摘要 线粒体功能障碍会导致一系列早期神经疾病,并导致神经退行性疾病。 然而,神经损伤的机制尚不清楚,因为很难从患者身上获取相关组织,而且合适的模型也有限。 因此,我们评估了CI(复合物I)缺陷患者神经相关原代细胞系的线粒体功能 恩杜夫斯4 KO(敲除)鼠标( 恩杜夫斯4 飞/飞 )线粒体疾病LS(Leigh综合征)和MEFs(小鼠胚胎成纤维细胞)的建模方面。 尽管CI的结构和功能在所有方面都受到了损害 恩杜夫斯4 飞/飞 细胞类型,MEFs中线粒体膜电位选择性受损,与CI依赖性ATP合成减少有关。 此外,只有在 恩杜夫斯4 飞/飞 主要MEF。 相反, 恩杜夫斯4 飞/飞 初级等皮质神经元和初级等皮质星形胶质细胞仅显示ATP生成受损,而线粒体膜电位无变化。 因此 恩杜夫斯4 飞/飞 即使以维持线粒体膜电位为代价,小鼠的部分原因可能是神经元和星形胶质细胞中ATP的严重耗竭。 这可能会保护细胞免受死亡,但最终会损害神经元和星形胶质细胞的细胞功能。 此外,从复合物II到CI的RET(反向电子转移)在神经元中比MEF或星形胶质细胞更显著,在 恩杜夫斯4 飞/飞 细胞。
关键词: 代谢应激、线粒体疾病、小鼠模型、神经病理学、原代细胞、活性氧物种
缩写: BN-PAGE,蓝色native-PAGE; CI–CV,复合物I–复合物V; CS,柠檬酸合成酶; CV,复合物V; 二氢乙硫醚; FCCP,羰基氰化物 第页 -三氟甲氧基苯腙; H(H) 2 O(运行) 2 ,过氧化氢; HBSS、Hanks缓冲盐水; KO,击倒; LS,Leigh综合征; MEF,小鼠胚胎成纤维细胞; NBM,神经基础培养基; 氧化磷酸化; PI,碘化丙啶; RET,反向电子转移; 活性氧; ΔΨ 米 ,线粒体膜电位
简短摘要 我们描述了神经相关原代星形胶质细胞和神经元的代谢反应如何与分离自 恩杜夫斯4 飞/飞 线粒体疾病小鼠模型。
简介 线粒体疾病是一种罕见疾病的集合,每种疾病都以线粒体功能障碍为特征,估计每5000名活产婴儿中就有一例受到影响[ 1 ]. 这种疾病的表现形式多种多样,可能涉及任何组织、任何严重程度和任何年龄[ 2 ]. 然而,通常情况下,表现出高能量需求的代谢活性更强的组织参与其中,尤其是大脑[ 三 , 4 ]. 该病的遗传基础也很复杂,包括OXPHOS(氧化磷酸化)系统在内的100多个功能线粒体所需的基因发生了致病性突变[ 4 , 5 ]. 虽然疾病相关OXPHOS基因的突变与广泛的经典线粒体疾病有关,但其中最常见的是LS(Leigh综合征)[ 4 ].
LS是一种进行性神经退行性疾病,患者表现出广泛的神经系统症状,包括共济失调、呼吸异常、视神经萎缩、发育迟缓、听力障碍和癫痫[ 6 – 8 ]. 病理学上,该病的特征是基底节、丘脑或脑干对称性双侧病变,血液和/或脑脊液中乳酸增加,神经胶质增生和神经元脱髓鞘[ 6 – 8 ]. 虽然LS的大体生化和临床表现特征明确,但从感兴趣的神经区域获取新鲜患者样本的困难限制了我们在细胞水平上对生化特征的理解。
为此,最近产生了一些原发性线粒体功能障碍的动物和细胞模型系统(在[ 9 , 10 ]),包括 恩杜夫斯4 飞/飞 和 恩杜夫斯4 缺少OXPHOS CI(复合物I)亚单位NDUFS4的KO(敲除)小鼠[ 11 , 12 ]. 与LS患者一样,NDUFS4缺陷小鼠出现双侧脑干高强度损伤[ 13 , 14 ]. 小鼠也表现出与LS患者相似的临床表现[ 11 – 14 ]. 此外,脑特异性 恩杜夫斯4 KO小鼠在生化和表型上与整个动物KO几乎没有区别,证实了该病的神经基础及其临床特征[ 13 ]. 由于NDUFS4缺陷小鼠重述了LS的许多共同特征,这些小鼠提供了机会来研究线粒体系统和动力学在神经环境中是如何受到影响的。
到目前为止,NDUFS4缺乏小鼠的全脑制剂分析[ 12 , 13 ]发现,像NDUFS4缺陷患者和小鼠成纤维细胞[ 15 , 16 ],CI结构和功能被破坏。 在每个系统中,CI在BN-PAGE(蓝色原生-PAGE)上形成≈830 kDa的致残复合体。 此外,发现CI缺陷影响了分离的脑线粒体中CI依赖性ATP的合成[ 12 ]. 也有证据表明NDUFS4缺乏小鼠嗅球中的蛋白质受到氧化损伤,嗅球是受疾病影响的大脑区域之一[ 13 ]. 考虑到NDUFS4缺乏的患者和成年小鼠成纤维细胞产生的ROS(活性氧物种)数量增加,包括超氧化物(O 2 •− )和H 2 O(运行) 2 (过氧化氢)[ 17 – 19 ]. 因此,有理由预计,神经相关细胞类型中CI的破坏也会导致ROS的生成增加,从而促进神经病变。 然而,在 恩杜夫斯4 KO小鼠初级中脑神经元 2 •− 分离线粒体和H的生成 2 O(运行) 2 与对照组相比,整个细胞的产量正常[ 20 , 21 ].
因此,表征细胞类型特异性疾病机制,如活性氧动力学,将增加我们对LS疾病发病模式的了解。为此,我们试图从细胞中分离和表征初级等皮质星形胶质细胞和等皮质神经元 恩杜夫斯4 飞/飞 鼠标。 尽管星形胶质细胞是哺乳动物大脑中最丰富的细胞类型[ 22 , 23 ]它们在线粒体疾病中的作用往往被忽视。 星形胶质细胞通过诸如神经递质循环和细胞外钾同质化等过程与神经元进行代谢错综复杂的联系,以实现适当的细胞功能[ 23 , 24 ]. 此外,它们与LS患者和NDUFS4缺乏小鼠的疾病进展有关,激活的星形胶质细胞浸润到受影响的大脑区域是这两个系统的特征[ 7 , 13 ]. 同时,我们还研究了 恩杜夫斯4 飞/飞 MEFs(小鼠胚胎成纤维细胞),作为我们研究中非神经组织的对照细胞类型。 我们对OXPHOS系统及其相关线粒体过程的研究 恩杜夫斯4 飞/飞 因此,星形胶质细胞、神经元和MEF提供了在细胞类型特异性水平上研究疾病机制的机会。
实验 鼠标模型 这个 恩杜夫斯4 飞/飞 小鼠是由B2 SINE(短间隔核元素)自发插入 NADH脱氢酶(泛醌)Fe-S蛋白4(Ndufs4) 如前所述的基因[ 12 ],并在BALB/c背景下维护。 根据默多克儿童研究所动物伦理委员会(A662)批准的协议,繁殖杂合小鼠以产生幼崽和胚胎。 将小鼠置于12小时的光/暗循环中,并提供 随意 获得食物和水。
细胞培养方法 如前所述,分别从3-5日龄幼鼠和17-18日龄胚胎等皮质中分离出初级等皮质星形胶质细胞和等皮质神经元[ 25 , 26 ]. 简单地说,组织收集在不含镁的HBSS(Hanks缓冲盐水溶液)中 2+ 和钙 2+ (Gibco),并在0.6%(w/v)胰蛋白酶和2.78 mM葡萄糖中消化[30分钟,37°C,5%(v/v)CO 2 ]. 在用含有100单位/ml DNAse I(Invitrogen)的1 mg/ml大豆胰蛋白酶抑制剂(Invit罗gen)中和胰蛋白酶后,细胞被机械分离并在含有mg的HBSS中稀释 2+ 和钙 2+ 将分离的星形胶质细胞在含有25mM葡萄糖的DMEM(文本中缩写为“葡萄糖”培养基;Thermo-scientific)中稀释,并补充10%(v/v)FCS,并将其置于预涂有50μg/ml poly的烧瓶中 D类 -赖氨酸。 在汇合处,将星形胶质细胞培养物在200 rpm、37°C下摇晃14-16 h,以去除污染细胞类型。 分离的神经元在补充了B-27(Invitrogen)的NBM(神经基础培养基;Invitrogen)中稀释,292 mg/l L(左) -谷氨酰胺,并在烧瓶中镀上50μg/ml聚乙烯 D类 -赖氨酸或预先涂有50μg/ml聚乙烯的盖玻片 D类 -赖氨酸和10μg/ml层粘连蛋白。 通过免疫细胞化学(补充材料和方法部分和补充图S1)评估神经元和星形胶质细胞培养物的纯度。
从受精后12.5-14.5天的胚胎中分离出初级MEF。 去除头部和内脏器官后,组织被机械破坏,随后用0.125%胰蛋白酶消化(30分钟,37°C,5%CO 2 ). 然后,将细胞机械分离、造粒,并将其置于涂有0.1%明胶的培养瓶中的葡萄糖培养基中。
细胞保持在37°C和5%CO 2 在加湿的房间里。 所有细胞隔离缓冲液(HBSS)和培养基均含有60.3 mg/l青霉素和100 mg/l链霉素。 线粒体DNA缺陷(ρ°)和对照(ρ + )鼠标LM(TK − )单元格[ 27 ]用50μg/ml尿苷维持在葡萄糖培养基(如上所述)上。 如前所述,星形胶质细胞和MEF在含有5 mM半乳糖(文中缩写为“半乳糖”培养基;Gibco)的DMEM中交替培养,补充10%透析的FCS和292 mg/l L(左) -谷氨酰胺。
线粒体分离 如前所述,从原代细胞中分离出线粒体[ 28 ].
