甲基四氢叶酸,其氧化生成NADPH(图1).64定量通量分析表明,脂质的合成是利用NADPH生成的大部分的途径。60通过追踪利用NADPH作为辅因子的分区路径中的氢,估计了线粒体或胞浆中发生的反应通量。65这种有趣的新代谢组学方法,允许胞质和线粒体NADPH区室鉴定,表明大多数线粒体NADPH和甘氨酸是由线粒体丝氨酸羟甲基转移酶(SHMT2)和亚甲基四氢叶酸脱氢酶2(MTHFD2)产生的。与Lewis及其同事一致,叶酸代谢分区的数学模型已在胚胎组织和癌细胞中确定,SHMT2和MTHFD2主要支持胞浆嘌呤和嘧啶的合成通过甲酸盐的出口。66此外,最近的一项研究表明,SHMT2在低氧条件下被致癌myc的癌细胞激活,并且SHMT2的敲除显示细胞NADPH/NADP降低+比率。67采用这种方法,可以研究PHGDH生成的NADH是否通过苹果酸脱氢酶(MDH1)和苹果酸酶(ME1)的联合活性转化为NADPH(图2). 如果是这样,那么人们可以回答这样一个问题:丝氨酸/甘氨酸糖基化转移产生的还原力是否被用来增加NADPH/NADP+比率,从而刺激非必需氨基酸(NEAA)的合成并增加活性氧猝灭所需的还原/氧化谷胱甘肽比率(GSH/GSSG)(图2).
众所周知,大多数癌细胞不能仅依靠谷氨酰胺生长,而需要积极的葡萄糖代谢通过糖酵解23以及购买力平价。68所述的糖酵解丝氨酸/甘氨酸转移可被视为促进谷氨酰胺利用的一种手段,谷氨酰胺利用可促进癌细胞的生长和存活。
CMR对成长和生存的作用
为了更好地理解CMR的生理作用,有意思的是定义生长的构建块的供应,这些构建块来自于CMR的两个过程:糖酵解和通过还原羧基化利用谷氨酰胺。鉴于蛋白质是最大的细胞大分子成分,约占标准哺乳动物细胞干重500 pgr的70%,26我们利用报告的每种氨基酸在总蛋白中的比例贡献,并计算每个NEAA必须从代谢中获得的相应分子数(图3a). 已知合成途径的分配表明,“从头开始'癌细胞的蛋白质合成活性来源于谷氨酰胺代谢(图3b). 由于缺乏特定的示踪实验,虽然丙酮酸也可能由谷氨酰胺产生,但从丙酮酸衍生的氨基酸被指定为糖酵解途径(图1). 当然,这只是对谷氨酰胺在癌细胞生长中的作用的粗略估计,因为谷氨酰胺还提供了大量核苷酸和脂质。
图3。
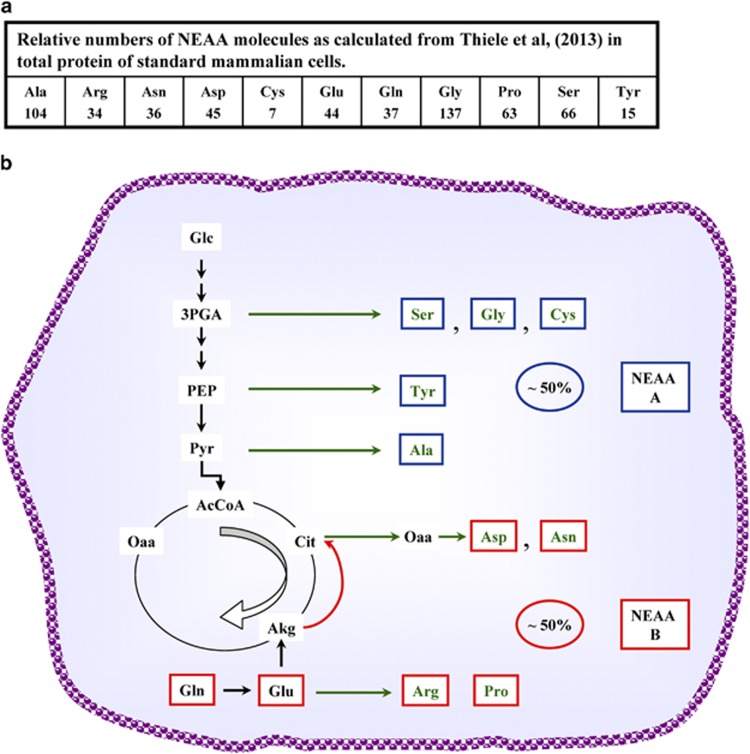
NEAA合成示意图。(一)每个氨基酸在总蛋白质中的比例贡献示意图,以及必须从代谢中获得的每个NEAA的相应分子数。(b条)癌症细胞中谷氨酰胺利用产生的NEAA合成示意图
对生长中哺乳动物细胞蛋白质周转率的全蛋白质分析表明,大多数蛋白质的半衰期从几分钟到几小时不等。因此,蛋白质生物合成活性的很大一部分用于维持稳定水平,而不是为新细胞生产蛋白质。69在这种情况下,营养过剩促进了线粒体吞噬过程(线粒体自噬消除),导致线粒体功能障碍和去极化,70特别令人感兴趣。事实上,在癌细胞中可以观察到线粒体在有丝分裂之前的碎片化,当葡萄糖和谷氨酰胺的摄取受到刺激时就会发生。59至于ATP的使用,维持所需的金额从头开始“细胞生长远远低于基础细胞维持所需的细胞生长。61,62来自广泛蛋白质降解的氨基酸可能被循环利用或进入尿素循环,产生多胺和脯氨酸,这两种物质在癌细胞中均被上调。71,72
