

摘要
介绍
GnRH,睾丸生理的历史调节剂
图1。
![]()
图2。
![]()
Kisspeptins,目前正在研究睾丸生理学的可能参与者
图3。
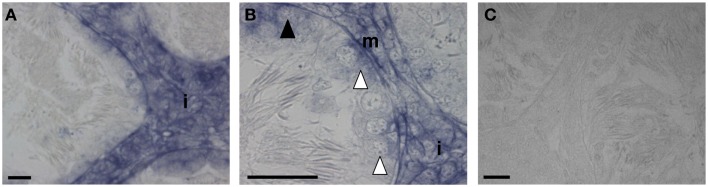
雌激素与精子质量
结束语
利益冲突声明
致谢
作者贡献
工具书类
-
1 Pierantoni R、Cobellis G、Meccarello R、Fasano S。 脊椎动物下丘脑-垂体-性腺轴细胞通讯的进化方面。 《细胞国际评论》(2002)218:69–14110.1016/S0074-7696(02)18012-0 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
2 Amoss M、Burgus R、Blackwell R、Vale W、Fellows R、Guillemin R。 绵羊源下丘脑促黄体激素释放因子(LRF)的纯化、氨基酸组成和N末端。 《生物化学与生物物理研究通讯》(1971)44:205–1010.1016/S0006-291X(71)80179-1 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
三。 Kah O、Lethimonier C、Somoza G、Guilgur LG、Vaillant C、Lareyre JJ。 后生动物中的GnRH和Gn RH受体:历史、比较和进化视角。 Gen Comp内分泌(2007)153:346–6410.1016/j.ygcen.2007.01.030 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
4 Kavanaugh SI、Nozaki M、Sower SA。 脊椎动物促性腺激素释放激素(GnRH)的起源:在基础脊椎动物海鳗中发现一种新的GnRH。 内分泌学(2008)149:3860–910.1210/en2008-0184 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
5 Millar RP、Lu ZL、Pawson AJ、加利福尼亚州弗拉纳根、Morgan K、Maudsley SR。 促性腺激素释放激素受体。 Endocr版本(2004)25:235–7510.1210/er.2003-0002 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
6 Morgan K,Millar注册会计师。 原脊索动物和脊椎动物中GnRH配体前体和GnRH-受体的进化。 Gen Comp内分泌(2004)139:191–710.1016/j.ygcen.2004.09.015 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
7 Oh DY、Song JA、Moon JS、Moen MJ、Kim JI、Kim K等。 促性腺激素释放激素受体(GnRHR)羧基末端的膜近端区域在哺乳动物和非哺乳动物GnRHRs之间传递差异信号。分子内分泌(2005)19:722–3110.1210/me.2004-0220 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
8 King JA,Millar注册会计师。 促性腺激素释放激素及其受体的进化。 细胞分子神经生物学(1995)15:5–2310.1007/BF02069556 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
9 吴HM、程JC、王和顺、黄海明、麦考曼CD、梁振英PC。 促性腺激素释放激素II型通过激活GADD45α诱导人子宫内膜癌细胞凋亡。 癌症研究(2009)69:4202–810.1158/0008-5472.CAN-08-4591 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
10 Aguilar-Rojas A,Huerta-Reyes M。 人促性腺激素释放激素受体激活垂体外组织和癌细胞的细胞功能和信号通路。 肿瘤代表(2009)22:981–9010.3892/或_00000525 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
11 Pawson AJ、Morgan K、Maudsley SR、Millar RP。 生殖生物学中的II型促性腺激素释放激素(GnRH-II)。 复制(2003)126:271–810.1530/rep.0.1260271 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
12 夏普RM。 睾丸的旁分泌控制。 《临床内分泌代谢》(1986)15:185–20710.1016/S0300-595X(86)80049-4 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
13 莫拉莱斯·P、皮萨罗·E、孔·M、克尔·B、塞里克·F、守夜·P。 促性腺激素释放激素刺激的精子与人类带状带结合是由钙内流介导的。 《生物报告》(2000)63:635–4210.1095/biolreprod63.2.635 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
14 Morales P、Pasten C、Pizzarro E。 抑制 体内 和 体内 促性腺激素释放激素拮抗剂对啮齿动物的受精作用。 《生物报告》(2002)67:1360–510.1095/biolreprod67.4.1360 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
15 Pierantoni R、Cobellis G、Meccarello R、Cacciola G、Chianese R、Chioccarelli T等。 脊椎动物睾丸促性腺激素释放激素活性、精子发生进展和精子运输。 Ann N Y科学院(2009)1163:279–9110.1111/j.1749-6632.2008.03617.x [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
16 Chianese R、Chioccarelli T、Cacciola G、Ciaramella V、Fasano S、Pierantoni R等。 低等脊椎动物模型在人类生殖研究中的贡献。 Gen Comp内分泌(2011)171:17–2710.1016/j.ygcen.2010.12.011 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
17 Bahk JY、Hyun JS、Chung SH、Lee H、Kim MO、Lee BH等。 GnRH mRNA在成熟大鼠和成人睾丸中表达和定位的阶段特异性鉴定。 《泌尿外科杂志》(1995)154:1958–6110.1097/00005392-199511000-00105 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
18 怀特RB,费尔纳德RD。 一个物种中三个促性腺激素释放激素基因的基因组结构和表达位点。 Gen Comp内分泌(1998)112:217–2510.1006/gcen.1998.7125 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
19 van Biljon W、Wykes S、Scherer S、Krawetz SA、Hapgood J。 人类精子中II型促性腺激素释放激素受体转录物。 《生物报告》(2002)67:1741–910.1095/biolreprod.101.002808 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
20 Millar R、Conklin D、Lofton Day C、Hutchinson E、Troskie B、Illing N等人。 一个新的人类GnRH受体同源基因:反义转录物的丰富而广泛的组织分布。 内分泌杂志(1999)162:117–2610.1677/joe.01620117 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
21 Pierantoni R、Fasano S、Di Matteo L、Minucci S、Varriale B、Chieffi G。 促性腺激素释放激素激动剂(buserelin)在 在体外 和 体内 青蛙产生睾酮( 天蛙 )睾丸。 分子细胞内分泌(1984)38:215–910.1016/0303-7207(84)90120-5 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
