摘要 维持成人器官生发区的稳态是一个基本但在机制上知之甚少的过程。 尤其是,干细胞激活的控制因素尚不清楚。 我们之前已经证明,Notch信号限制成年斑马鱼大脑皮层神经干细胞(NSC)的增殖。 结合药理学和遗传操作,我们在此证明,长期Notch无效主要通过从静止状态激活NSC和增加对称分裂的发生来诱导NSC扩增。 表达分析、吗啉介导的失效和 缺口3 -空突变体直接与Notch3有关。 相反,废除 缺口1b 功能导致神经元的生成,但以激活的NSC状态为代价。 总之,我们的结果支持Notch受体在NSC招募的连续步骤中的不同参与。 他们暗示Notch3位于这一层次结构的顶部,以控制NSC的激活和扩增,保护成年NSC贮存器在生理条件下的体内平衡。
材料和方法 斑马鱼和 体内 操纵 所有动物实验均符合埃松部的官方监管标准(协议编号:A91-577 to L.B.C.)。 野生型AB斑马鱼品系(转基因品系)的3至9个月大或幼鱼 Tg(gfap:gfp) 2001年3月 ( Bernardos和Raymond,2006年 )(简称 gfap:gfp )和 缺口3 fh332(传真332) 使用了突变等位基因(见下文)。 为了使用一种胸苷类似物,将鱼保存在含有1 mM IdU、CldU或BrdU的池水中6小时。 为了应用多种类似物,将鱼在0.02%的三卡因中麻醉,并腹膜内注射等摩尔浓度的BrdU和EdU(10mM)(0.5μl/0.1g体重)。 受精后第五天(dpf)或第七天(7dpf)的幼仔在胚胎培养基(EM)中的10 mM BrdU、15%DMSO中浸泡20分钟,然后在EM中清洗 LY411575号 (重量/体积)( Fauq等人,2007年 )在28°C的游泳水中。 对于少于7天的治疗,每天更换LY溶液,对于较长的治疗,每周更换( 补充材料 图S3),没有效率损失(未显示)。 对照鱼用相同终浓度(0.04%)的二甲基亚砜载体处理。
为了选择性阻断Notch3或Notch1b的功能,我们对荧光素标记的剪接进行了电穿孔 缺口3 或 缺口1b 吗啉(MOs)(GeneTools,Philomath,OR,USA)注入成人大脑皮层的神经祖细胞:如前所述,将1.2 mM的MOs注入麻醉成人的脑室( Rothenaigner等人,2011年 )使用Intracel TSS20 ovodyne电穿孔器施加两个脉冲(70 V,50毫秒),在鱼头上方和下方的电极之间放置一个EP21电流放大器。 使用的MO( 缺口3 -MO,5′AAGGATCAGTCATCTTACCTTCGCTT3′; 缺口1b -MO,5′AATCTCAAACTGACCTGACCTCAAACCGAC3′)之前已经过验证( 马和江,2007 ; Milan等人,2006年 ). 以一种普通荧光标记MO的电穿孔作为对照(对照MO:5′CCTCTTACCTCACATTATA 3′)。
免疫组织化学 为了对成人大脑进行分析,将以下主要抗体应用于自由浮动的50μm振动体切片:GFP(1:500,Aves Laboratories)、MCM5(1:500;由德国海德堡马普医学研究所Soojin Ryu善意提供)、HuC/D(1:600,小鼠,Invitrogen;1:2000,人类,B博士的礼物。 Zalc,Salpétrière Hospital,Paris),BrdU(1:1000,小鼠克隆MoBU-1,Invitrogen;1:250,大鼠,Abcam),谷氨酰胺合成酶(1:500,小鼠,Millipore)和S100β(1:1000;兔,Dako)。 对于BrdU、IdU和CldU检测,将切片在37°C的2 M HCl中培养30分钟。 为了检测EdU,我们使用了Click-iT成像工具包(Invitrogen)。 使用山羊体内产生的与AlexaFluor染料(Invitrogen)偶联的二级抗体(1:1000)。 对于青少年的分析,冷冻切片的大脑用上述抗体进行免疫组织化学处理,此外还用抗BLBP(兔子,1:500,Millipore)和抗增殖细胞核抗原(小鼠抗PCNA,1:250,Santa Cruz Biotechnology)进行免疫组化处理。
现场 杂交