考虑到谷氨酰胺衍生的氨基酸是从Oaa和Akg获得的氨基酸,它们也是TCA循环的中间产物,与从葡萄糖中提取骨架碳和从谷氨酰胺中提取氨基的正常细胞相比,癌细胞利用谷氨酰胺通过还原羧基化合成氨基酸的生理优势是什么?
第一个明显的区别是从环境中立即获得Gln和Glu,从而避免细胞从TCA循环产生的Akg开始生物合成。第二个是几乎所有的Gln,无论遵循哪种代谢途径,都会转化为Oaa。因此,中所示的NEAA(B)图3b可以快速生产。
此外,CMR确保谷氨酰胺利用率与糖酵解刺激率相协调,糖酵分解刺激率是癌细胞的特征。事实上,糖酵解对谷氨酰胺途径具有主控作用,通过调节丝氨酸/四氢呋喃的转移来实现,NADPH的生成速率(可能还有NADH)在谷氨酰胺途径的几个步骤中被利用。相反,正常细胞以一种更严格的方式控制NEAA的产生速率:正是丙酮酸盐对Oaa的再填充速率允许中间产物Akg和Oaa在TCA循环之外的利用。
综上所述,到目前为止讨论的结果首次指出NAD(H)和NADP(H)代谢在维持癌细胞生长中的强大调节作用。
NADH与肿瘤侵袭性
许多实验的结果支持了NAD代谢在肿瘤发展中的相关性,其中NAD(H)的可用性是通过遗传或生物化学手段控制的。最近,NAMPT受到了相当大的关注(图4)作为一个潜在的治疗靶点,因为一些报道显示其在几种73,74相反,它通过基因或化学手段下调抑制肿瘤细胞生长在体外在肿瘤异种移植中也是如此。75,76,77NAMPT最广泛使用的抑制剂之一是FK866,它正在作为一种抗癌药物开发。24,78
图4。
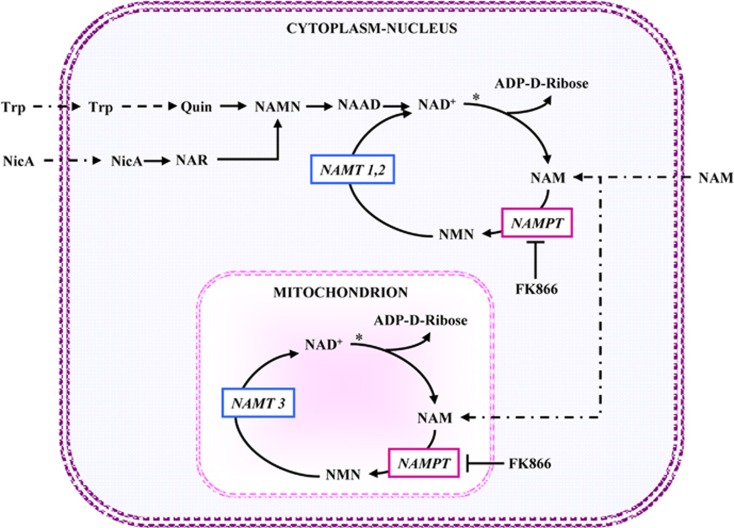
NAD±生物合成途径图。澄清“体内“测试中描述的实验,NAD+生物合成途径被简要报道:NAD+可以由色氨酸合成(从头开始合成)或烟酸-NicA-或烟酰胺-Nam-(补救途径)。北美+合成通过色氨酸通过几个反应进行,从而合成喹啉酸(Quin):然后转化为烟酸单核苷酸(NAMN),再转化为脱氨基-NAD(NAAD),最后转化为NAD+不同的是,在NAD的主要抢救途径中+合成时,NAM可以通过烟酰胺磷酸核糖基转移酶(NAMPT)转化为烟酰胺单核苷酸(NMN),NMN进一步转化为NAD+通过NMN腺苷酸转移酶(NMNAT 1和2核和细胞质亚型,NMNAT3线粒体亚型)。星号表示NAD催化的反应+糖水解酶,与ADP核糖基环化酶家族有密切的相似性。然而,所有NAD+-上述转化酶家族(即单ADP-核糖基转移酶,PARP秒,sirtuins)与ADP核糖基环化酶/NAD共享+糖水解酶——NAD释放NAM的特性+