22 Ramakrishnappa N、Rajamahendran R、Lin YM、Leung PC。 非下丘脑生殖组织中的促性腺激素释放激素。 Anim Reprod Sci(2005)88:95–11310.1016/j.anireprosci.2005.05.009 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
23 Lin YM、Liu MY、Poon SL、Leu SF、Huang BM。 促性腺激素释放激素I和II刺激青春期前小鼠Leydig细胞的类固醇生成 在体外 《亚洲安德洛杂志》(2008)10:929–3610.1111/J.1745-7262.2008.00434.x [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
24 Fasano S、Pierantoni R、Minucci S、Di Matteo L、D’Antonio M、Chieffi G。 睾丸内注射雌二醇和促性腺激素释放激素(GnRHA,HOE 766)对完整和垂体切除患者血浆雄激素水平的影响 电鳐 和 短尾鱼雷 Gen Comp内分泌(1989)75:349–5410.1016/0016-6480(89)90169-X [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
25 King JA、Millar RP、Vallarino M、Pierantoni R。 肺鱼脑、性腺和血浆中促性腺激素释放激素的定位和特征, 环节原翅目 ). Regul Pept(1995)57:163–7410.1016/0167-0115(95)00025-7 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
26 Lin YM、Poon SL、Choi JH、Lin JS、Leung PC、Huang BM。 不育男性睾丸促性腺激素释放激素、类固醇生成酶和睾丸内睾酮水平的转录物。 费蒂尔·斯特尔(2008)90:1761–810.1016/j.fertnstert.2007.08.078 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
27 姚斌,刘海燕,顾永春,石石生,陶晓强,李晓杰,等。 促性腺激素释放激素通过细胞外信号调节激酶在大鼠Leydig细胞中正向调节类固醇生成。 《亚洲安德洛杂志》(2011)13:438–4510.1038/aja.2010.158 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
28 Anjum S、Krishna A、Sridaran R、Tsutsui K。 促性腺激素释放激素(GnRH)、促性腺素抑制激素(GnIH)、kisspeptin和GnRH-受体的定位及其在小鼠从出生到衰老的睾丸活动中的可能作用。 动物实验杂志(2012)317:630–4410.1002/jez.1765 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
29 Ubuka T、Son YL、Tobari J、Narihiro M、Bentley GE、Kriegsfelf LJ等人。 通过促性腺激素抑制激素及其受体对睾丸活动进行中央和直接调节。 前内分泌(2014)5:8.10.3389/fendo.2014.0008 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
30 郭JJ、马X、王春秋、葛玉芬、连庆群、哈迪·道等。 促黄体生成素和雄激素对大鼠Leydig祖细胞发育的影响 在体外 和 体内 《亚洲安德洛杂志》(2013)15:685–9110.1038/aja.2013.55 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
31 So WK、Cheng JC、Poon SL、Leung PC。 促性腺激素释放激素与卵巢癌:功能和机制概述。 FEBS期刊(2008)275:5496–51110.1111/J.1742-4658.2008.06679.x [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
32 White CD、Stewart AJ、Lu ZL、Millar RP、Morgan K。 促性腺激素释放激素激动剂的抗增殖作用:癌症治疗的前景和问题。 神经内分泌(2008)88:67–7910.1159/000119093 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
33 Andreu-Vieyra CV,滴定管公司,Habibi HR。 促性腺激素释放激素诱导金鱼睾丸细胞凋亡( 鲫鱼 ). 内分泌(2005)146:1588–9610.1210/en.2004-0818 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
34 Soverchia L、Carotti M、Andreu-Vieyra C、Mosconi G、Cannella N、Habibi H等人。 促性腺激素释放激素(GnRH)在金边鲷性腺分化调节中的作用( 海鲷 ). 摩尔-Reprod Dev(2007)74:57–6710.1002/日20484 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
35 Ogawa T、Dobrinski I、Avarbock MR、Brinster RL。 白脯氨酸是一种促性腺激素释放激素激动剂,可增强精原细胞移植到小鼠睾丸后的定植。 组织细胞(1998)30:583–810.1016/S0040-8166(98)80039-6 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
36 Ogawa T、Dobrinski I、Brinster RL。 受体准备对于大鼠精原细胞移植至关重要。 组织细胞(1999)31:461–7210.1054/tice.1999.0060 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
37 佐治亚州沙特尔斯沃思、德鲁伊DG、胡塔尼米一世、莱斯曼T、拉塞尔LD、谢蒂G等人。 促性腺激素释放激素拮抗剂对受照大鼠精原细胞增殖和分化的促进作用。 内分泌(2000)141:37–4910.1210/en.141.1.37 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
38 Treen N、Itoh N、Miura H、Kikuchi I、Ueda T、Takahashi KG等。 软体动物促性腺激素释放激素直接调节性腺功能:一种控制生殖的原始内分泌系统。 Gen Comp内分泌(2012)176:167–7210.1016/j.ygcen.2012.01.008 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
39 Minucci S、Di Matteo L、Pierantoni R、Varriale B、Rastogi RK、Chieffi G。 体内和 在体外 促性腺激素释放激素类似物(HOE 766)对青蛙精原细胞增殖的刺激作用, 天蛙 内分泌学(1986)119:731–610.1210/endo-119-2-731 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
40 Minucci S、Fasano S、Pierantoni R。 促性腺激素释放激素激动剂(buserelin)诱导青蛙进入S期, 埃斯库伦塔蛙 ,初级精原细胞。 Comp Biochem Physiol C药物毒性内分泌(1996)113:99–10210.1016/0742-8413(95)02046-2 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
41 Di Matteo L、Minucci S、Fasano S、Pierantoni R、Varriale B、Chieffi G。 促性腺激素释放激素(GnRH)拮抗剂降低青蛙雄激素生成和精原细胞增殖( 天蛙 ):垂体和睾丸中存在GnRH或GnRH-样物质受体的间接证据。 内分泌学(1988)122:62–710.1210/endo-122-1-62 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