在65°C的温度下,在2 ng/μl DIG和荧光素标记的mRNA探针中孵育整个解剖的成人大脑18小时 缺口3、缺口1b、锯齿1a、锯齿1b 和 锯齿状2 (由英国伦敦研究所朱利安·刘易斯实验室提供的质粒)和DIG标记的 增量A,增量B 和 增量D 探针(由美国科罗拉多州奥罗拉科罗拉多大学布鲁斯·阿佩尔提供)。 接下来,振动切割横截面,并用抗DIG POD(绵羊,罗氏,1:500)或抗Fluo POD(羊,罗氏(1:500))孵育。 使用自制FITC和Cy3-共轭酪胺显示信号( http://www.xenbase.org/other/static/methods/FISH.jsp ). 对于单一染色,用抗DIG AP Fab片段(绵羊,罗氏,1:5000)培养切片,并用SIGMAFAST快速红/TR萘酚(Sigma)显示。
解剖的幼年大脑与DIG标记探针杂交,然后冷冻切片,与抗DIG-AP Fab片段(Roche)孵育,并用快速红处理。
图像采集和细胞计数 所有图像均在蔡司LSM700共聚焦显微镜上使用20×空气、40×油或63×油物镜拍摄。 使用ZEN软件(蔡司)处理图像。 测量( 补充材料 图S3)是使用ImageJ获得的。 对于图中的细胞计数 1 , 2 和 5 ,计数是在上手动执行的 z(z) -每种情况下,四个端脑50μm连续切片的堆栈。 对于 图7 每个青少年大脑中有三个非连续切片,每个实验中至少有五个大脑。
图1。
在Notch阻滞期间,对称分裂的放射状胶质细胞(RG)被招募到细胞周期中。 ( A类 )实验设计。 S期RG的初始队列标记为EdU脉冲,然后在Notch阻断(LY)期间标记为BrdU的第二个脉冲。 1d,间隔1天。 ( B类 )三联免疫染色显示苍白球Dm区GFAP-(绿色)、BrdU-(品红色)和EdU-(蓝色)阳性细胞(共焦投影分别超过16和19μm)。 ( C类 )EdU-阳性、BrdU-阳性和EdU/BrdU-阳性RG的量化。 红色条, P(P) <0.0001; 蓝色条, P(P) =0.07; 粉色条, P(P) >1 ( n个 =每个治疗组4个大脑,计数的细胞总数:1156)。 ( D-G公司 )如A所示,RG的分裂模式和实验环境中BrdU阳性双重体的量化。对称的胶质细胞分裂, P(P) <0.0001; 其他划分模式, P(P) >1; n个 =计数273倍。 比例尺:白色,20μm; 红色10μm。
图2。
缺口失效通过RG扩增触发大脑皮层GZ的持续过度生长。 ( A-E公司 )不同长度大脑皮层脑室区的横截面 LY411575号 RG标记GFAP-GFP(绿色)和增殖标记MCM5(品红色)染色。 ( F类 )LY治疗期间循环中RG的百分比(GFAP/MCM5阳性)。 ( G公司 )RG总数(GFAP阳性)。 ** P(P) <0.005; *** P(P) <0.0001( n个 =每种情况下4个大脑)。 比例尺:20μm。 四个光学平面(每个1μm)的共焦投影。
图5。
Notch3抑制解释了Notch阻断对RG激活的影响。 ( A-F公司 )成人端脑横截面上RG标志物谷氨酰胺合成酶(GS,绿色)、MCM5(品红色)和HuC/D(蓝色)的三重免疫组织化学 缺口3 +/+ 兄弟姐妹和 缺口3 fh332(传真332)/+ 控制条件下的杂合子(顶行)或LY处理下的杂合子(中行和底行)。 比例尺:20μm。 来自四个光学平面的共焦投影图像,每个平面厚1μm。 ( G公司 , 小时 )在不同基因型和处理条件下,以及在标准AB野生型系中,每段MCM5阳性祖细胞的总数(G)和RG细胞在增殖中的比例(H)。 P(P) <0.0001( n个 =3个大脑用于AB, 缺口3 +/+ 和 缺口3 fh332/+ )。 ( 我 )notch3-MO击倒实验示意图:在苍白脑室细胞中荧光标记的notch3-MO或对照-MO电穿孔(EP)后2天应用BrdU脉冲。 大脑立即进行分析。 ( J型 , K(K) )通过免疫细胞化学评估电穿孔(荧光素阳性)(绿色)RG(抗S100β)(品红色)的增殖状态(抗BrdU)(蓝色)分析。 箭头表示荧光标记的放射状胶质细胞BrdU阳性细胞。 比例尺:20μm。 来自四个光学平面的共焦投影图像,每个平面厚1μm。 ( L(左) )放射状胶质细胞MO靶向人群中BrdU阳性细胞的比例。 P(P) <0.001 ( n个 =每种情况下3个大脑)。
图7。