CI和CS(柠檬酸合酶)活性 如前所述,在30°C下测定原代细胞培养物中相对于总蛋白的CI和CS活性[ 29 ]. 简单地说,细胞在缓冲液(200 mM甘露醇、70 mM蔗糖、5 mM HEPES游离酸、1 mM EGTA、pH 7.2)中用Teflon-glass均质器和台式钻机以1000 rpm的速度进行均质。 碎片在600时通过低速旋转被清除 克 对于CI活性测量,样品在含有辅酶Q的缓冲液中稀释 1 在鱼藤酮存在或不存在的情况下,NADH、氰化钾和抗霉素A。 CI的活性在分光光度计中测量为340 nm下3分钟NADH氧化的鱼藤酮敏感性速率。 对于CS活性测量,样品在含有5,5′-二硫代双-(2-硝基苯甲酸)、草酰乙酸和乙酰辅酶A的缓冲液中稀释。 用分光光度计在412 nm处监测3 min以上硫代硝基苯甲酸盐阴离子生成速率。
蛋白质印迹分析 将原代细胞颗粒溶解在含有蛋白酶抑制剂(Roche)的RIPA缓冲液中[50 mM Tris/HCl、150 mM NaCl、1%(v/v)Triton X-100、1%(w/v)Na脱氧胆酸盐、0.1%(w/v)SDS、1 mM EDTA,pH至7.4],在LDS(十二烷基硫酸锂)样品缓冲液(Invitrogen)中稀释,热变性(10分钟,60°C) 并在10%Novex双三凝胶(Invitrogen)上分离。 将凝胶转移到0.45μm PVDF膜(Millipore)上进行免疫修饰。 用抗NDUFS4(MitoSciences,MS104)或抗SDHA(Molecular Probes,A-11142)探测膜。
BN-页码 将细胞颗粒溶解在1%洋地黄素或1%Triton X-100中,并使用BN-PAGE分离蛋白质复合物[ 30 ]. 简单地说,在4–10或4–13%BN-PAGE凝胶(Invitrogen)上,每个通道分离50–80μg全细胞裂解物。 将凝胶转移到PVDF膜(Millipore)上进行免疫修饰。 用针对CI亚单位NDUFA9的抗体探测膜[ 31 ]、CII亚单位SDHA(Invitrogen)或复合III亚单位Core1(Invit罗gen)。
ATP合成分析 如前所述,在原代细胞培养物和分离线粒体的技术副本中测量ATP合成速率[ 32 ]. 简言之,10μg细胞或2μg分离的线粒体在含有40–80μg/ml洋地黄素(仅限全细胞)和有/无抑制剂底物的ATP合成缓冲液中稀释10倍[琥珀酸(10 mM);丙二酸(1 mM),谷氨酸(10 M M);丙酮酸(10 m2);苹果酸(10 m3);鱼藤酮(2.5μM)] 并在37°C下培养20分钟。 用冰上的高氯酸停止反应,用氢氧化钾和MOPS中和反应。 使用基于荧光素酶的分析(Roche,11699695001)在微孔板阅读器(BMG Labtech,FLUOstar OPTIMA)中测定样品中的ATP浓度。 速率报告为琥珀酸盐+鱼藤酮测量值的比率。
膜电位测量 细胞培养在96个黑色板(2×10)中 4 原代星形胶质细胞和MEF的细胞/孔,5×10 4 ρ°和ρ的电池/孔 + LM(油箱 − )细胞),并用含有75 nM四甲基罗丹明甲酯(TMRM;Sigma)和2μg/ml Hoechst-33258(Sigma 2 ). 随后,在ΔΨ 米 (线粒体膜电位)在至少三个孔中测定为TMRM与Hoechst荧光的比值(BMG Labtech,FLUOstar OPTIMA)。 用CI抑制剂鱼藤酮(2.5μM)或原核FCCP(羰基氰化物)处理的细胞 第页 -三氟甲氧基苯腙); 西格玛; 20μM)作为对照。
或者,ΔΨ 米 通过显微镜在玻璃底培养皿(WPI,FD-35-100)中培养的细胞中测定。 细胞在含镁的HBSS中培养 2+ 和钙 2+ 含37.5 nM TMRM(30分钟,37°C,5%CO 2 ). 在37°C的DeltaVision OMX V3成像系统(Applied Precision)上,使用TRITC(四甲基罗丹明β-异硫氰酸盐)过滤器组(带油浸物镜(Olympus,IX71),并配备CoolSNAP HQ)对细胞进行可视化 2 相机(光度学)。 初始成像后,将20μM FCCP或2.5μM鱼藤酮添加到重新捕获的盘子和感兴趣区域中。 以最大强度对图像进行解卷积、裁剪和投影(Applied Precision,SoftWoRx v5.5)。 在每盘培养皿中选择超过60个细胞的富含线粒体的核周区域,并在FCCP或鱼藤酮应用前后测定平均TMRM荧光。
超氧化物(O 2 •− )检测 细胞培养在96个黑色板(2×10)中 4 细胞/孔),并用含有10μg/ml DHE(二氢乙硫酸氢钠;Sigma)和2μg/ml Hoechst-33258(40分钟,37°C,5%CO)的培养基培养 2 ). 细胞在PBS中清洗 2 •− 在至少三口井中测定的产量为DHE与Hoechst荧光的比值(BMG Labtech,FLUOstar OPTIMA)。 用CI抑制剂鱼藤酮(2.5μM)处理的细胞作为对照。
H(H) 2 O(运行) 2 侦查 H的比率 2 O(运行) 2 产量是按照前面描述的进行测量的[ 33 ]使用荧光探针Amplex Red在5μg分离的线粒体中添加底物和抑制剂组合,详见正文[琥珀酸(10 mM);丙二酸(1 mM)、谷氨酸(10 M M);丙酮酸(10 m2);苹果酸(10 m3);鱼藤酮(2.5μM)]。 H(H) 2 O(运行) 2 在20分钟内对产量进行荧光测量(BMG Labtech,FLUOstar OPTIMA)。
细胞死亡分析 如前所述,用50μM H处理细胞 2 O(运行) 2 在培养基中培养6h。 用胰蛋白酶收集细胞,并稀释至2×10 5 annexin-V结合缓冲液中的细胞数/ml(10 mM Hepes,140 mM NaCl,2.5 mM CaCl 2 pH至7.4)。 将500μl的等分样品与5μl膜联蛋白-V(Invitrogen)和1μg/ml PI(碘化丙啶,Sigma)在室温下共同孵育10分钟,然后使用FITC-A和PI-488-695/40-A激光通过流式细胞术(Becton Dickinson,LSR II流式细胞仪)进行分析。 使用正向和侧向散射参数(De Novo,FCS express V4)区分活细胞和死细胞。 健康细胞被认为是膜联蛋白V和PI双重阴性,凋亡细胞为膜联蛋白-V阳性,坏死细胞为膜连蛋白-V和PI双重阳性。 选通策略如补充图S2(A)和S2(B)所示。