42 Cobellis G、Meccarello R、Fienga G、Pierantoni R、Fasano S。 细胞质和核Fos蛋白形式调节青蛙精子发生的恢复, 天蛙 内分泌学(2002)143:163–7010.1210/en.143.1.163 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
43 Cobellis G、Meccarello R、Minucci S、Palmiero C、Pierantoni R、Fasano S。 青蛙体内Fos相关蛋白的细胞质与细胞核定位, 天蛙 ,睾丸: 体内 和直接 在体外 促性腺激素释放激素激动剂的作用。 Biol Reprod(2003)68:954–6010.1095/生物制品102.008938 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
44 Savulescu D、Feng J、Ping YS、Mai O、Boehm U、He B等。 促性腺激素释放激素调节抑制素介导促性腺细胞凋亡。 摩尔内分泌(2013)27:1856–7010.1210/me.2013-1210 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
45 Minucci S、Di Matteo L、Chieffi Baccari G、Pierantoni R。 促性腺激素释放激素类似物诱导完整和垂体切除青蛙的精子生成, 天蛙 《经验》(1989)45:1118–2110.1007/BF01950175 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
46 Chianese R、Ciaramella V、Scarpa D、Fasano S、Pierranoni R、Meccarello R。 阿南达胺调节 GnRH1号机组 , GnRH2型 和 促性腺激素释放激素受体 在青蛙睾丸中。 美国生理内分泌代谢杂志(2012)303:E475–8710.1152/ajpendo.0086.2012 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
47 Zerani M、Catone G、Quassinti L、Maccari E、Bramucci M、Gobbetti A等。 体外 促性腺激素释放激素(GnRH)对成年羊驼Leydig细胞的影响( 羊驼 )睾丸:GnRH受体免疫定位、睾酮和前列腺素合成以及环氧合酶活性。 Domest Anim内分泌(2011)40:51–910.1016/j.domaniend.2010.08.006 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
48 Chianese R、Ciaramella V、Fasano S、Pierantoni R、Meccariello R。 安奈达胺调节青蛙GnRH-II和GnRHRs的表达, 天蛙 间脑。 Gen Comp内分泌(2011)173:389–9510.1016/j.ygcen.2011.07.001 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
49 Cacciola G、Chianese R、Chioccarelli T、Ciaramella V、Fasano S、Pierantoni R等。 大麻与生殖:一段持久而有趣的历史。 制药(2010)3:3275–32310.3390/ph3103275 [ 内政部 ] [ 谷歌学者 ] -
50 Pierantoni R、Cobellis G、Meccarello R、Cacciola G、Chianese R、Chioccarelli T等。 雄性生殖中的CB1活性:哺乳动物和非哺乳动物动物模型。 维塔姆·霍姆(2009)81:367–8710.1016/S0083-6729(09)81014-5 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
51 Battista N、Meccarello R、Cobellis G、Fasano S、Di Tommaso M、Pirazzi V等人。 内源性大麻素在性腺功能和生育中沿进化轴的作用。 摩尔细胞内分泌(2012)355:1–1410.1016/j.mce.2012.01.014 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
52 Meccarello R、Battista N、Bradshaw HB、Wang H。 来自内源性大麻素系统的繁殖更新。 国际内分泌杂志(2014)2014(412354):16.10.1155/2014/412354 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
53 Scorticati C、Fernandez Solari J、De Laurentiis A、Mohn C、Prestifilipo JP、Lasaga M等人。 anandamide对促黄体生成素释放激素分泌的抑制作用被雌激素逆转。 美国国家科学院院刊(2004)32:11891–610.1073/pnas.0404366101 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
54 Farkas I、Kallo I、Deli L、Vida B、Hrabovszky E、Fekete C等。 逆行内源性大麻素信号减少GABA能突触向促性腺激素释放激素神经元的传递。 内分泌(2010)151:5818–2910.1210/en.2010-0638 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
55 Cacciola G、Chioccarelli T、Altucci L、Ledent C、Mason JI、Fasano S等。 CNR1基因敲除小鼠的低17-β-雌二醇水平通过干扰染色质重组影响精子细胞染色质重塑。 生物再现(2013)152:1–1210.1095/biolreprod.112.105726 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
56 Meccarello R、Franzoni MF、Chianese R、Cottone E、Scarpa D、Donna D等。 内源性大麻素系统与 GnRH-I型 在前脑哦,无尾两栖动物 天蛙 内分泌学(2008)149:2149–5810.1210/en.2007-1357 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
57 Chianese R、Cobellis G、Pierantoni R、Fasano S、Meccarello R。 非哺乳动物脊椎动物模型和内源性大麻素系统:与促性腺激素释放激素的关系。 摩尔细胞内分泌(2008)286:S46–5110.1016/j.mce.2008.01.09 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
58 Chianese R、Ciaramella V、Scarpa D、Fasano S、Pierranoni R、Meccarello R。 内源性大麻素和内源性香草素:睾丸GnRH信号调节的可能平衡。 国际内分泌杂志(2013)2013:904748.10.1155/2013/904748 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
59 Lee JH、Miele ME、Hicks DJ、Phillips KK、Trent JM、Weissman BE等人。 Kiss-1,一种新的人类恶性黑色素瘤转移抑制基因。 美国国家癌症研究所杂志(1996)88:1731–710.1093/jnci/88.23.1731 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
60 Kotani M、Detheux M、Vandenbogarede A、Communi D、Vanderwinden JM、Le Poul E等人。 转移抑制基因Kiss-1编码kisspeptin,kisspeptins是孤儿G蛋白偶联受体GPR54的天然配体。 生物化学杂志(2001)276:34631–610.1074/jbc。 M104847200 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