Notch3促进NSC静止,限制青少年大脑皮层的放大分裂。 ( A-D公司 )RG标记物BLBP(绿色)和增殖标记物PCNA(洋红)(灰色:DAPI)在5(C,D)和7(A,B)dpf处的苍白脑室区横截面 缺口3 fh332/fh332 纯合子突变体(B、D)和 缺口3 +/+ 兄弟姐妹(A、C)。 7 dpf处的白色箭头表示静止RG。 ( E-G公司 )在7 dpf(E)和5 dpf(F)条件下,每个切片的增殖细胞总数(PCNA)、RG(BLBP)和增殖RG(PCNA/BLBP),以及增殖RG的比例(G)。 ( H-K公司 )RG标记物BLBP或GS(绿色)和BrdU(洋红)(灰色:DAPI)在5(H,I)和7(J,K)dpf处的苍白脑室区横截面 缺口3 fh332/fh332 纯合子突变体(I,K)和 缺口3 +/+ 兄弟姐妹(H,J)。 7只dpf动物在5 dpf下用BrdU进行脉冲处理,并追逐2天。 白色箭头表示BrdU-阳性细胞在突变体中仍为RG。 ( L(左) , 米 )在BrdU脉冲(L)后5 dpf和追踪2天后(M),每段BrdU阳性细胞(BrdU)、RG(BLBP或GS)和BrdU正RG(BrdU/BLBP或BrdU/GS)的总数。 * P(P) <0.05; ** P(P) <0.005; *** P(P) <0.0001( n个 =两者均为3 缺口3 +/+ 和 缺口3 fh332/fh332 鱼类)。 比例尺:10μm。 来自四个光学平面的共焦投影图像,每个平面厚1μm。
统计 所有实验数据均表示为平均值±标准偏差。使用未配对双侧Student’s t吨 -检验,比较两组时的Fisher精确检验和单因素方差分析,然后是Bonferroni 事后(post-hoc) 在比较两组以上时进行测试。 重要性设置为 P(P) <0.05. 使用Excel软件和Statistica分析数据。
结果 Notch致残招募静止RG扩大胶质生成区 为了评估经典Notch信号对RG增殖的影响,我们使用γ-分泌酶抑制剂阻断Notch LY411575号 (简称LY)( Rothenaigner等人,2011年 ; Ninov等人,2012年 )应用于 gfap:gfp 转基因鱼( Bernardos和Raymond,2006年 )GFP忠实地标记苍白RG( Chapouton等人,2010年 ). 由此产生的增殖表型读出了γ-分泌酶抑制剂对Notch信号的特定作用( Chapouton等人,2010年 ). 这种治疗也会导致对 她 基因 her4.1(her4.1) RG中的表达( 补充材料 图S1A、B),确认 her4.1(her4.1) 在这个系统中是一个Notch目标,就像在胚胎中一样( Takke等人,1999年 ).
2天的缺口抑制增加了RG在大脑皮层生发池中的分裂比例( Chapouton等人,2010年 ). 这可能是由于加速治疗时已经处于增殖状态的RG的分裂(Notch作用于细胞分裂速度)、将静止RG招募到细胞周期(Notch影响RG激活)或两者兼而有之。 为了确定初始循环和静止RG对Notch阻断效应的相对贡献,我们给成年斑马鱼注射一次胸苷类似物EdU脉冲,以标记S期RG队列。 我们随后阻断了Notch,并用模拟BrdU应用第二个脉冲来标记在Notch阻断期间分裂的RG细胞( 图1A ). 选择EdU脉冲和LY治疗之间的时间足以使大多数EdU标记RG在Notch阻滞开始之前终止其细胞周期(未显示)。 然后用三重免疫标记法定量RG(GFP阳性)人群中EdU阳性、BrdU阳性和EdU/BrdU双标记细胞( 图1B、C ). 值得注意的是,我们观察到,在对照组和LY处理的条件下,每个切片中EdU阳性RG的数量,无论是仅标记EdU还是标记EdU和BrdU,都没有显著差异( 图1C 、蓝色和紫色条)。 EdU阳性细胞呈下降趋势; 然而,这并没有达到意义。 与对照组相比,在Notch阻断期间进入S期且仅BrdU阳性的RG数量显著增加( 图1C ,红色条)。 因此,Notch阻断诱导静止的RG进入细胞周期,对之前分裂的RG的细胞周期重新进入没有显著影响。
成年斑马鱼大脑皮层前体细胞可经历对称的胶质生成分裂,产生两个RG,对称的神经生成分裂产生两个神经元前体细胞(非RG前体细胞),不对称分裂产生一个RG和一个神经元前体内( Rothenaigner等人,2011年 ). 我们之前的克隆分析表明,Notch阻断增加了所有分裂事件的数量,但没有改变它们的相对比例( Rothenaigner等人,2011年 ). 然而,分析时间点发生在细胞标记后很久(4周),并在接受2天的LY治疗后。 因此,我们无法区分Notch阻断对细胞分裂模式的主要和次要影响,这些细胞分裂模式在谱系进展过程中可能是连续的。 为了解决这个问题,我们在 图1A 并在LY治疗结束时(即BrdU标记后1天)计数BrdU阳性(Notch-induced)细胞加倍,以强调Notch阻断的即时效果。 与M期后的姊妹细胞相对应的定量双倍体被定义为与BrdU共同标记的相邻细胞,并且与下一个相同标记的心室细胞之间的距离至少为四个细胞直径(25μM)。 我们观察到对称性胶质细胞分裂的数量显著增加( 图1D、G ),但不适用于其他划分模式( 图1E-G ),用于在Notch阻断期间新分裂的细胞。 我们得出的结论是,通过Notch阻断被募集到周期中的RG细胞主要经历对称的胶质细胞分裂。 因此,Notch信号抑制RG激活并特别限制其放大分裂。