统计 除非另有说明,误差条表示平均值的标准误差(S.E.M.)和 P(P) 值由双尾未配对生成 t吨 测试(GraphPad,PRISM V6.0b)并在≤0.05时报告。
结果 CI活动在年同样减少 恩杜夫斯4 飞/飞 细胞系 未检测到NDUFS4亚单位 恩杜夫斯4 飞/飞 检查的原代细胞类型( 图1 A) 与NDUFS4在小鼠中的系统性破坏一致,并与之前关于所有研究组织的报告一致[ 12 ]. 培养并没有改变这一点 恩杜夫斯4 飞/飞 初级MEF和星形胶质细胞在有利于OXPHOS系统(半乳糖)生成ATP的条件下。 CI活性在 恩杜夫斯4 飞/飞 标准培养条件下的原代MEF(+/+的21%)、星形胶质细胞(+/+中的23%)和神经元(+/+里的42%)(葡萄糖培养)( 图1 B) ,类似于关于小鼠组织的报告[ 12 ]. 为了解释线粒体体积和/或数量的变化,CI活性被归一化为CS活性,而CS活性不受NDUFS4缺失的影响( 图1 C) ●●●●。 值得注意的是,虽然CI相对于CS的活性在每个 恩杜夫斯4 飞/飞 细胞类型,神经元中相对于CS的CI活性低于星形胶质细胞和MEF( 图1 B) ●●●●。
图1。 中的CI活动 恩杜夫斯4 飞/飞 细胞系。
在细胞裂解物中检测CI亚单位NDUFS4的表达,所述细胞裂解物由+/+(白色条)和 恩杜夫斯4 飞/飞 使用CI亚基NDUFS4和CII亚基SDHA抗体作为负荷对照,通过Western blot分析在葡萄糖或半乳糖培养基(原代MEF、原代星形胶质细胞)或NBM(原代神经元)中生长的(灰条)小鼠( A类 ). 为了清晰起见,图像已经进行了数字裁剪。 CI活动相对于CS活动( B类 )和CS活动( C类 )在来自+/+和 恩杜夫斯4 飞/飞 通过分光光度法在葡萄糖培养基(原代MEF,原代星形胶质细胞)或NBM(原代神经元)上培养小鼠。 误差线=S.E.M。, n个 ≥7, P(P) 由双尾未配对生成的值 t吨 测验。 CI,复合物I; CII,复合体II; CS,柠檬酸合成酶。
结合之前从 恩杜夫斯4 飞/飞 小鼠[ 12 ]CI活性的组织特异性差异似乎不太可能是主要神经功能障碍的基础 恩杜夫斯4 飞/飞 鼠标。
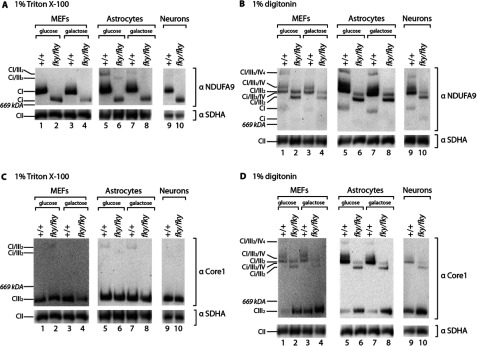
CI在 恩杜夫斯4 飞/飞 细胞系 接下来我们研究了CI在原代细胞系中的组装状态。 与之前的报告一致,我们使用BN-PAGE观察到缺少NDUFS4的CI形成≈830 kDA的残缺组装中间产物(CI),当细胞溶解在Triton X-100中以分离单个OXPHOS复合物时,其数量会减少( 图2 A: +/+,奇数车道; 恩杜夫斯4 飞/飞 ,甚至车道)[ 12 , 15 , 16 , 34 ]. 这与所有原代细胞的培养条件无关( 图2 A) ●●●●。 此外,当 恩杜夫斯4 飞/飞 细胞颗粒溶解在温和的去污剂洋地黄中,以保持与其他OXPHOS复合物的超分子相互作用( 图2 B和 2 D: +/+,奇数车道; 恩杜夫斯4 飞/飞 超复合物以较小的分子量出现,对应于含有受损CI的超复合物。同样,这与所有原代细胞的培养条件无关( 图2 B和 2 D) ●●●●。
图2。 呼吸道复合物的BN-PAGE分析 恩杜夫斯4 飞/飞 细胞系。
源自+/+和 恩杜夫斯4 飞/飞 将维持在葡萄糖或半乳糖培养基(原代MEFs、原代星形胶质细胞)或NBM(原代神经元)上的小鼠溶解在含有1%Triton X-100(A和C)或1%洋地黄苷的缓冲液中( B类 , D类 )和由BN-PAGE分离的蛋白质复合物。 将凝胶转移到膜上,并用针对CI亚单位NDUFA9的抗体进行免疫修饰( A类 , B类 )或针对CIII亚单位Core1( C类 , D类 ). CII亚单位SDHA用作负荷控制。 n个 =1–3.为了清晰起见,图像已进行了数字裁剪。 CI–IV,复合物I–IV; Ci,在无NDUFS4的情况下形成的残缺Ci。
此外,成熟复合物III(CIII)的水平 2 )在所有细胞和所有检测的培养条件下都是相似的( 图2 C) ●●●●。 然而,大量的游离CIII 2 超复合体中的非束缚态似乎增加了 恩杜夫斯4 飞/飞 单元格( 图2 D) 与含有残缺CI的超复杂物种丰度减少一致。
这些数据表明,CI本身和含有残缺CI的超复合体的组装在所有初级细胞中的洗涤剂溶解细胞颗粒中受到同样的破坏 恩杜夫斯4 飞/飞 细胞类型和培养条件。 因此,这些数据并没有进一步揭示疾病组织特异性本质背后的机制,也没有表明MEF和星形胶质细胞对半乳糖的适应性变化,以应对细胞ATP生成对OXPHOS的更大依赖。
ΔΨ 米 选择性受损 恩杜夫斯4 fky/fk(英尺/英尺) 年 主要MEF 如上所述,所有患者的CI活性和装配均严重受损 恩杜夫斯4 飞/飞 研究了原始细胞类型。 考虑到CI对IMM质子泵的整体性,可以预计这可能会影响ΔΨ 米 此外,半乳糖培养细胞会增加对OXPHOS的依赖性,这可能会进一步影响ΔΨ 米 然而,在基本条件下,ΔΨ 米 仅在 恩杜夫斯4 飞/飞 主要MEF( 图3 A和 三 B) 而在星形胶质细胞中表现正常( 图3 C和 三 D) 和神经元( 图3 E和 三 F) ●●●●。 更具体地说,ΔΨ 米 在中休息 恩杜夫斯4 飞/飞 葡萄糖培养基上的原代MEFs去极化(更多的电中性; 图3 A; 75%的+/+),并在半乳糖培养基上出现进一步去极化( 图3 B类; +/+的60%)。 作为对照实验,我们发现ΔΨ 米 在应用CI抑制剂鱼藤酮或原核细胞FCCP后,在所有培养条件下,所有类型的细胞都同样去极化( 图3 A– 三 F) ●●●●。 此外,使用不具备功能性OXPHOS系统的小鼠成纤维细胞ρ°细胞验证了方法学方法[ 27 ]. 在静止的ρ°单元中,ΔΨ 米 当用两个平板阅读器测量时,明显去极化(补充图S3A;ρ的62% + )和显微镜检查(补充图S3B和S3C;ρ的48% + )与之前在类似细胞中的观察结果一致[ 35 , 36 ].