当复合物I活性有缺陷的人类乳腺癌细胞被转染以表达酵母NADH脱氢酶Ndi1时,它们氧化线粒体NADH的能力被恢复,同时,它们在异种移植物中的肿瘤形成能力被显著降低。79鉴于Ndi1表达增强了NAD+/NADH在整个细胞和线粒体中的比例,通过提供NAD分析其影响+前体、NicA和NAM80同一复合物I缺陷乳腺癌细胞。用NicA和NAM治疗后,异种移植物的肿瘤形成能力显著降低,这两种药物都能补充NAD水平+线粒体和细胞质/核室(图4). 此外,NAD的供应+在动物模型中去除原发性乳腺癌后,饮用水中的前体物质可以提高生存率。79与之前的观察结果一致,这些发现证实ETC或TCA循环酶的功能障碍是通过还原羧基化触发谷氨酰胺利用所必需的,并表明NADH/NAD可能发挥调节作用+谷氨酰胺利用率。
用FK866治疗可显著降低NAD+可用于3-磷酸甘油醛脱氢酶(GAPDH)。一方面,这种抑制导致被抑制的GAPDH步骤上游糖酵解中间产物的积累,同时增加了进入PPP的流量68另一方面,减少了进入丝氨酸/四氢呋喃途径的碳流量,并且在非限制性谷氨酰胺存在的情况下,减少了TCA循环中的碳流量。77
癌细胞生长对NAD(H)的依赖性似乎比之前描述的要复杂得多。例如,已经证明NAD的减少+通过shRNA抑制NAMPT的表达而获得的水平可以使肿瘤细胞更具攻击性,79而用FK866治疗,靶向相同的酶,81导致癌细胞死亡在体外和体内.76这种差异支持了这样的观点,即FK866除了对NAMPT具有抑制作用外,还可能对癌细胞具有额外的细胞毒性。79
此外,最近的数据表明,FK866对NAMPT的药理学抑制可以通过拯救NAD来抵消+由NAM以外的前体产生,尤其是NMN和烟酰胺核苷(NR)。82具体来说,不同的FK866治疗的肿瘤细胞受到两种能够产生NAD的胞外酶的影响+前体。CD73(从NAD生成NR+和NMN)启用,而CD38(生成无法转换为NAD的NAM+由于NAMPT抑制)损害细胞内NAD+生物合成和细胞活性。
最后,为了更严格地评估FK866治疗后观察到的结果,应该澄清NAD(H)的线粒体池是否对FK866敏感,因为文献数据在这一点上并不一致。80,83
结论和未来展望
近年来,为了开发大量新的靶向抗癌药物,人们对促进癌症暴发和进展的信号转导和转录事件给予了大量关注。不幸的是,由于耐药分子机制的频繁反叛,这些药物对患者的疗效往往是暂时的。84最近人们意识到许多信号通路都会汇聚到一个共同的底线,即以线粒体功能障碍、糖酵解增强和通过还原羧基化反应刺激谷氨酰胺利用为特征的代谢重组,这为开发抗癌药物的新战略提供了可能。85事实上,尽管对Marin-Valencia的研究等研究表明,来源于胶质母细胞瘤细胞移植的肿瘤利用葡萄糖维持合成代谢过程,积累谷氨酰胺,86一些研究已经证明了谷氨酰胺酶抑制剂作为BPTES(双-2-(5-苯基乙酰氨基-1,2,4-二硫唑-2-基)乙基硫醚)和CB-839的功效在体外和体内.21,87,88,89,90,91此外,由于代谢差异主要是由不同类型的肿瘤表型引起的,因此应该对人类肿瘤进行详细的代谢分析,以阐明发生CMR的癌症类型。对于这些癌症类型,本文提出的癌症重新布线代谢的概念图为开发旨在靶向CMR限速节点的药物发现策略提供了合理的基础。此外,要使这种药物战略真正有效,需要对一些定量方面进行评估。