61 Messager S、Chatzidaki EE、Ma D、Hendrick AG、Zahn D、Dixon J等人。 基斯肽通过G蛋白偶联受体54直接刺激促性腺激素释放激素的释放。 美国国家科学院院刊(2005)102:1761–610.1073/pnas.0409330102 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
62 Seminara SB、Messager S、Chatzidaki EE、Thresher RR、Acierno JS、Shagory JK等人。 GPR54基因作为青春期调节器。 《英国医学杂志》(2003)349:1614–2710.1056/NEJMoa035322 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
63 de Roux N、Genin E、Carel JC、Matsuda F、Chaussain JL、Milgrom E。 KiSS1衍生肽受体GPR54功能丧失导致的促性腺激素减退。 美国国家科学院院刊(2003)100:10972–610.1073/pnas.1834399100 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
64 Beck BH、Fuller SA、Peatman E、McEntire ME、Darwish A、Freeman DW。 慢性外源性kisspeptin给药加速了Morone属低音的性腺发育。 Comp Biochem Physiol A Mol Integr Physiol(2012)162:265–7310.1016/j.cbpa.2012.03.019 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
65 Nocillado JN、Zohar Y、Biran J、Levavi-Sivan B、Elizur A。 慢性应用kisspeptin刺激青春期前雄性黄尾金鱼性腺发育( 拉氏师 ; perciformes)在繁殖和非繁殖季节。 Gen Comp内分泌(2013)191:168–7610.1016/j.ygcen.2013.06.005 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
66 Selvaraj S、Ohga H、Nyuji M、Kitano H、Nagano N、Yamaguchi A等。 皮下注射Kiss1十五肽促进青春期前雄性鲐鱼精子发生( 日本血吸虫 ). Comp Biochem Physiol A Mol Integr Physiol(2013年)166:228–3610.1016/j.cbpa.2013.06.007 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
67 Selvaraj S、Ohga H、Kitano H、Nyuji M、Yamaguchi A、Matsuyama M。 外周给药Kiss1十五肽可诱导性未成熟成鱼的性腺发育。 动物科学(2013)30:446–5410.2108/zsj.30.446 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
68 Saito H、Sawada T、Yaegashi T、Goto Y、Jin J、Sawai K等。 Kisspeptin-10刺激青春期前和青春期后雄性山羊体内黄体生成激素和睾酮的释放。 《动画科学杂志》(2012)83:487–9210.1111/J.1740-0929.2011.00978.x [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
69 考夫曼公司。 kisspeptin时代的成年:性别差异、发育和青春期。 分子细胞内分泌(2010)324:51–6310.1016/j.mce.2010.01.017 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
70 Demirbilek H、Gonc EN、Ozon A、Alikasifoglu A、Kandimir N。 女孩血清kisspeptin水平在中枢性性早熟诊断和青春期抑制评估中的评估。 《儿科内分泌代谢杂志》(2012)25:313–610.1515/jpem-2011-0445 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
71 Ratnasabapathy R,Dhillo WS。 kisspeptin在人类生殖功能治疗中的作用。 当前药物目标(2013)14:365–7110.2174/138945013804998981 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
72 Oakley AE、Clifton DK、Steiner RA。 大脑中的Kisspeptin信号。 Endocr版本(2009)30:713–4310.1210/er.2009-0005 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
73 Mayer C,Boehm U。 没有kisspeptin/GPR54信号的雌性生殖成熟。 《国家神经科学》(2011)14:704–1010.1038/nn.2818 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
74 Popa SM、Moriyama RM、Caligioni CS、Yang JJ、Cho CM、Concepcion TL等。 Kiss1表达中的冗余保护了鼠标中的复制。 内分泌(2013)154:2784–9410.1210/en.2013-1222 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
75 Ohtaki T、Shintani Y、Honda S、Matsumoto H、Hori A、Kanehashi K等。 转移抑制基因KiSS-1编码G蛋白偶联受体的肽配体。 《自然》(2001)411:613–710.1038/35079135 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
76 Funes S、Hedrick JA、Vassileva G、Markowitz L、Abbondanzo S、Golovko A等人。 KiSS-1受体GPR54对小鼠生殖系统的发育至关重要。 生物化学-生物物理研究公社(2003)312:1357–6310.1016/j.brc.2003.11.066 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
77 Terao Y、Kumano S、Takatsu Y、Hattori M、Nishimura A、Ohtaki T等人。 转移抑制基因KiSS-1在大鼠胎盘滋养层巨细胞中的表达。 Biochim Biophys Acta(2004)1678:102–1010.1016/j.bbaexp.2004.02.005 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
78 Tariq AR、Shahab M、Clarke IJ、Pereira A、Smith JT、Khan SH等。 猕猴睾丸中Kiss1和Kiss1受体的表达:睾丸功能的一种可能的局部调节器。 中欧生物杂志(2013)8:968–7410.2478/s11535-013-0219-4 [ 内政部 ] [ 谷歌学者 ] -
79 Irfan S、Ehmcke J、Wahab F、Shahab M、Schlatt S。 kisspeptin在恒河猴睾丸内的作用( 猕猴 ). 安德洛里亚(2013),10.1111/和.12121 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
80 Chianese R、Ciaramella V、Fasano S、Pierantoni R、Meccarello R。 Kisspeptin受体,GPR54,作为青蛙睾丸活动调节的候选者 天蛙 生物报告(2013)88:73.10.1095/biolreprod.112.103515 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
81 库雷希工业区拉姆赞F。 腹腔注射kisspeptin-10诱导成熟大鼠睾丸的剂量依赖性退行性改变。 生命科学(2011)88:246–5610.1016/j.lfs.2010.11.019 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