图3。 ΔΨ分析 米 在里面 恩杜夫斯4 飞/飞 细胞系。
ΔΨ 米 通过线粒体中TMRM的积累进行测量,并与原始MEF和+/+(白色条)星形胶质细胞中的细胞数量相关 恩杜夫斯4 飞/飞 (灰条)小鼠,用葡萄糖维持( A类 , C类 )或半乳糖( B类 , D类 )中等。 ΔΨ 米 如图所示,用CI抑制剂鱼藤酮或原粒FCCP降低,或用CV抑制剂寡霉素调节。 ΔΨ 米 通过显微镜对初级神经元进行评估( E类 )通过测量阳离子染料TMRM在富含线粒体的核周区的积累,用鱼藤酮进行预培养和后培养。 使用≥2个细胞培养物分析每个培养皿中超过60个细胞。 显示了中初级神经元的代表性图像( F类 )与FCCP孵育前和孵育后。 误差线=S.E.M。, n个 ≥3 ( A类 – D类 ), P(P) 由双尾未配对生成的值 t吨 测验。 CI和CV,配合物I和V; ΔΨ 米 线粒体膜电位。
然而,初级MEF和星形胶质细胞对应用CV(复合V)抑制剂寡霉素的反应不同。 抑制CV会阻止电子流回线粒体基质,从而导致ΔΨ的超极化(更负电性) 米 当OXPHOS正在运行时。 在葡萄糖培养基上培养的原代MEF中( 图3 A) ,寡霉素对ΔΨ无影响 米 但它使ΔΨ超极化 米 当细胞在半乳糖上培养时( 图3 B) ●●●●。 相反,在原代星形胶质细胞中,ΔΨ 米 不考虑培养基,应用寡霉素后去极化( 图3 C和 三 D) 。 但这种现象并不是基因型特异性的,可以反映寡霉素对原代星形胶质细胞的毒性。
依赖CI的ATP合成能力在 恩杜夫斯4 飞/飞 原代星形胶质细胞和神经元,但仅在半乳糖培养的原代MEF中受损 ΔΨ 米 用于驱动许多线粒体过程。 其中最关键的一个,被认为是线粒体的核心功能,是通过CV合成ATP[ 37 ]. 在这种情况下,我们试图确定透性化中CI依赖型ATP合成的最大容量 恩杜夫斯4 飞/飞 原代细胞在最佳底物条件下。 这些测量是在洋地黄素渗透的情况下获得的 恩杜夫斯4 飞/飞 原代细胞具有CI依赖性底物谷氨酸+苹果酸或丙酮酸+苹果酸酯,有或没有CI抑制剂鱼藤酮。 所有的比率都是相对于使用琥珀酸盐+鱼藤酮确定的CII依赖性比率进行的,因为这些比率不应受到CI缺乏的影响。 重要的是,在CII-依赖型ATP合成率的基因型之间没有检测到统计上的显著差异( 表1 ).
表1。 分离线粒体和渗透细胞中依赖CII的ATP合成速率 恩杜夫斯4 飞/飞 原代细胞系。 *使用底物琥珀酸盐和复合I抑制剂鱼藤酮±S测量ATP合成速率。 E.M.公司( n个 ). 未检测到具有统计学意义的差异( P(P) <0.050水平)介于+/+和 弗基 / 弗基 样品,通过未配对测量 t吨 测试,使用Holm–Sidak方法校正多重比较(GraphPad,PRISM V6.0b)。
ATP(μmol/g/min)*
原代细胞系 中等 准备 +/+ 飞/飞
MEF公司 葡萄糖 渗透细胞 36±5.1 (9) 37±5.0 (13)
半乳糖 可渗透细胞 26±4.1 (7) 19±4.6 (8)
葡萄糖 分离线粒体 61±12 (6) 65±11 (6)
星形细胞 葡萄糖 渗透细胞 61±6.1 (13) 68±7.6 (11)
半乳糖 渗透细胞 48±3.9 (7) 45±1.4 (5)
葡萄糖 分离线粒体 84±11 (3) 89±22 (4)
神经元 国家统计局 渗透细胞 24±1.7 (17) 29±2.3 (21)
在 恩杜夫斯4 飞/飞 主要MEF,CI依赖的ATP合成速率与葡萄糖培养基上的对照几乎没有区别( 图4 A和 4 E; 82–105%(+/+),但半乳糖降低( 图4 C类; +/-的55–60%)。 相反,在 恩杜夫斯4 飞/飞 谷氨酸+苹果酸降低了原代星形胶质细胞的CI依赖率( 图4 B、, 4 D和 4 F; 67–79%(葡萄糖+/+)和77%(半乳糖+/+),但丙酮酸+苹果酸正常,与培养基无关( 图4 B、, 4 D和 4 F) ●●●●。 在 恩杜夫斯4 飞/飞 所有CI依赖性底物都会降低初级神经元ATP合成速率( 图4 G; +/-的63–74%)。 为了证实底物的特异性,我们确定在CI抑制剂鱼藤酮存在的情况下,ATP合成的CI依赖速率几乎完全消失 恩杜夫斯4 飞/飞 所有培养条件下的原代细胞类型( 图4 A– 4 G) ●●●●。
图4。 ATP合成速率 恩杜夫斯4 飞/飞 细胞系,相对于用琥珀酸+鱼藤酮(S+R)测定的CII依赖率表达。
在渗透性细胞或从MEF分离的线粒体和葡萄糖维持的星形胶质细胞中测量ATP合成的CII-(S)和CI-依赖性(G+M,P+M)速率( A类 , B类 , E类 , F类 )或半乳糖( C类 , D类 )和神经元维持在NBM培养基上( 克 )来自+/+(白色条)和 恩杜夫斯4 飞/飞 (灰色条)老鼠。 如有指示,添加鱼藤酮以抑制CI。错误条=S.E.M.复制:S( n个 ≥3); G+M公司( n个 ≥3); G+M+R( n个 ≥2); P+M公司( n个 ≥3); 和P+M+R( n个 ≥2). P(P) 由双尾未配对生成的值 t吨 测验。 CI,复合物I; G、 谷氨酸; M、 苹果酸; P、 丙酮酸; R、 鱼藤酮; S、 琥珀酸盐。
我们还注意到渗透性初级神经元( 图4 G) 与渗透性MEF或星形胶质细胞相比,单用琥珀酸盐合成ATP的速率相对于琥珀酸+鱼藤酮的速率更低(<1)( 图4 A– 4 D) 。 此外,该相对比率在+/+比 恩杜夫斯4 飞/飞 神经元( 图4 G; 0.60对0.74)。 此外,在+/+MEF和星形胶质细胞分离的线粒体中观察到<1的相对比率,但没有 恩杜夫斯4 飞/飞 单元格( 图4 E和 4 F) ●●●●。 这些数据表明,RET(反向电子转移)可能发生在所有检查的原代细胞类型中。 术语RET描述了由CII产生的泛喹诺酮将电子提供给反向CI反应而不是正向CIII反应的过程[ 38 ]. RET被CI的鱼藤酮抑制所阻断。数据还表明,RET在 恩杜夫斯4 飞/飞 初级细胞,最突出的是神经元。 这可能是因为CI缺陷阻断了CI的正向和反向电子传递。此外,这些观察结果表明,在我们的渗透性细胞中,少量不存在于孤立线粒体中的残余CI依赖性底物可能会人为地增加ATP合成的CII依赖性速率。 事实上,当不提供底物时,虽然ATP合成的一般速率非常低(补充图S4A–S4G),但与分离的线粒体和神经元相比,渗透性细胞中的ATP合成速率略高。 作为对照,我们确定所有细胞类型中CII依赖性ATP合成的速率对CII抑制剂丙二酸敏感(补充图S4A–S4G;相对于琥珀酸+鱼藤酮的速率<1)。