首先,利用各种代谢组学技术,如气相色谱-质谱(GC-MS)或液相色谱(LC-MS)结合稳定同位素示踪剂,通过代谢通量分析(MFA)进行代谢通量重建和酶功能定量评估。8,29,41,92,93,94此外,这种实验/计算方法应允许估计各种途径中的通量(尤其是那些目前不太为人所知的途径,例如丝氨酸/四氢呋喃途径产生的还原力的利用60,65在各种癌细胞中在体外也可能是体内).95考虑到目前的技术甚至可以准确描述福尔马林固定石蜡包埋肿瘤活检样本的代谢曲线,96回顾性研究是可行的。还可以通过NAD途径前体或抑制剂的治疗获得更多关于线粒体和细胞质NAD(P)H/NAD(P)比值在维持癌症代谢重组中的作用的信息,同时仔细考虑目前正在讨论的方面。定量动态模型的发展97癌症新陈代谢的重组及其调控将为新的认识增加预测能力(方框1).85
方框1。癌症代谢重组的计算模型和设计原则。
深入理解癌症代谢重组需要发展计算模型85,97这应该至少考虑到概念图中包含的所有代谢反应图1和2.
文献中报道了一些研究,通常涉及特定途径,如糖酵解或TCA循环。目前流行两种方法:定量数学模型或全基因组代谢模型,这两种方法都有局限性,也有新的见解。例如,有趣的是对Warburg效应的全基因组研究,该效应将代谢步骤与癌细胞迁移联系起来101以及NCI60肿瘤衍生细胞系中丝氨酸/甘氨酸转化率的数学模型,该模型显示了甘氨酸对癌细胞中嘌呤、ATP和NADPH生成的贡献。102下一步是开发可靠的定量模型,将代谢重组与增强癌细胞的生长和存活联系起来。同时,对区分癌细胞和正常细胞的设计原则进行评论可能会很有趣。人们常说,癌细胞新陈代谢效率低下,因为它们仅部分利用葡萄糖的化学势,进行乳酸发酵并分泌乳酸,103而正常细胞效率更高,因为它们在糖酵解和TCA循环中充分利用葡萄糖。但谷氨酰胺(哺乳动物细胞的另一种相关底物)的情况正好相反在体外'和'体内'). 癌细胞使用谷氨酰胺的碳和氮,而正常细胞只使用氮并分泌谷氨酸。8
在癌细胞中,进入PPP或丝氨酸/甘氨酸途径的每个葡萄糖分子的还原型辅酶的产生是不同的:对于在第一个途径中形成的两个NADPH分子,在第二个途径中有两个NADH和两个NADPH。
关于新陈代谢与生长的关系,如果有人回忆起蛋白质是生物量的最大组成部分,那么就必须关注氨基酸的代谢途径(图3). 在正常细胞中,TCA循环中氨基酸的利用率受到丙酮酸向TCA循环补充Oaa的速度以及作为氨基酸提供者将影响TCA循环效率的条件的可用性的限制(有利的ADP/ATP比率、电子转移链的功能和执行速率以及氧气的可用性)。在癌细胞中,A和B通路的少数酶图1和2NEAA B的生成速度仅取决于PPP和/或丝氨酸/甘氨酸途径再生NADPH的能力,而NEAA A的生成速度预计会随着糖酵解速度的增加而增加。
总之,在癌细胞中,糖酵解及其相关途径似乎对谷氨酰胺利用途径具有主前馈控制,这种情况可能与维持肿瘤形成能力有关体内'.