82 Thompson EL、Murphy KG、Patterson M、Bewick GA、Stamp GW、Curtis AE等。 慢性皮下注射kisspeptin-54会导致成年雄性大鼠睾丸退化。 美国生理内分泌代谢杂志(2006)291:E1074–8210.1152/ajpendo.00040.2006 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
83 Ramzan F、Qureshi IZ、Ramzan M、Ramzan MH、Ramzhan F。 kisspeptin-10治疗后,未成熟大鼠精囊出现组织形态学和超微结构改变。 Reprod Biol Endocrinol(2012)10:18.10.1186/1477-7827-10-18 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
84 Ramzan F、Qureshi IZ、Ramzan M、Ramzan MH、Ramzhan F。 Kisspeptin-10诱导青春期前大鼠前列腺的剂量依赖性变性。 前列腺(2013)73:690–910.1002/pros.22609 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
85 Mei H、Doran J、Kyle V、Yeo SH、Colledge WH。 kisspeptin信号在睾丸中有作用吗? 前内分泌(2013)4:198.10.3389/fendo.2013.0198 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
86 Shahab M、Mastronardi C、Seminara SB、Crowley WF、Ojeda SR、Plant TM。 下丘脑GPR54信号增强:灵长类青春期开始的潜在机制。 美国国家科学院院刊(2005)102:2129–3410.1073/pnas.0409822102 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
87 Roseweir AK、Kauffman AS、Smith JT、Guerriero KA、Morgan K、Pielecka-Fortuna J等人。 高效kisspeptin拮抗剂的发现揭示了促性腺激素调节的生理机制。 神经科学杂志(2009)29:3920–910.1523/JNEUROSCI.5740-08.2009 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
88 Pinto FM、Cejudo-Román A、Ravina CG、Fernández-Sánchez M、Martín-Lozano D、Illanes M等人。 人类精子中kisspeptin系统的特征。 国际安德洛尔杂志(2012)35:63–7310.1111/J.1365-2605.2011.717.x [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
89 许MC、王JY、李YJ、钟DS、徐凯、邱奇。 Kisspeptin调节小鼠精子的受精能力。 复制(2014).10.530/REP-13-0368 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
90 Pierantoni R、Cobellis G、Meccarello R、Palmiero C、Fienga G、Minucci S等人。 两栖类睾丸作为研究精子发生过程中生殖细胞进展的模型。 生物化学物理B生物化学分子生物学(2002)132:131–910.1016/S1096-4959(01)00543-7 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
91 Rastogi RK、Iela L、Saxena PK、Chieffi G。 绿蛙精子发生的控制, 天蛙 .J Exp Zool(1976)169:151–6610.1002/jez.1401960203 [ 内政部 ] [ 谷歌学者 ] -
92 Fasano S、Minucci S、Di Matteo L、D’Antonio M、Pierantoni R。 青蛙类固醇分布调节中的睾丸内反馈机制, 天蛙 《生殖内分泌》(1989)75:335–4210.1016/0016-6480(89)90167-6 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
93 Sirianni R、Chimento A、Ruggiero C、De Luca A、Lapano R和o S等人。 新型雌激素受体G蛋白偶联受体30介导17-β-雌二醇对小鼠精原细胞GC-1细胞株的增殖作用。 内分泌(2008)149:5043–5110.1210/en.2007-1593 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
94 Chimento A、Sirianni R、Casaburi I、Pezzi V。 雌激素受体(ER)和G蛋白偶联雌激素受体(GPER)在调节下丘脑垂体-睾丸轴和精子发生中的作用。 前内分泌(2014)5:1.10.3389/fendo.2014.0001 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
95 Lidke AK、Bannister S、Löwer AM、Apel DM、Podleschny M、Kollmann M等人。 17β-雌二醇在多毛类胚胎中诱导大量原始生殖细胞 环节蠕虫 Gen Comp内分泌(2014)196:52–6110.1016/j.ygcen.2013.11.017 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
96 Huma T、Ullah F、Hanif F、Rizak JD、Shahab M。 外周注射kisspeptin拮抗剂不会改变成年雄性恒河猴的基础血浆睾酮水平,但会降低血浆脂联素水平。 欧洲科学研究杂志(2013)109:668–7710.3906/双年度1401-53 [ 内政部 ] [ 谷歌学者 ] -
97 Carreau S、Bouraima-Lelong H、Delalande C。 男性生殖细胞中的雌激素。 精子发生(2011)1:90–410.4161/spmg.1.2.16766 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
98 Maggiolini M,Picard D。 GPR30是一种新的膜结合雌激素受体。 内分泌杂志(2010)204:105–1410.1677/JOE-09-0242 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
99 刘旭,朱鹏,沈KW,袁JM,谢C,张毅,等。 斑马鱼膜雌激素受体与哺乳动物GPR30同源性的鉴定及其在睾丸早期生殖细胞中的高表达。 《生物报告》(2009)80:1253–6110.1095/biolreprod.108.070250 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
100 Chimento A、Sirianni R、Delalande C、Silande D、Bois C和O S等人。 17β-雌二醇通过GPR30和ERα激活参与大鼠粗线期精母细胞凋亡的快速信号通路。 摩尔细胞内分泌学(2010)320:136–4410.1016/j.mce.2010.01.035 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
101 Alves MG、Socorro S、Silva J、Barros A、Sousa M、Cavaco JE等人。 体外培养的人支持细胞分泌大量的醋酸盐,醋酸盐由17β-雌二醇刺激并被胰岛素剥夺抑制。 Biochim Biophys Acta(2012)1823:1389–9410.1016/j.bbamcr.2012.06.002 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
102 拉戈五世、罗密欧五世、佐丹奴五世、马焦里尼五世、卡皮诺五世。 肿瘤和非肿瘤人类睾丸中雌激素受体GPER的鉴定。 生殖生物内分泌(2011)9:135.10.1186/1477-7827-9-135 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