缺口失效通过RG群体的快速扩增触发大脑皮层GZ的过度生长 缺口阻断2天导致60%RG中增殖标记的表达( Chapouton等人,2010年 ). 目前尚不清楚这一比例是否揭示了RG人群中的隐性异质性,即面对Notch封锁,合格RG与不合格RG之间的差异。 此外,RG激活是短暂的,然后是NSC耗竭还是减弱,类似于Notch阻断对小鼠的影响( Ables等人,2010年 ; Ehm等人,2010年 ; Imayoshi等人,2010年 ),未知。 为了研究这些方面,我们检查了较长的Notch失效的影响。
在对照鱼中,15%的RG表达增殖标记,如微染色体维持5(MCM5)( Ryu等人,2005年 ) ( 图2A、F ),与之前的报告一致( Chapouton等人,2010年 ; März等人,2010年 ). 我们验证了阻断Notch 2天可以激活60%RG中MCM5的表达( 图2B,F ). 治疗3天后,几乎所有RG都在增殖(95%的MCM5阳性RG)( 图2C、F )治疗5天和7天后,这一比例保持在最高水平( 图2D-F ). 因此,几乎所有RG都能够在3天内对Notch封锁作出反应,尽管一些RG的激活出现延迟。
经过几天的LY治疗,我们注意到沿着苍白脑室区出现了几个RG层( 图2D,E )这表明MCM5阳性RG比例的增加可能伴随着RG人群的扩大。 为了验证这一观察结果,我们对GFAP阳性细胞随时间的变化进行了量化。 在最初的2天内,每个切片的RG细胞总数保持在其初始值,但在整个治疗期间,该数量翻了两倍,即在第2天到第3天,然后是第5天到第7天( 图2G ). 这些发现表明,长期Notch无效后会发生两个不同的反应阶段:前3天招募全部RG人群进入周期,以及从第2天起扩大RG池的连续分裂事件。 此时,所有RG保持MCM5阳性( 图2E、F )这表明它们不会重新进入静止状态,并且容易再次分裂。 证实了这一点,在停止治疗后至少3天,再激活RG的分裂状态似乎稳定( 补充材料 图S2)。
为了挑战这种扩增的极限,我们将LY治疗的持续时间延长至5周( 补充材料 图S3)。 虽然我们观察到,经过3周的LY治疗后,RG的数量仍大幅增加,但这一点不如持续时间短的RG明显( 图2G )5周后趋于正常( 补充材料 图S3I)。 然而,与对照鱼形成鲜明对比的是,即使在连续5周的Notch阻断后,RG仍保持稳定增长的增殖率( 补充材料 图S3J)。 有趣的是,RG活性的这些长期改变并没有影响室上区的长度,但与之平行的是室下HuC/D阳性结构域的明显扩大( 补充材料 图S3K,L)。 这些观察结果突显了长期Notch阻断后RG扩增的某种程度的调节,其中大部分扩增的RG最终产生大量新生神经元。 然而,激活的RG层维持在心室位置,与正常NSC池相似,只是缺乏静止。
放大RG在单细胞水平显示NSC属性 为了首先确定LY扩增RG群体是否保持了NSC特性,我们分析了其自我更新和多潜能的能力。 使用7天的治疗模式,我们在Notch失效开始48小时后使用CldU标记增殖细胞( 补充材料 图S4A)。 此时,90%的CldU标记人群由Notch阻滞(未显示)导致的RG招募到细胞周期中。 在7天的LY治疗结束时,15%的CldU标记细胞仍在增殖RG(白色箭头 补充材料 图S4B),说明了分割RG状态的自我更新。 治疗结束三天后,这些细胞产生了不分裂的胶质细胞和分化的神经元( 补充材料 图S4C、D)。 因此,通过Notch阻断进入细胞周期的RG细胞群在长期Notch失效后也保持双功能特性。
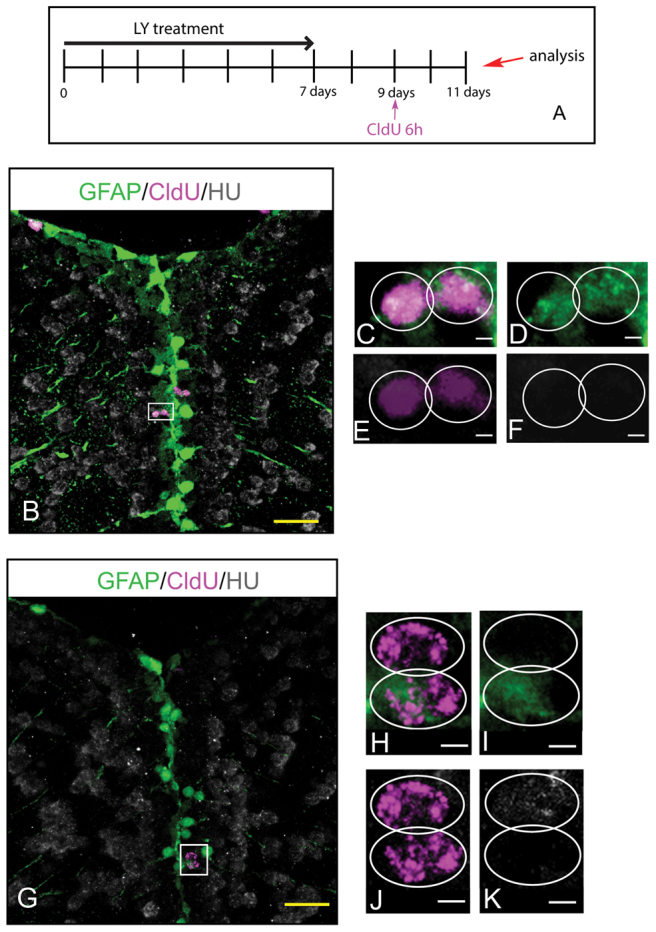
为了进一步研究扩增池中的单个RG是否表现为NSC,我们使用CldU介导的追踪追踪来自RG分裂的姐妹细胞的命运。 在为期7天的LY治疗结束后2天应用CldU(该阶段几乎所有标记细胞都属于扩增的RG池,未显示,请参阅 补充材料 图S2),并在两天追踪后监测CldU细胞加倍的身份( 图3A ). 沿着LY处理的鱼的心室区,我们观察到对称的CldU细胞双倍体对胶质标记物呈阳性反应,证明存在自我更新的对称性胶质细胞分裂( 图3B-F )以及不对称双峰,其中一个细胞表达胶质标记,另一个细胞表示神经元标记( 图3G-K ),突出单细胞水平的双电位。 这些结果表明,扩增池中的RG在单个细胞水平上保留了NSC特性,表明长期的Notch失效会导致大脑皮层生发池的扩张,除了缺乏静止外,还保持了NSC的特性。