活性氧生成速率在 恩杜夫斯4 飞/飞 主要MEF CI功能障碍不仅影响ΔΨ 米 以及相关的ATP合成,但也可能与ROS生成增加有关。 活性氧如果不被细胞迅速解毒,则有可能对细胞结构造成氧化损伤,包括DNA、蛋白质和脂质[ 39 , 40 ]. 因此,我们试图量化O的数量 2 •− 生产单位: 恩杜夫斯4 飞/飞 原代MEF和星形胶质细胞在葡萄糖或半乳糖培养基上培养,使用O 2 •− 特定探针DHE。 此外,CI和CII-依赖H 2 O(运行) 2 使用Amplex Red测量了在葡萄糖培养基上生长的MEFs和星形胶质细胞的分离线粒体的产量。由于样品可用性的限制,我们无法对O进行类似的测量 2 •− 和H 2 O(运行) 2 生产 恩杜夫斯4 飞/飞 初级神经元。
与对照细胞相比, 恩杜夫斯4 飞/飞 主要MEF产生的O量增加 2 •− 在葡萄糖培养基上休息( 图5 A; 150%+/+),但不在半乳糖培养基上( 图5 B) ●●●●。 相比之下,O的比率 2 •− 静态生产 恩杜夫斯4 飞/飞 原代星形胶质细胞在任一葡萄糖上培养( 图5 D) 或半乳糖( 图5 E) 培养基和对照组无法区分。 鱼藤酮治疗刺激O 2 •− 在所有细胞类型中,与基因型或培养条件无关( 图5 A、, 5 B、, 5 D和 5 E) ●●●●。 然而,鱼藤酮依赖性增加O 2 •− 两者的产量均较低 恩杜夫斯4 飞/飞 主要MEF( 图5 C) 和星形胶质细胞( 图5 F) 与+/+相比,不考虑文化媒体。
图5。 O(运行) 2 •− 和H 2 O(运行) 2 生产 恩杜夫斯4 飞/飞 细胞系。
O的速率 2 •− 生成量通过DHE的细胞积累进行测量,并与原始MEF和来自+/+(白条)和 恩杜夫斯4 飞/飞 (灰条)小鼠,用葡萄糖维持( A类 , D类 ),或半乳糖( B类 , E类 )中等。 O的速率 2 •− 如图所示,用CI抑制剂鱼藤酮调节产量。 O的比例 2 •− 从鱼藤酮处理细胞到非处理细胞(+鱼藤酮/-鱼藤酮)的产量在( C类 )和( F类 ). H的比率 2 O(运行) 2 用Amplex Red测量葡萄糖生长的初级MEF的分离线粒体和+/+和 恩杜夫斯4 飞/飞 老鼠( 克 , H(H) )结果显示为琥珀酸盐+鱼藤酮(左 年 -轴)或谷氨酸+苹果酸(右侧 年 -轴)。 为了保持统计能力,只对基因型进行了比较,但对琥珀酸盐与鱼藤酮的组间比较除外,因为这种差异特别有趣( 克 , H(H) ). 误差线=S.E.M。, n个 ≥3, P(P) 由双尾未配对生成的值 t吨 测验。
相反,H的比率 2 O(运行) 2 其产量相当于分离的线粒体中的对照 恩杜夫斯4 飞/飞 MEF公司( 图5 G) 和星形胶质细胞( 图5 H) 同时含有CI-(谷氨酸+苹果酸)和CII-依赖底物(琥珀酸)。 此外,鱼藤酮诱导H 2 O(运行) 2 与CI相关的产量在+/+和 恩杜夫斯4 飞/飞 原发性MEF和星形胶质细胞( 图5 G和 5 H) ●●●●。 但值得注意的是,H的CII依赖性比率 2 O(运行) 2 当与CI抑制剂鱼藤酮一起孵育时,MEF和星形胶质细胞线粒体中的产量均低于不孵育时。 此外,尽管不显著,但这一趋势在这两个领域似乎都更加明显 恩杜夫斯4 飞/飞 MEF和星形胶质细胞与对照组比较。 这进一步表明RET在我们的原代细胞中发生,并且该过程在 恩杜夫斯4 飞/飞 细胞。
恩杜夫斯4 飞/飞 初级MEF更容易因半乳糖进入细胞死亡
我们的结果表明线粒体功能的改变是细胞类型特异性的,因此在葡萄糖培养基上的标准培养条件下, 恩杜夫斯4 飞/飞 主MEF表现出减小的ΔΨ 米 ROS生成增加,而星形胶质细胞和神经元的CI依赖性ATP合成能力降低。 此外, 恩杜夫斯4 飞/飞 初级MEF(而非星形胶质细胞)对半乳糖的维持平均更敏感,ΔΨ的进一步去极化证明了这一点 米 ATP合成相应下降。 所有这些过程都有可能影响细胞死亡的启动[ 41 ]. 因此,我们检测了 恩杜夫斯4 飞/飞 原代细胞。 这些调查表明 恩杜夫斯4 飞/飞 主要MEF( 图6 A、, 6 C和 6 E) 但不是星形胶质细胞( 图6 B、, 6 D和 6 F) ,对细胞死亡更敏感(有关活细胞分析的统计数据,请参阅补充表S1)。 然而,这种对细胞死亡敏感性的增加仅见于 恩杜夫斯4 飞/飞 在应用急性应激源H后,长时间使用半乳糖维持原发性MEF 2 O(运行) 2 分析时死亡细胞的百分比不仅增加了( 图6 A; 1.8倍+/+),但进入坏死细胞死亡的活细胞百分比增加( 图6 E; 13次+/+)。 与…对比 恩杜夫斯4 飞/飞 主要MEF, 恩杜夫斯4 飞/飞 星形胶质细胞( 图6 B、, 6 D和 6 F) 在所有测试的培养条件下,与+/+相比,对细胞死亡的敏感性没有明显变化。
图6。 细胞死亡 恩杜夫斯4 飞/飞 细胞系。
来自+/+和 恩杜夫斯4 飞/飞 流式细胞术检测小鼠进入细胞死亡状态。 原发性MEF的死亡细胞与+/+(葡萄糖培养基上)的比率如下所示( A类 )和星形胶质细胞( B类 )在葡萄糖培养基上培养1天或半乳糖培养基上培育1天或4天,有或没有急性H 2 O(运行) 2 在分析之前处理6小时。 可行的单细胞初级MEF的百分比如下所示( C类 , E类 )和星形胶质细胞( D类 , F类 )在葡萄糖或半乳糖中培养1天或4天( E类 , F类 )或没有( C类 , D类 )急性H 2 O(运行) 2 分析前治疗6h。 误差线=S.E.M。, n个 ≥4, P(P) 由双尾未配对生成的值 t吨 测试使用Holm–Sidak方法(GraphPad,PRISM V6.0b)校正多重比较*= P(P) ≤0.050, **= P(P) ≤0.005. PI,碘化丙啶。
讨论 人们普遍认为,由于不同的细胞和组织类型使用不同的能量需求和代谢途径,OXPHOS突变可能导致细胞特异性效应[ 9 , 42 ]. 在这里,我们试图通过分析神经和非神经原代细胞类型的线粒体功能来进一步描述这一点 恩杜夫斯4 飞/飞 模拟LS方面的小鼠模型[ 12 ]. 同时,我们使用培养条件(半乳糖)强制 恩杜夫斯4 飞/飞 主要MEF和星形胶质细胞较少依赖糖酵解ATP生成,更多依赖OXPHOS相关ATP生成,以强调任何潜在的线粒体缺陷。 特别是星形胶质细胞已被证明对呼吸抑制有上调糖酵解的反应[ 43 , 44 ]. 这项研究显著地直接比较了来自OXPHOS缺乏症同一动物模型的原代细胞 恩杜夫斯4 飞/飞 鼠标。 此外,它还包括对线粒体CI缺乏哺乳动物模型中的星形胶质细胞进行的第一次综合分析。