103 O'Donnell L、Robertson KM、Jones ME、Simpson ER。 雌激素和精子发生。 Endocr Rev(2001)22:289–31810.1210/er.223.289 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
104 Shanbacker BD公司。 内源性雌激素对雄性绵羊黄体生成激素分泌的调节。 内分泌学(1974)115:944–5010.1210/endo-115-3-944 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
105 Eddy EM、Washburn TF、Bunch DO、Goulding EH、Gladen BC、Lubahn DB等。 雄性小鼠雌激素受体基因的靶向性破坏导致精子发生改变和不育。 内分泌(1996)137:4796–80510.1210/en.137.11.4796 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
106 Hess RA、Bunick D、Lee KH、Bahr J、Taylor JA、Korach KS等人。 雌激素在男性生殖系统中的作用。 《自然》(1997)390:509–1210.1038/37352 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
107 Miura T、Miura C、Ohta T、Nader MR、Todo T、Yamauchi K。 雌二醇-17β刺激男性精原干细胞的更新。 生物化学-生物物理研究公社(1999)264:230–410.1006/bbrc.1999.1494 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
108 Cobellis G、Pierantoni R、Minucci S、Pernas-Alonso R、Meccarello R、Fasano S。 c-Fos活性 天蛙 睾丸:季节性和雌二醇引起的变化。 内分泌(1999)140:3238–4410.1210/en.140.7.3238 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
109 Cobellis G、Lombardi M、Scarpa D、Izzo G、Fienga G、Meccariello R等。 青蛙中的Fra-1活性, 天蛙 ,睾丸。 Ann N Y Acad Sci(2005)1040:264–810.1196/annals.1327.039 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
110 Cobellis G、Lombardi M、Scarpa D、Izzo G、Fienga G、Meccariello R等。 青蛙中的Fra1活性, 天蛙 睾丸:精子运输中一个新的潜在作用。 《生物报告》(2005)72:1101–810.1095/biolreprod.104.036541 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
111 Cobellis G、Cacciola G、Chioccarelli T、Izzo G、Meccarello R、Pierantoni R等。 青蛙雄性生殖道的雌激素调节, 埃斯库伦塔蛙 :在肾小管周围肌样细胞的Fra-1激活和精子释放中发挥作用。 Gen Comp内分泌(2008)155:838–4610.1016/j.ygcen.2007.10.004 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
112 Lucas TF、Pimenta MT、Pisolato R、Lazari MF、Porto CS。 17β-雌二醇信号传导与支持细胞功能的调节。 精子发生(2011)1:318–2410.4161/spmg.1.4.18903 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
113 Ruwanpura SM,McLachlan RI,Meachem SJ。 雄性生殖细胞发育的激素调节。 内分泌杂志(2010)205:117–3110.1677/JOE-10-0025 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
114 Shetty G、Krishnamurthy H、Krishnamurthy HN、Bhatnagar S、Moudgal RN。 雌激素缺乏对雄性和雌性灵长类动物生殖生理的影响。 类固醇生物化学与分子生物学杂志(1997)61:157–6610.1016/S0960-0760(97)80008-8 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
115 Shetty G、Krishnamurthy H、Krishnamurthy HN、Bhatnagar AS、Moudgal NR。 芳香化酶抑制剂长期治疗对成年雄性帽猴睾丸功能的影响( 辐射支原体 ). 类固醇(1998)63:414–2010.1016/S0039-128X(98)00042-7 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
116 Staub C、Rauch M、Ferrière F、Trépos M、Dorval-Coiffec I、Saunders PT等。 睾丸引带雌激素受体ESR1及其46-kDa变异体的表达。 《生物报告》(2005)73:703–1210.1095/biolreprod.105.042796 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
117 Schulz RW、de França LR、Lareyre JJ、Le Gac F、Chiarini-Garcia H、Nobrega RH等。 鱼类精子发生。 Gen Comp内分泌(2010)165:390–41110.1016/j.ygcen.2009.02.013 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
118 Rolland AD、Lareyre JJ、Goupil AS、Montfort J、Ricordel MJ、EsquerréD等人。 虹鳟睾丸发育的表达谱确定了参与精子发生的进化保守基因。 BMC基因组学(2009)10:546.10.1186/1471-2164-10-546 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
119 Pinto PI、Teodósio HR、Galay-Burgos M、Power DM、Sweeney GE、Canário AV。 鲷鱼睾丸雌激素反应基因的鉴定( 金头鲷 )采用抑制消减杂交。 Mol Reprod Dev(2006)73:318–2910.1002/mrd.20402 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
120 Nitta H、Bunick D、Hess RA、Janulis L、Newton SC、Milette CF等。 小鼠睾丸生殖细胞表达P450芳香化酶。 内分泌(1993)132:1396–40110.1210/en.132.3.1396 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
121 Carreau S、Bouraima-Lelong H、Delalande C。 雌激素在精子发生中的作用。 Front Biosci(精英版)(2012)4:1–1110.2741/356 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
122 Tsutsumi I、Toppari J、Campeau JD、Di Zerega GS。 通过卵泡调节蛋白的系统治疗降低雄性大鼠的生育能力。 Fertil Steril(1987)47:689–95 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
123 Tsutsumi I、Fugimori K、Nakamura RM。 卵巢蛋白对大鼠生精上皮功能的破坏。 《生物报告》(1987)36:451–6110.1095/biolreprod36.2.451 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