由于肿瘤发生所需的代谢条件是Complex I功能障碍和葡萄糖和谷氨酰胺利用增强,本文提出的概念图也可能为报告的癌症发病率随年龄增长而增加提供新的解释。98复合物I活性降低是衰老过程中新陈代谢下降的特征;99这一事件为能够刺激营养吸收的体细胞突变的致癌表达提供了有利的基础。如果没有与衰老(或其他原因)相关的线粒体功能障碍,那么简单刺激葡萄糖和谷氨酰胺摄取可能是非肿瘤性的。对癌症代谢重组的更深入理解图1和2以及其调节可能阐明sirtuins是否如Wu及其同事所提出的那样在年龄诱导的肿瘤发生中具有普遍作用,100并可能表明激活复合物I活性以防止衰老和年龄诱导的肿瘤发生的方法。
致谢
我们衷心感谢安东尼奥·德·弗洛拉对本文前一版本提出的宝贵意见和建议,以及匿名推荐人的周到评论。这项工作得到了SysBioNet项目的资助,该项目是欧洲研究基础设施战略论坛(ESFRI)意大利路线图的MIUR倡议,由洛杉矶和Tecnomed基金会提供给DG。
词汇表
- 1,3BPGA(英国石油天然气协会)
双磷酸甘油酯
- 3PGA公司
3-磷酸甘油酯
- 6加仑
6-磷酸葡萄糖内酯
- AcCoA公司
乙酰辅酶A
- 国际计算语言学协会
ATP柠檬酸裂解酶
- 阿克格
α-酮戊二酸
- 阿拉
丙氨酸
- 精氨酸
精氨酸
- Asn公司
天冬酰氨
- 天冬氨酸
天冬氨酸
- 中国+-Thf公司
5-亚甲基四氢叶酸
- 中国2-Thf公司
5,10-亚甲基四氢叶酸
- Cit公司
柠檬酸盐
- 1.6BP财年
1,6-二磷酸果糖
- 金融流量账户
脂肪酸
- 福姆
富马酸盐
- G6P公司
6-磷酸葡萄糖
- GA3P基因
3-磷酸甘油醛
- GDH公司
谷氨酸脱氢酶
- Glc公司
葡萄糖
- 格林
谷氨酰胺
- 谷氨酸
谷氨酸盐
- 格莱
甘氨酸
- 得到了
谷氨酸-草酰乙酸转氨酶
- 谷胱甘肽
还原型谷胱甘肽
- GSSG公司
氧化型谷胱甘肽
- Isocit公司
异柠檬酸盐
- 拉克
乳酸盐
- LDHA公司
乳酸脱氢酶
- 马尔
苹果酸
- MDH 1和2
苹果酸脱氢酶
- 我
苹果酸酶
- MTHFD1和2
亚甲基四氢叶酸脱氢酶(NADP+依赖)
- 美国国家航空航天局
非必需氨基酸
- NNT公司
烟酰胺核苷酸转氢酶
- Oaa公司
草酸盐
- PDH公司
丙酮酸脱氢酶
- PDHK公司
丙酮酸脱氢酶激酶
- 购买力平价
戊糖磷酸途径
- P-Pyr公司
3-磷酸羟基丙酮酸
- 梨
丙酮酸盐
- 5便士
5-磷酸核糖
- ROS公司
活性氧物种
- 序号
丝氨酸
- SHMT1和2
丝氨酸羟甲基转移酶
- 成功
琥珀酸盐