恩杜夫斯4 飞/飞 MEF被选为对照非神经细胞类型,因为OXPHOS缺陷患者成纤维细胞是我们目前关于线粒体疾病细胞生物化学知识的来源[ 45 ]. 标准培养条件下的生化剖面 恩杜夫斯4 飞/飞 本研究中描述的主要MEF与在NDUFS4缺乏的人和小鼠成纤维细胞模型系统中进行的研究密切相关。 值得注意的是,CI活性降低,CI组装受损,ΔΨ 米 轻微去极化,ROS生成通常增加[ 15 – 17 , 46 , 47 ]. 这些数据表明,疾病的发病机制可能部分源于ΔΨ受损 米 对维持基本细胞功能至关重要,如ATP合成、线粒体蛋白导入、细胞钙 2+ 同型性,最终细胞存活。 此外,活性氧还与疾病进展有关,导致细胞结构氧化损伤。 然而,由于LS主要是一种神经系统疾病,因此成纤维细胞不适合研究神经病理学的机制。
然后我们发现 恩杜夫斯4 飞/飞 初级MEF和神经相关的星形胶质细胞和神经元不同。 也就是说,尽管有神经和非神经 恩杜夫斯4 飞/飞 初级细胞类型表现为CI活性降低和CI形成中断,对线粒体功能的影响不同。 特别地, 恩杜夫斯4 飞/飞 初级星形胶质细胞和神经元的ΔΨ非常正常 米 在标准培养条件下。 尽管如此,我们的数据表明,CI-依赖型ATP合成速率 恩杜夫斯4 飞/飞 原代星形胶质细胞和神经元减少,与之前的脑研究类似 恩杜夫斯4 飞/飞 小鼠还报告CI依赖性ATP合成减少(+/+的65%)[ 12 ]. 这代表了与 恩杜夫斯4 飞/飞 主要MEF,可能指ΔΨ的维护 米 作为自适应响应 恩杜夫斯4 飞/飞 神经细胞类型可以防止细胞死亡。 这种反应可能是由于心血管活动的调节,可能是通过改变IF的水平 1 调节蛋白[ 48 ]或复合体的反向作用[ 49 , 50 ]. 值得注意的是,虽然我们已经报道了ATP合成的最大速率,但ΔΨ 米 在稳态条件下进行监测。 此外,与其他类型的细胞相比,神经元本身可能对CI活性的破坏更为敏感,我们的结果表明,与星形胶质细胞和MEF相比,神经元相对于线粒体体积和/或数量而言,CI容量降低。
此外,虽然活性氧生成增加是NDUFS4缺陷患者和小鼠成纤维细胞的特征[ 17 , 46 ],在我们的 恩杜夫斯4 飞/飞 原代星形胶质细胞,也不是之前描述的NDUFS4缺陷中脑神经元[ 20 , 21 ]. 先前的细胞研究表明CI抑制程度与ROS生成水平之间存在相关性[ 18 , 51 ]. 相反,我们观察到来自同一小鼠模型的不同细胞类型与相同CI抑制水平之间ROS生成的差异。 这种差异可能部分归因于细胞抗氧化剂的不同水平,例如在神经元和星形胶质细胞中观察到的谷胱甘肽减少[ 52 , 53 ]. 这再次强调了在适当的细胞中研究这些参数的致病相关性。 同时,应该注意到 恩杜夫斯4 飞/飞 添加鱼藤酮后,初级MEF和星形胶质细胞的ROS生成量增加较小。 这可能反映了对NDUFS4丢失和观察到的不稳定CI的适应 在体外 在这些细胞中,CI-缺乏细胞中ROS生成的基本速率比+/+细胞更接近最大速率。
尽管如此,在大脑中观察到氧化损伤 恩杜夫斯4 KO小鼠[ 13 ]. 虽然氧化损伤可能是导致疾病的主要因素,但活性氧的来源可能是受影响细胞类型的外部因素。 卡瓦纳和哈丁[ 7 ]提示LS患者神经组织的氧化损伤是由pH值变化对神经血管系统的影响(高乳酸血症)介导的。 此外,由于代谢活动的高速度,这些pH值的变化在大脑的局部水平上可能更加明显[ 7 , 54 ]. 结合哺乳动物大脑的高氧环境,条件可能有助于催化活性氧的形成,从而介导受影响大脑区域的氧化损伤[ 7 ]. 另外,脑中的氧化损伤可能是由胶质增生引起的,胶质增生是包括星形胶质细胞在内的活化胶质细胞的浸润。 神经胶质增生是LS的一个特征[ 7 , 8 ],也在 恩杜夫斯4 KO小鼠[ 13 , 14 ]. 这些活化的胶质细胞可能释放促炎细胞因子,进一步向受影响的大脑区域招募免疫细胞,产生活性氧爆发并导致氧化损伤[ 7 , 13 , 14 , 55 ]. 此外,星形胶质细胞的异常激活可能通过促进炎症、形成疤痕和阻止神经元轴突再生而导致组织损伤[ 55 , 56 ]. 事实上,在线粒体疾病的神经相关模型中观察到的ROS生成和氧化损伤之间几乎没有什么例外[ 9 ].
这项研究还发现证据表明,来自泛喹诺酮(QH)的电子 2 )在CII处琥珀酸氧化生成的产物可以正向转移到CIII,反向转移到CI(RET)[ 38 , 57 ]. CII依赖型ATP合成速率的效率提高和H 2 O(运行) 2 当在CI抑制剂鱼藤酮的存在下测量时。 值得注意的是,神经元的这种趋势似乎比原代MEF或星形胶质细胞更为明显,并且在 恩杜夫斯4 飞/飞 原代细胞与对照组比较。 尽管RET诱导的氧化还原平衡变化或ROS生成可能影响细胞信号传导,但这种现象与疾病的生理相关性尚不清楚。 因此,令人感兴趣的是,神经元的RET比率高于其他细胞。 尽管如此,由于这些实验测量的是最大活动率,它们可能没有反映出 体内 系统的活动。 此外,与 体内 CI衬底可能影响通过CI的正向电子传输的环境。
当用半乳糖代替葡萄糖生长时,以迫使氧磷依赖性呼吸, 恩杜夫斯4 飞/飞 原代MEF再次表型偏离星形胶质细胞。 While期间 恩杜夫斯4 飞/飞 主MEF通过进一步去极化其ΔΨ来响应 米 CI依赖型ATP合成能力相应降低, 恩杜夫斯4 飞/飞 原代星形胶质细胞的反应与在标准培养条件下维持时没有差异。 此外, 恩杜夫斯4 飞/飞 半乳糖的初级MEF增加了细胞死亡的倾向,而 恩杜夫斯4 飞/飞 原代星形胶质细胞则不然。 这表明 恩杜夫斯4 飞/飞 原代MEFs没有足够的OXPHOS储备能力来适应对半乳糖的维持。 相反, 恩杜夫斯4 飞/飞 星形胶质细胞可能有更大的备用氧磷容量,或者能够更容易地降低其代谢需求,从而优先维持其ΔΨ 米 然而,这种差异仅在急性应用H时才显著 2 O(运行) 2 ,这可能再次表明了这两种细胞类型中抗氧化防御系统的潜在差异,而这两种类型的细胞在本研究中并未进行研究。
恩杜夫斯4 飞/飞 初级神经细胞类型可能优先维持一些线粒体过程,例如ΔΨ 米 过度ATP合成,以避免对细胞死亡敏感。 这与LS患者所描述的相对神经元保留现象相一致,其特征是保留细胞死亡局部区域的神经元[ 7 , 8 ]. 同样,在CI缺乏患者中观察到 mt-Nd5/6 ΔΨ 米 在休息状态下升高了40%[ 53 ]但应用CV抑制剂寡霉素后病情恶化。 此外,神经元还具有其他机制来逃避细胞死亡,例如对细胞色素的敏感性降低 c(c) -介导的细胞凋亡[ 58 ].
考虑到CI-依赖型ATP合成能力 恩杜夫斯4 飞/飞 星形胶质细胞和神经元大大减少,这些细胞中的能量依赖过程,例如产生动作电位的能力,可能会受到影响。 同样,这可能会影响星形胶质细胞循环神经元释放的谷氨酰胺和钾作为神经递质的能力,维持血脑屏障并传播动作电位[ 23 , 24 , 56 , 59 ]. CI缺乏对星形胶质细胞循环神经元释放的神经递质的能力的影响尤为重要,因为这一过程可能占大脑能量预算的很大一部分,并且可能对ATP缺乏过敏[ 54 ].