124 D'Souza R、Gill-Sharma MK、Pathak S、Kedia N、Kumar R、Balasinor N。 睾丸内高雌激素对成年雄性大鼠生精上皮的影响。 摩尔细胞内分泌(2005)241:41–810.1016/j.mce.2005.04.011 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
125 Pentikäinen V、ErkkiläK、Suomalainen L、Parvinen M、Dunkel L。 雌二醇在体外人睾丸中作为生殖细胞存活因子。 临床内分泌代谢杂志(2000)85:2057–6710.1210/jcem.85.5.6600 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
126 Lanfranco F、Zirilli L、Baldi M、Pignatti E、Corneli G、Ghigo E等。 人类芳香化酶基因的一个新突变:雌激素替代治疗下成年男性血清雌二醇、纵向生长和骨密度之间关系的见解。 《骨骼》(2008)43:628–3510.1016/j.Bone.2008.05.011 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
127 Robertson KM、O'Donnell L、Jones ME、Meachem SJ、Boon WC、Fisher CR等。 缺乏功能性芳香化酶(cyp 19)基因的小鼠精子发生障碍。 美国国家科学院院刊(1999)96:7986–9110.1073/pnas.96.14.7986 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
128 Ebling FJ、Brooks AN、Cronin AS、Ford H、Kerr JB。 性腺机能减退小鼠精子发生的雌激素诱导。 内分泌学(2000)141:2861–910.1210/en.141.8.2861 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
129 Allan CM、Couse JF、Simanainen U、Spaliviero J、Jimenez M、Rodriguez K等人。 雌二醇诱导精子发生是通过雌激素受体-α机制介导的,该机制涉及刺激卵泡激素分泌的神经内分泌激活。 内分泌学(2010)151:2800–1010.1210/en.2009-1477 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
130 Baines H、Nwagwu MO、Hastie GR、Wiles RA、Mayhew TM、Ebling FJ。 雌二醇和FSH对性腺机能减退(hpg)小鼠睾丸成熟的影响。 生殖生物内分泌(2008)6:4.10.1186/1477-7827-6-4 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
131 Robertson KM、O'Donnell L、Simpson ER、Jones ME。 芳香化酶敲除小鼠的表型表明,饮食中的植物雌激素对睾丸功能有显著影响。 内分泌(2002)143:2913–2110.1210/en.143.8.2913 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
132 Shetty G、Weng CC、Bolden-Tiller OU、Huhtaniemi I、Handelsman DJ、Meistrich ML。 甲羟孕酮和雌二醇对受照大鼠精子发生恢复的影响。 内分泌(2004)145:4461–910.1210/en.2004-0440 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
133 周伟(Zhou W)、鲍登·蒂勒(Bolden-Tiller OU)、邵珊(Shao SH)、翁CC(Weng CC)、谢蒂·G(Shetty G)、阿布·埃利贾·M(AbuElhija M)等。 大鼠睾丸中雌激素调节基因及其与照射后精子发生恢复的关系。 《生物报告》(2011)85:823–3310.1095/biolreprod.111.091611 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
134 Porter KL、Shetty G、Shuttlesworth GA、Weng CC、Huhtaniemi I、Pakarin P等。 雌激素促进大鼠睾丸受辐射后精原细胞阻滞的恢复。 J Androl(2009)30:440–5110.2164/jandrol.108.006635 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
135 Chi H、Chun K、Son H、Kim J、Kim G、Roh S。 染料木素给药对白消安处理的大鼠睾丸精子发生恢复的影响。 临床实验与再生医学(2013)40:60–610.5653/学期2013.40.260 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
136 Ledent C、Valverde O、Cossu G、Petitet F、Aubert JF、Beslot F等。 CB1受体敲除小鼠对大麻素无反应,阿片类药物的成瘾效应降低。 《科学》(1999)283:401–410.1126/Science.283.540.401 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
137 Cobellis G、Cacciola G、Scarpa D、Meccarello R、Chianese R、Franzoni MF等。 青蛙和啮齿动物睾丸内大麻素系统:雄性生殖细胞中的1型大麻素受体和脂肪酸酰胺水解酶活性。 《生物报告》(2006)75:82–910.1095/biolreprod.106.051730 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
138 Cacciola G、Chioccarelli T、Ricci G、Meccarello R、Fasano S、Pierantoni R等。 脊椎动物雄性生殖中的内源性大麻素系统:比较概述。 分子细胞内分泌(2008)286:S24–3010.1016/j.mce.2008.01.004 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
139 Wenger T、Ledent C、Tramu G。 内源性大麻素anandamide激活CB1大麻素受体敲除小鼠的下丘脑-垂体-肾上腺轴。 神经内分泌(2003)78:294–30010.1159/000074882 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
140 Maccarrone M,Wenger T。 大麻素对下丘脑和生殖功能的影响。 Handb实验药理学(2005)168:555–7110.1007/3-540-26573-2_18 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
141 奥拉·M、米洛·h、温格·T。 内源性大麻素在调节黄体生成激素和催乳素释放中的作用。 AEA和2AG作用的差异。分子细胞内分泌(2008)286:S36–4010.1016/j.mce.2008.01.05 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
142 Grimaldi P、Orlando P、Di Siena S、Lolicato F、Petrosino S、Bisogno T等人。 内源性大麻素系统和CB2受体在小鼠精子发生中的关键作用。 美国国家科学院院刊(2009)106:11131–610.1073/pnas.0812789106 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
143 Fasano S、Meccarello R、Cobellis G、Chianese R、Cacciola G、Chioccarelli T等人。 内源性大麻素系统:一种参与男性生育控制的古老信号。 Ann N Y科学院(2009)1163:112–2410.1111/j.1749-6632.2009.04437.x [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
144 Cacciola G、Chiocarelli T、Mackie K、Meccariello R、Ledent C、Fasano S等人。 大鼠生后睾丸发育过程中1型大麻素受体的表达:可能参与成年Leydig细胞分化。 《生物报告》(2008)79:758–6510.1095/biolreprod.108.070128 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