图3。
单个RG细胞在Notch阻断后保持干细胞特性。 ( A类 )实验设计。 ( B-K公司 )大脑皮层脑室区的单光学切片 gfap:gfp 转基因大脑,GFP(RG,绿色)、CldU(品红色)和HuC/D(灰色)三重标记。 (C-F)B框中区域的高倍放大; 自我更新、对称性胶质细胞分裂的例子。 (H-K)G框中区域的高倍放大; 自我更新和不对称分裂的例子。 比例尺:黄色,20μm; 白色,2μm。
这些观察结果表明,当缺口水平恢复时,激活的RG状态可能是可逆的,从而挽救了GZ的大小和性质。 为了验证这一点,我们在中断10天的Notch阻滞1个月后分析了成年动物的大脑皮层脑室区。 GZ形态、RG数量和增殖RG的比例与对照鱼没有区别( 补充材料 图S5)。 我们的结论是,10天长的Notch无效并不会耗尽斑马鱼成年大脑皮层GZ中的神经干细胞。
缺口3 表达式,但不是 槽口1 ,是成人苍白RG的特征
几个 缺口 受体沿着成年斑马鱼皮层的GZ表达( Chapouton等人,2010年 ). 为了确定控制RG静止/激活周期的Notch受体,我们仔细分析了 缺口 基因与RG和增殖标记的比较( 图4 ). 静止和增殖的RG分别被定义为I型和II型细胞,而增殖的神经元前体被称为III型( März等人,2010年 ). 的表达式 缺口1a 似乎太弱,无法可靠地分配给特定的单元类型(未显示),以及 槽口2 未检测到表达。 然而,我们发现所有RG细胞都强烈表达 缺口3 RNA,无论其增殖状态如何,而 缺口3 III型细胞大多无表达( 图4A ). 相比之下 缺口1b MCM5阳性细胞的特征,无论是RG(II型)还是增殖性神经元前体(III型),但在静止的RG中不存在( 图4B ). 的微分表达式 缺口1b 和 缺口3 在苍白球-丘脑下边界周围尤为明显,RG和增殖的神经元前体细胞在空间上分离( 图4C ). 这些观察结果在很大程度上证明了 缺口1b 和 缺口3 基因( 图4D ),并表明RG激活的控制可能依赖于Notch3。 沿着这些线,我们发现Notch目标的表达 her4.1(her4.1) ( 补充材料 图S1A、B)主要表现为静止 缺口3 -RG阳性,尽管一些激活和 her4.1(her4.1) -可见阳性RG( 补充材料 图S1C,D)。 缺口配体的表达在GZ和紧邻的新生神经元中分布不同,可能形成局部生态位( 补充材料 图S6、S7)。
图4。
缺口3 和 缺口1b 在成人大脑皮层祖细胞中有差异表达。 ( A类 ) 缺口3 荧光显示的表达 就地 a的端脑横截面杂交(品红色) gfap:gfp 大脑,以及GFP(RG,灰色)和MCM5(绿色)的双荧光免疫染色。 (a′)大脑皮层背内侧(Dm)区, 缺口3 表达于静止(I型)(黄色星号)和增殖(II型)(黄箭头)RG。 III型细胞中没有转录物(白色箭头)。 (a′)丘脑-丘脑下交界区,富含III型细胞: 缺口3 表情很低或缺失。 ( B类 ) 缺口1b 表达(洋红色)与GFAP-GFP(RG,灰色)和MCM5(绿色)比较。 (b′,b′)Dm; 白色和黄色箭头表示 缺口1b -阳性的II型RG(b′)和III型祖细胞(b′。 静止RG(b′,黄色星号)中没有抄本。 (b′′)腭咽下交界处: 缺口1b 在III型细胞中强烈表达。 ( C类 )双荧光灯 就地 杂交 缺口3 (绿色)和 缺口1b (洋红色),MCM5免疫染色(蓝色)。 (c′)大脑皮层-丘脑下交界处高倍放大: 缺口3 由Dm的RG细胞表达 缺口1b 通过III型祖细胞。 ( D类 )的图形表示 缺口3 和 缺口1b 不同祖细胞类型中的表达(根据 März等人,2010年 ). 以Dm为单位, 缺口3 由97%的I型细胞表达( n个 =229个细胞计数),88%的II型细胞( n个 =31个细胞)和2%的III型细胞( n个 =23个单元格); 缺口1b 由2.8%的I型细胞表达( n个 =141个细胞),90%的II型细胞( n个 =43个细胞)和85%的III型细胞。 比例尺:白色,100μm; 品红,20μm。 单光学共焦平面,1μm。