我们共同证明了这一点 恩杜夫斯4 飞/飞 与星形胶质细胞和神经元相比,初级MEF对CI缺乏的反应不同,这可能反映出MEF来源的组织的代谢活性低于大脑。 此外,在休息时对初级星形胶质细胞和神经元进行测量,尚待确定受刺激细胞中线粒体功能的强健程度。 最终,这些结果强调了研究相关细胞和组织疾病机制的必要性,因为对不太相关的样本(如成纤维细胞)的研究可能会产生误导。
致谢 我们衷心感谢Tomris Mustafa(Florey神经科学和心理健康研究所)对神经元和星形胶质细胞培养程序的帮助,以及Ian Trounce(澳大利亚眼科研究中心)和Doug Wallace(美国费城儿童医院线粒体和表观基因组医学中心) 为ρ°细胞的馈赠。 我们还感谢Matthew Burton(澳大利亚默多克儿童研究所)在执行和分析流式细胞术和活细胞成像收集的数据方面提供的帮助。
作者贡献 以下作者对本研究的概念和设计做出了贡献:Matthew J.Bird; 小南·W·维杰耶拉特纳; 贾斯珀·C·科门; Michael T.Ryan; David R.Thorburn; 和Ann E.Frazier。 数据的收集和/或汇编由Matthew J.Bird、Xiaonan W.Wijeyeratne、Adrienne Laskowski和Ann E.Frazier完成。 Matthew J.Bird、Xiaonan W.Wijeyeratne、Michael T.Ryan、David R.Thorburn和Ann E.Frazier对数据进行了分析和解释。 手稿由马修·J·伯德、贾斯珀·C·科曼、迈克尔·T·瑞恩、大卫·R·托本和安·E·弗雷泽撰写。
基金 作者的研究得到了澳大利亚国家卫生与医学研究委员会(NHMRC)的资助[资助号APP102222]和肌肉营养不良协会(MDA)的资助[资助号MDA113072],这是NHMRC对David Thorburn的主要研究奖学金[资助号1022896], Ann Frazier获得NHMRC职业发展奖[授予编号541920],Matthew Bird获得澳大利亚研究生奖[授予号码APA2011]。 澳大利亚线粒体疾病基金会和维多利亚政府的运营基础设施支持计划提供了额外支持。
工具书类
1. Skladal D.、Halliday J.、Thorburn D.R.儿童线粒体呼吸链疾病的最低出生流行率。 大脑。 2003; 126:1905–1912. doi:10.1093/brain/awg170。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
2 Munnich A.、Rustin P.线粒体疾病的临床谱和诊断。 美国医学遗传学杂志。 2001; 106:4–17. doi:10.1002/ajmg.1391。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
三。 Pfeffer G.,Chinnery P.F.线粒体肌病的诊断和治疗。 医学年鉴2013; 45:4–16. doi:10.3109/07853890.2011.605389。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
4 Koopman W.J.H.、Distelmaier F.、Smeitink J.A.M.、Willems P.H.G.M.OXPHOS突变和神经变性。 EMBO J.2013; 32:9–29. doi:10.1038/emboj.2012.300。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
5 DiMauro S.,Schon E.A.神经系统中的线粒体疾病。 每年。 神经科学版。 2008; 31:91–123. doi:10.1146/annurev.neuro.30.051606.094302。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
6 Baertling F.、Rodenburg R.J.、Schaper J.、Smeitink J.A.、Koopman W.J.H.、Mayatepek E.、Morava E.、Distelmaier F.《Leigh综合征诊断和治疗指南》。 神经学杂志。 神经外科。精神病学。 2014; 85:257–265. doi:10.1136/jnnp-2012-304426。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
7 Cavanagh J.B.,Harding B.N.利氏病病变的致病因素。 细胞能量剥夺的组织反应及其临床病理后果。 大脑。 1994; 117:1357–1376. doi:10.1093/brain/117.6.1357。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
8 Rahman S.、Blok R.B.、Dahl H.H.、Danks D.M.、Kirby D.M.,Chow C.W.、Christodoulou J.、Thorburn D.R.Leigh综合征:临床特征、生化和DNA异常。 安。神经。 1996; 39:343–351. doi:10.1002/ana.410390311。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
9 Bird M.J.、Thorburn D.R.、Frazier A.E.线粒体神经病理学生化特征建模。 生物化学。 生物物理学。 《学报》。 2014; 1840:1380–1392. doi:10.1016/j.bbagen.2013.10.017。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
10 Wallace D.C.,Fan W.小鼠线粒体疾病的病理生理学模型。 基因开发2009; 23:1714–1736. doi:10.1101/gad.1784909。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
11 Kruse S.E.、Watt W.C.、Marcinek D.J.、Kapur R.P.、Schenkman K.A.、Palmiter R.D.线粒体复合物I缺陷小鼠会发展成致命的脑肌病。 单元格元数据。 2008; 7:312–320. doi:10.1016/j.cmet.2008.02.004。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
12 Leong D.W.、Komen J.C.、Hewitt C.A.、Arnaud E.、McKenzie M.、Phipson B.、Bahlo M.、Laskowski A.、Kinkel S.A.、Davey G.M.等人。自发B2短穿插核元素(SINE)插入NADH脱氢酶(泛醌)生成的线粒体复合物I缺陷小鼠模型的蛋白质组学和代谢组学分析 Fe–S蛋白4(Ndufs4)基因。 生物学杂志。 化学。 2012; 287:20652–20663. doi:10.1074/jbc。 M111.327601。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
13 Quintana A.、Kruse S.E.、Kapur R.P.、Sanz E.、Palmiter R.D.Complex I因大脑中Ndufs4缺失而导致类似Leigh综合征的进行性脑病。 程序。 国家。 阿卡德。 科学。 2010年美国; 107:10996–11001. doi:10.1073/pnas.1006214107。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
14 Quintana A.、Zanella S.、Koch H.、Kruse S.E.、Lee D.、Ramirez J.M.、Palmiter R.D.Leigh综合征小鼠模型中的致命呼吸功能障碍。 临床杂志。 投资。 2012; 122:2359. doi:10.1172/JCI62923。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
15 Assouline Z.、Jambou M.、Rio M.、Bole-Feysot C.、de Lonlay P.、Barnerias C.、Desguerre I.、Bonnemains C.、Guillermet C.、Steffann J.等人。线粒体呼吸链复合物I的持续和类似组装缺陷允许快速识别Leigh综合征患者的NDUFS4突变。 生物化学。 生物物理学。 《学报》。 2012; 1822:1062–1069. doi:10.1016/j.bbadis.2012.01.013。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
16 Valsecchi F.、Monge C.、Forkink M.、de Groof A.J.C.、Benard G.、Rossignol R.、Swarts H.G.、van Emst de Vries S.E.、Rodenburg R.J.、Calvaruso M.A.等。NDUFS4基因缺失在永生化小鼠胚胎成纤维细胞中的代谢后果。 生物化学。 生物物理学。 《学报》。 2012; 1817:1925–1936. doi:10.1016/j.bbabio.2012.03.006。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
17 Valsecchi F.、Grefte S.、Roestenberg P.、Joosten-Wagenaars J.、Smeitink J.A.M.、Willems P.H.G.M.、Koopman W.J.H.NDUFS4−/−小鼠的初级成纤维细胞显示出ROS水平增加和线粒体形态异常。 线粒体。 2013; 13:436–443. doi:10.1016/j.mito.2012.12.001。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
18 Verkaart S.、Koopman W.J.H.、van Emst-de Vries S.E.、Nijtmans L.G.J.、van-den Heuvel L.W.P.J.,Smeitink J.A.M.、Willems P.H.G.M.超氧化物的产生与遗传性复合物I缺乏症的复合物I活性呈负相关。 生物化学。 生物物理学。 《学报》。 2007; 1772:373–381. doi:10.1016/j.bbadis.2006.12.009。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
19 Verkaart S.、Koopman W.J.H.、Cheek J.、van Emst-de Vries S.E.、van den Heuvel L.W.P.J.,Smeitink J.A.M.、Willems P.H.G.M.线粒体和细胞液硫醇氧化还原状态在孤立的人NADH中未检测到改变:泛醌氧化还原酶缺乏。 生物化学。 生物物理学。 《学报》。 2007; 1772:1041–1051. doi:10.1016/j.bbadis.2007.05.004。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
20 Choi W.S.、Kruse S.E.、Palmiter R.D.、Xia Z.鱼藤酮、MPP+或百草枯诱导的多巴胺能神经元死亡不需要线粒体复合物I抑制。 程序。 国家。 阿卡德。 科学。 美国2008年; 105:15136–15141. doi:10.1073/pnas.0807581105。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
21 Choi W.S.,Palmiter R.D.,Xia Z.帕金森病模型中线粒体复合物I活性的丧失会加剧微管功能障碍导致的多巴胺神经元死亡。 《细胞生物学杂志》。 2011; 192:873–882. doi:10.1083/jcb.201009132。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
22 Freeman M.R.星形胶质细胞的规格和形态发生。 科学。 2010; 330:774–778. doi:10.1126/science.1190928。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
23 Barres B.A.胶质细胞的神秘与神奇:对其在健康和疾病中作用的透视。 神经元。 2008; 60:430–440. doi:10.1016/j.neuron.2008.10.13。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
24 Dienel G.A.兴奋性神经传递期间的星形胶质细胞能量学:谷氨酸氧化和糖酵解的作用是什么? 神经化学。 国际2013年; 63:244–258. doi:10.1016/j.neuint.2013.06.015。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
25 杜峰,钱志明,朱磊,吴晓明,钱灿,陈荣,柯毅。不同培养方法培养的星形胶质细胞的纯度,细胞活性,GFAP和bystin的表达。 J.细胞。 生物化学。 2010; 109:30–37. doi:10.1002/jcb.22375。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
26 Meberg P.J.、Miller M.W.培养海马和皮层神经元。 方法细胞生物学。 2003; 71:111–127. doi:10.1016/S0091-679X(03)01007-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