145 Meccarello R、Chianese R、Cacciola G、Cobellis G、Pierantoni R、Fasano S。 青蛙中1型大麻素受体的表达, 天蛙 组织:可能参与睾丸活动的调节。 摩尔再现偏差(2006)73:551–810.1002/mrd.20434 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
146 温格·T、莱登特·C、塞尔努斯五世、格伦达·I。 中枢大麻素受体失活抑制内分泌生殖功能。 《生物化学与生物物理研究通讯》(2001)284:363–810.1006/bbrc.2001.4977 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
147 Chianese R、Ciaramella V、Fasano S、Pierantoni R、Meccarello R。 下丘脑-垂体轴:内源性大麻素抑制青蛙睾丸类固醇生成的强制性靶点。 Gen Comp内分泌学(2014).1016/j.ygcen.2014.02.010 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
148 Cobellis G、Ricci G、Cacciola G、Orlando P、Petrosino S、Cascio MG等。 2-花生酰甘油梯度调节小鼠附睾精子细胞的启动。 《生物报告》(2010)82:451–810.1095/biolreprod.109.079210 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
149 Ricci G、Cacciola G、Altucci L、Meccarello R、Pierantoni R、Fasano S等人。 内源性大麻素对精子运动的控制:附睾的作用。 Gen Comp内分泌(2007)153:320–210.1016/j.ygcen.2007.02.003 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
150 Cacciola G、Chioccarelli T、Altucci L、Viagjano A、Fasano S、Pierantoni R等。 细胞核大小作为小鼠精子雌激素反应染色质质量参数。 Gen Comp内分泌(2013)193:201–910.1016/j.ygcen.2013.07.018 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
151 Cacciola G、Chioccarelli T、Fasano S、Pierantoni R、Cobellis G。 雌激素与精子生成:来自1型大麻素受体敲除小鼠的新见解。 国际内分泌杂志(2013)2013:501350.10.1155/2013/501350 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
152 Lazaros LA、Xita NV、Kaponis AI、Zikopoulos KA、Plachouras NI、Georgiou IA。 雌激素受体α和β多态性与精液质量相关。 J Androl(2010)31:291–810.2164/jandrol.109.007542 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
153 Lambard S、Galeraud-Denis I、Bouraíma H、Bourguiba S、Chocat A、Carreau S。 芳香化酶在人类射精精子中的表达:一种推测的活动性标记。 Mol-Hum Reprod(2003)9:117–2410.1093/molehr/gag020 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
154 Lambard S、Galeraud-Denis I、Martin G、Levy R、Chocat A、Carreau S。 正常精子捐献者精液中信使核糖核酸的分析及其意义:与精子活力和获能的关系。 Mol-Hum Reprod(2004)10:535–4110.1093/molehr/gah064 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
155 Lambard S、Galeraud-Denis I、Saunders PT、Carreau S。 人类未成熟生殖细胞和射精精子含有芳香化酶和雌激素受体。 《分子内分泌杂志》(2004)32:279–8910.1677/jme.0.320279 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
156 赛义德·L、萨阿德·A、卡劳·S。 不育男性射精精子中芳香化酶mRNA的差异表达与弱精子症或畸形精子症的关系。 安德洛里亚(2014)46:136–4610.1111/和12058 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
157 Beck KJ、Herschel S、Hungershöfer R、Schwinger E。 类固醇激素对携带X和Y的人类精子运动和选择性迁移的影响。 费蒂尔·斯特里尔(1976)27:407–12 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
158 Idaomar M、Guerin JF、Lornage J、Czyba JC。 17β-雌二醇刺激弱精子症患者精子的运动和能量代谢。 阿奇·安德洛(1989)22:197–20210.3109/01485018908986772 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
159 Mattingly KA、Ivanova MM、Riggs KA、Wickramasinghe NS、Barch MJ、Klinge CM。 雌二醇刺激核呼吸因子-1的转录并增加线粒体的生物生成。 摩尔内分泌(2008)22:609–2210.1210/me.2007-0029 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
160 Adeoya Osiguwa SA、Markoulaki S、Pocock V、Milligan SR、Fraser LR。 17β-雌二醇和环境雌激素显著影响哺乳动物精子功能。 Hum Reprod(2003)18:100–710.1093/humrep/deg037 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
161 Ded L、Dostalova P、Dorosh A、Dvorakova-Hortova K、Peknicova J。 雌激素对公猪精子获能的影响 在体外 《生殖生物内分泌》(2010)8:87.10.1186/1477-7827-8-87 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
162 Carani C、Qin K、Simoni M、Faustini-Fustini M、Serpente S、Boyd J等。 睾酮和雌二醇对芳香化酶缺乏症患者的影响。 《英国医学杂志》(1997)337:91–510.1056/NEJM199707103370204 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
163 Herrmann BL、Saller B、Janssen OE、Gocke P、Bockisch A、Sperling H等人。 雌激素替代治疗对一名先天性芳香化酶缺乏症男性患者的影响,该患者由CYP19基因的新突变引起。 临床内分泌代谢杂志(2002)87:5476–8410.1210/jc.2002-020498 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
164 Maffei L、Murata Y、Rochira V、Tubert G、Aranda C、Vazquez M等人。 芳香化酶基因新突变男性的痛经综合征:睾酮、阿仑膦酸盐和雌二醇治疗的影响。 临床内分泌代谢杂志(2004)89:61–7010.1210/jc.2003-030313 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]