Notch3活性门控成人大脑皮层神经干细胞激活 为了有选择地分析Notch3信号对NSC增殖的影响,我们应用TILLING策略生成一个 缺口3 突变等位基因( Draper等人,2004年 ). 我们恢复了 缺口3 fh332 它含有一个无义突变,在第8个Notch3 EGF重复序列的氨基酸669位引入了一个提前终止密码子。 这种突变预计会产生一种非功能性截短蛋白,该蛋白缺乏Notch3的大部分细胞外和所有跨膜及细胞内结构域。 缺口3 fh332/fh332 纯合子胚胎和幼虫发育速度正常,没有明显的形态缺陷(未显示),尽管这种突变在成年前是致命的。
缺口3 fh332(传真332)/+ 杂合子是成虫存活的,没有表现出任何明显的形态或行为异常(未显示)。 在Palium的细胞水平上,它们的心室区与野生型兄弟姐妹或标准AB系的野生型鱼类基本上没有区别,尽管增殖细胞的总数(MCM5阳性)比野生型水平略有但显著增加( 图5A、B、G ). 然而,增殖RG的比例保持正常( 图5H ). 作为评估Notch3介导的信号在成人RG静止中的相关性的第一种方法,我们测试了成人RG的敏感性 缺口3 fh332/+ 低剂量LY的杂合子。我们发现心室区增殖和大脑皮层增殖RG的比例在 缺口3 fh332(传真332)/+ 在野生型兄弟姐妹或AB鱼中未触发任何反应的剂量下的杂合子( 图5A-H ). 因此,Notch3信号的减少降低了RG对LY的敏感性阈值,表明Notch3至少部分介导了Notch阻断对RG激活的影响。
为了进一步确定Notch3在限制RG循环中的作用,我们通过电穿孔荧光标记的剪接吗啉(MO)靶向来阻断Notch3的功能 缺口3 ( 马和江,2007 )活鱼的大脑皮层脑室祖细胞。 电穿孔48小时后,我们通过测量S100β阳性细胞中BrdU掺入量来评估RG的激活状态( 图5I ). BrdU阳性RG的比例在 缺口3 -MO电穿孔(荧光标记)RG群体与使用对照MO电孔RG细胞的比较( 图5J-L ). 此外,获得的激活值(三倍)与RG对2天LY治疗的反应一致(参见 图2F ). 总之,我们得出结论,Notch3信号传导限制了成人大脑皮层静止RG的激活,是该细胞群LY抑制的主要靶点。
Notch3活性限制RG向扩增分裂激活 为了评估Notch3基因完全阻断对RG行为(包括分裂模式)的影响,我们分析了 缺口3 fh332/fh332 纯合子青少年。 青少年端脑的形态与成人类似,有一个位于外部的(“翻转”的)心室区( Folgueira等人,2012年 )如使用胶质标记物GS或BLBP(图 6 , 7 ).
图6。
缺口3 表达式,但不是 缺口1b 是斑马鱼幼鱼大脑皮层RG的特征。 ( A-D公司 )荧光灯 就地 杂交(品红色) 缺口3 (A、B)或 缺口1b (C,D)和RG标记物GS(绿色)(蓝色:DAPI)的免疫细胞化学。 比例尺:10μm。 来自四个光学平面的共焦投影图像,每个平面厚1μm。
在发育阶段,大脑皮层RG正在积极增殖( Dong等人,2012年 ). 我们发现,在内侧大脑皮层,RG从5dpf开始逐渐退出细胞周期,在7dpf时达到RG的25%左右(PCNA-阴性)(L.Dirian和I.Foucher,未发表;参见 图7D、E ). 在这些阶段, 缺口3 通常在位于中央的苍白球RG中强烈且特异表达( 图6A、B ). 相比之下, 缺口1b 在RG中几乎检测不到表达,总体而言,RG是位于腹侧的活跃增殖祖细胞的特征( 图6C、D ). 最后,Notch目标的表达式 her4.1(her4.1) RG从5 dpf开始下降 缺口3 fh332/fh332 纯合突变体( 补充材料 图S1E,F)。 这些观察结果确定Notch3是丘脑RG细胞中Notch信号的主要成分,并验证了其功能丧失特征 缺口3 fh332/fh332 突变体。
为了测试Notch3对RG激活和/或增殖的作用,我们分析了 缺口3 fh332/fh332 5和7dpf的纯合子( 图7A-D、F、G ),这是一个与大脑皮层RG的可测量比例的静止进入开始重叠的时期( 图7E ). 增殖RG的比例在 缺口3 fh332/fh332 在7dpf时纯合子与野生型同胞的比较( 图7E ):在突变体中,这一比例接近100%,表明RG无法退出循环或保持静止。 此外,RG的总数在 缺口3 fh332/fh332 两个阶段的纯合子( 图7F、G )这表明突变体中也发生了额外的RG生成分裂。 当它们对增殖祖细胞总数的影响变得明显时,这些分裂可能在5-7 dpf之间增加(PCNA-阳性)( 图7G ). 总的来说,这种表型与成人大脑皮层中Notch信号消失引起的表型惊人地相似( 图2 ). 