27 Trounce I.A.、Schmiedel J.、Yen H.C.、Hosseini S.、Brown M.D.、Olson J.J.、Wallace D.C.通过与无mtDNA细胞的突触体融合在培养细胞中克隆神经元mtDNA变体。 核酸研究2000; 28:2164–2170. doi:10.1093/nar/28.10.2164。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
28 McKenzie M.、Lazarou M.、Thorburn D.R.、Ryan M.T.Barth综合征患者线粒体呼吸链超复合体不稳定。 分子生物学杂志。 2006; 361:462–469. doi:10.1016/j.jmb.2006.06.057。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
29 Frazier A.E.,Thorburn D.R.分光光度法电子传递链络合物的生化分析。 方法分子生物学。 2012; 837:49–62. doi:10.1007/978-1-61779-504-6。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
30 McKenzie M.、Lazarou M.、Thorburn D.R.、Ryan M.T.使用蓝色天然聚丙烯酰胺凝胶电泳分析线粒体亚单位组装成呼吸链复合物。 分析。 生物化学。 2007; 364:128–137. doi:10.1016/j.ab.2007.02.022。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
31 Dunning C.J.、McKenzie M.、Sugiana C.、Lazarou M.、Silke J.、Connelly A.、Fletcher J.M.、Kirby D.M.、Thorburn D.R.、Ryan M.T.人类CIA30参与线粒体复合体I的早期组装及其基因突变导致疾病。 EMBO J.2007; 26:3227–3237. doi:10.1038/sj.emboj.7601748。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
32 Wanders R.J.、Ruiter J.P.、Wijburg F.A.关于渗透性人皮肤成纤维细胞线粒体氧化磷酸化的研究:应用于线粒体脑肌病。 生物化学。 生物物理学。 《学报》。 1993; 1181:219–222. doi:10.1016/0925-4439(93)90024-U。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
33. Morten K.J.、Ackrell B.A.、Melov S.缺乏超氧化物歧化酶2小鼠的线粒体活性氧物种:通过抗氧化治疗的衰减。 生物学杂志。 化学。 2006; 281:3354–3359. doi:10.1074/jbc。 M509261200。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
34 Calvaruso M.A.、Willems P.、van den Brand M.、Valsecchi F.、Kruse S.、Palmiter R.、Smeitink J.、Nijtmans L.线粒体复合体III在不存在NDUFS4的情况下稳定复合体I以提供部分活性。 嗯,分子遗传学。 2012; 21:115–120. doi:10.1093/hmg/ddr446。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
35 Appleby R.D.、Porteous W.K.、Hughes G.、James A.M.、Shannon D.、Wei Y.-H.、Murphy M.P.缺乏线粒体DNA的人类细胞线粒体膜电位的定量和起源。 欧洲生物化学杂志。 1999; 262:108–116. doi:10.1046/j.1432-1327.1999.00350.x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
36 Buchet K.,Godinot C.维持人体线粒体DNA缺失ρ°细胞的生长和膜电位所必需的功能性F1-ATP酶。 生物学杂志。 化学。 1998; 273:22983–22989. doi:10.1074/jbc.273.36.22983。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
37. Boyer P.D.ATP合成酶——一个出色的分子机器。 每年。 生物化学评论。 1997; 66:717–749. doi:10.146/annrev.biochem.66.1.717。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
38 Dröse S.,Brandt U.线粒体呼吸链产生超氧化物的分子机制。 高级实验医学生物。 2012; 748:145–169. doi:10.1007/978-1-4614-3573-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
39. Balaban R.S.、Nemoto S.、Finkel T.线粒体、氧化剂和老化。 单元格。 2005; 120:483–495. doi:10.1016/j.cell.2005.02.001。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
40 Venditti P.、Di Stefano L.和Di Meo S.活性氧物种的线粒体代谢。 线粒体。 2013; 13:71–82. doi:10.1016/j.mito.2013.01.008。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
41 Galluzzi L.,Kepp O.,Kroemer G.线粒体:危险信号的主要调节因子。 自然修订版分子细胞生物学。 2012; 13:780–788. doi:10.1038/nrm3479。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
42 Lombès A.、AuréK.、Bellanné-Chantelot C.、Gilleron M.、Jardel C.与人类线粒体疾病相关的未解决问题。 生物化学。 2014; 100:171–176. doi:10.1016/j.biochi.2013.08.012。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
43 Bolaños J.P.、Almeida A.、Moncada S.糖酵解:生物能量还是生存途径? 生物化学趋势。 科学。 2010; 35:145–149. doi:10.1016/j.tibs.2009.10.006。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
44 Bouzier-Sore A.K.,Pellerin L.解开星形胶质细胞复杂代谢的本质。 前面。 单元格。 神经科学。 2013; 7:179. doi:10.3389/fncel.2013.00179。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
45 Saada A.线粒体:线粒体氧合酶(dys)在体外的功能——初级成纤维细胞的使用。 国际生物化学杂志。 细胞生物学。 2014; 48:60–65. doi:10.1016/j.bicel.2013.12.010。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
46 Distelmaier F.、Koopman W.J.H.、Van Den Heuvel L.P.、Rodenburg R.J.、Mayatepek E.、Willems P.H.G.M.、Smeitink J.A.M.线粒体复合物I缺陷:从细胞器功能障碍到临床疾病。 大脑。 2009; 132:833–842. doi:10.1093/brain/awp058。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
47 Lazarou M.、McKenzie M.、Ohtake A.、Thorburn D.R.、Ryan M.T.线粒体和细胞核DNA编码的亚基在复杂I.Mol.Cell中的组装谱分析。 生物学2007; 27:4228–4237. doi:10.1128/MCB.00074-07。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
48 Campanella M.、Casswell E.、Chong S.、Farah Z.、Wieckowski M.R.、Abramov A.Y.、Tinker A.、Duchen M.R.通过F1Fo-ATP酶抑制剂蛋白IF1调节线粒体结构和功能。 单元格元数据。 2008; 8:13–25. doi:10.1016/j.cmet.2008.06.001。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
49 McKenzie M.、Liolitsa D.、Akinshina N.、Campanella M.、Sisodiya S.、Hargreaves I.、Nirmalananthan N.、Sweeney M.G.、Abou-Sleiman P.M.、Wood N.W.等。与脑肌病和线粒体ATP消耗相关的线粒体ND5基因变异。 生物学杂志。 化学。 2007; 282:36845–36852. doi:10.1074/jbc。 M704158200。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
50 Chinopoulos C.,Adam-Vizi V.线粒体在细胞病理学中作为ATP消费者。 生物化学。 生物物理学。 《学报》。 2010; 1802:221–227. doi:10.1016/j.bbadis.2009.08.008。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
51 Barrientos A.,Moraes C.T.滴定线粒体复合物I损伤在细胞生理学中的作用。 生物学杂志。 化学。 1999; 274:16188–16197. doi:10.1074/jbc.274.23.16188。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
52 Dringen R.、Hirrlinger J.大脑中的谷胱甘肽途径。 生物化学。 2003; 384:505–516. doi:10.1515/BC.2003.059。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
53 Abramov A.Y.、Smulders-Sprinivasan T.K.、Kirby D.M.、Acin-Perez R.、Enriquez J.A.、Lightowlers R.N.、Duchen M.R.、Turnbull D.M.线粒体DNA突变神经元的神经退化机制。 大脑。 2010; 133:797–807. doi:10.1093/brain/awq015。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
54 Mergenthaler P.,Lindauer U.,Dienel G.A.,Meisel A.大脑糖:葡萄糖在生理和病理大脑功能中的作用。 《神经科学趋势》。 2013; 36:587–597. doi:10.1016/j.tins.2013.07.001。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
55 Singh S.、Swarnkar S.、Goswami P.、Nath C.星形胶质细胞和小胶质细胞:对神经病理学条件的反应。 国际神经科学杂志。 2011; 121:589–597. doi:10.3109/00207454.2011.598981。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
56 Sofroniew M.V.反应性星形胶质细胞在神经修复和保护中的作用。 神经科学家。 2005; 11:400–407. doi:10.1177/1073858405278321。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
57 Pryde K.R.,Hirst J.超氧化物是由线粒体复合体I中还原的黄素产生的:这是一种单一的、统一的机制,适用于正向和反向电子转移。 生物学杂志。 化学。 2011; 286:18056–18065. doi:10.1074/jbc。 M110.186841。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
58 Vaughn A.E.,Deshmukh M.葡萄糖代谢通过细胞色素c.Nat.Cell Biol的氧化还原失活抑制神经元和癌细胞的凋亡。 2008; 10:1477–1483. doi:10.1038/ncb1807。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
59 Walz W.星形胶质细胞在清除多余细胞外钾中的作用。 神经化学。 国际2000年; 36:291–300. doi:10.1016/S0197-0186(99)00137-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
关联数据 本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。