为了验证它是否与成年RG中RG分裂扩增的增加有关,我们使用5 dpf的BrdU脉冲,然后进行2天追踪,以追踪成年RG分裂的RG细胞 缺口3 fh332/fh332 纯合子直至7 dpf( 图7H-M ). BrdU标记细胞在5dpf期的早期特征显示,在S期RG的数量正常 缺口3 fh332/fh332 纯合子( 图7H、I、L ). 然而,经过2天的追踪,突变株中BrdU阳性RG的数量显著增加( 图7J、K、M )表明心室中RG生成分裂的数量增加。 由于大多数在5 dpf时合并BrdU的祖细胞是RG( 图7L ,将BrdU与BrdU/BLBP进行比较),我们得出结论: 缺口3 fh332/fh332 在7dpf观察到的纯合表型是从5dpf开始扩增(对称)胶质母细胞分裂数量增加的结果。 总之,这些结果暗示Notch3在驱动RG静止以限制RG分裂的放大中起作用。
Notch1b信号是维持成人大脑皮层GZ中激活的祖细胞所必需的 的表达式 缺口1b 对增殖细胞(状态II和III)具有特异性,表明与Notch3相比,成人祖细胞具有不同的功能。 为了解决这一点,我们重复将荧光标记的MO电穿孔成活鱼的成年苍白球祖细胞,这一次使用针对 缺口1b ( Milan等人,2006年 ). 我们发现,阻断Notch1b活性并不影响电穿孔后2天RG在循环中的比例(MCM5阳性,GS阳性细胞),但可将该比例显著降低5天( 图8A-D ). 这种减少可能是由于II型细胞分裂后加速转化为静止的胶质细胞,和/或RG特性的丧失。 为了区分这些可能性,我们使用BrdU脉冲相分析追踪了Notch1b阻断后增殖祖细胞的命运( 图8E ). 电穿孔两天后,在MO靶向(荧光素阳性)人群中,S期RG细胞的比例(用BrdU脉冲标记)不受电穿孔的影响 缺口1b -生产任务单( 图8F-H ). 这与MCM5的表达一致( 图8D ),表明RG细胞的增殖参数在那一点上没有改变,BrdU掺入可用于追踪对照组和 缺口1b -耗尽的动物。 经过3天的追踪,RG人群中BrdU阳性细胞的比例在 缺口1b -MO与对照-MO细胞的比较( 图8H ). 因此,在Notch1b基因废除后,II型RG倾向于失去其胶质细胞特性。 在追踪的BrdU阳性人群中,III型祖细胞的比例在 缺口1b -MO电穿孔细胞,但相互转换影响II型细胞和神经元( 图8I ). 总之,这些观察结果表明,分裂RG在Notch1b废除后退出细胞周期,并且至少其中一些RG过早获得神经元命运。
图8。
Notch1b是维持祖细胞分裂和命运所必需的。 ( A类 )的示意图 缺口1b -MO击倒实验:在荧光标记物电穿孔后2天和5天对大脑进行分析 缺口1b -大脑皮层心室细胞中的MO或对照-MO。 ( B类 , C类 )电穿孔后5天通过免疫细胞化学评估的电穿孔(荧光素阳性)(绿色)RG(抗GS)(品红色)的增殖状态(抗MCM5)(蓝色)的分析。 箭头表示RG增殖,通常 缺口1b -MO-阴性。 ( D类 )MO靶向人群中MCM5阳性、GS阳性细胞的比例,2天( P(P) =0.05)和5天( ** P(P) <0.001)电穿孔后( n个 =每种情况下3个大脑)。 ( E类 )中的命运分析示意图 缺口1b -MO敲除实验:荧光素标记的notch1b MO或对照MO电穿孔后2天,在苍白心室细胞中施加BrdU脉冲。 大脑会立即或在3天的追踪后进行分析。 ( F类 , G公司 )电穿孔(荧光素阳性)(绿色,箭头)RG(抗GS)(品红色)中BrdU标记(白色)的分析,在3天追踪后通过免疫细胞化学进行评估。 箭头表示对照-MO(胶质细胞F)和 缺口1b -MO(G)。 ( 小时 )MO目标人群中2天内BrdU阳性、GS阳性细胞的比例( P(P) =0.05)和5天( ** P(P) <0.001)电穿孔后( n个 =每种情况下3个大脑)。 ( 我 )类型I(GS阳性,MCM5阴性)、类型II(GS正,MCM5+阳性)、类型III(GS阴性,MCM5-+阳性)和非前体细胞(GS负,MCM5-阴性)的比例[可能是神经元,它们实际上构成了从大脑皮层GZ生成的唯一非前体电池类型( Chapouton等人,2010年 ; Rothenaigner等人,2011年 )]电穿孔后5天,BrdU阳性MO靶向人群(II型,神经元: ** P(P) <0.001) ( n个 =每种情况下3个大脑)。 比例尺:10μm。 来自四个光学平面的共焦投影图像,每个光学平面的厚度为0.5μm。
讨论 成年脊椎动物脑中NSC贮存库的长期平衡依赖于对NSC干细胞的严格控制,而包括Notch在内的几个信号通路也参与了这一过程。 关于神经干细胞激活调控的机制,尤其是放大神经干细胞分裂的机制,以及它们对神经GZ长期内稳态的影响,知之甚少。 成年NSC具有很强的静息能力,我们之前已经证明,短暂的Notch失效会触发成年斑马鱼大脑皮层的心室增殖( Chapouton等人,2010年 )从长远来看,增加所有划分模式,同时保持其相对比例( Rothenaigner等人,2011年 ). 在此,结合药理学、遗传学和吗啉介导的失效与细胞分裂和细胞命运的追踪,我们证明:(1)Notch对NSC增殖的控制主要针对细胞周期从静止状态重新进入,从而激活NSC,并影响对称(但非不对称)RG分裂; (2) 长期药物性Notch废除导致成人大脑皮层NSC池的持续激活和短暂扩增,通过控制NSC静止将Notch确定为体内平衡的保护者; (3)在RG中选择性表达的Notch3是参与这一过程的关键受体,因为它的失效模拟了完全废除规范的Notch信号对RG静止和分裂模式的影响。 相比之下,我们发现在分裂祖细胞中表达的Notch1b直接或间接地需要维持祖细胞状态。 青少年大脑皮层RG增殖状态的动态演变不允许我们得出Notch3门静息退出还是加速静息进入的结论。 相比之下,大多数成年RG最初的静止状态清楚地表明,Notch3在此阶段限制了静止退出。 在成人身上,监测分裂后进入静止状态的BrdU追踪实验也无法支持Notch会加速进入静止状态(未显示)的模型。 总之,我们的结果确定Notch3是Notch受体,它限制NSC的激活以放大分裂,从而充当成人GZ内稳态的主要检查点。 我们首次在成年脊椎动物脑成熟NSC池中揭示Notch3的作用,这让人想起Notch3在控制肌肉卫星干细胞静止方面的已证明功能,突出了我们结论的一般相关性: 槽口3 -尽管肌肉形态正常,但缺陷小鼠的卫星细胞池扩大,活化的卫星细胞比例增加( 北本和花冈,2010年 ). 此外,通过损伤对该池的刺激导致其在 缺口3 -/- 小鼠,表明放大分裂的发生被夸大了( 北本和花冈,2010年 ). 基于这些结果和我们的研究,我们认为Notch3是成年生发种群生理条件下干细胞池大小的一个关键调节器,通过限制干细胞的激活以扩大分裂。
哺乳动物大脑中是否存在类似的Notch3介导机制尚待证实,但一些观察结果支持这一结论。 虽然此处分析的哺乳动物SEZ/SGZ和斑马鱼内侧大脑皮层不是直接同源的结构域( Salas等人,2006年 ; Mueller等人,2011年 ),它们的神经干细胞具有基本特征,如静止、神经胶质特性和基因表达( Adolf等人,2006年 ). 最近,基于RBPJk与Notch1相比失效的不同表型,在小鼠中假设了Notch1以外的Notch受体必须控制成年NSC增殖/激活( Basak等人,2012年 ). SEZ和SGZ成年神经祖细胞中RBPJk的条件性消融导致NSC的增殖短暂增加( Ehm等人,2010年 )在更大程度上,TAP人口( Ehm等人,2010年 ; Imayoshi等人,2010年 ). 相比之下,Notch1的条件性失活并不影响NSC的增殖率,但主要降低了增殖NSC的维持/自我更新和TAP的扩增( Ables等人,2010年 ; Basak等人,2012年 ). 正如我们在这里演示的那样,斑马鱼中Notch1b的失效会导致激活RG的丢失和神经元的生成。 虽然我们没有具体分析III型祖细胞的命运(被认为等同于TAP; März等人,2010年 ),而且我们目前还无法判断Notch1b是否主要控制细胞分裂或祖细胞状态本身,我们的结果直接平行于Notch1在小鼠中的已证明功能,并强化了类似分子过程控制斑马鱼和小鼠成体祖细胞行为的概念。 综上所述,我们的结果和可用的小鼠数据强烈表明,招募神经干细胞参与神经发生涉及到连续的Notch介导事件,这些事件显示了对Notch受体的不同需求:Notch3是级联中最上游的,控制着神经干细胞的初始激活率, 而Notch1在激活时参与,以允许分裂和/或维持干细胞/祖细胞的命运。 斑马鱼成年大脑皮层GZ在放大NSC分裂方面高度富集( Rothenaigner等人,2011年 )这使得这种分割模式在实验上特别容易实现,而非对称分割非常罕见。 对称的神经干细胞分裂虽然不常见,但可以从成年啮齿动物神经干细胞中观察到( Morshead等人,1998年 ). 同样重要的是,确定在这种情况下,从Notch(3)介导的静止释放时,分裂模式中是否有类似的选择性。
成人神经干细胞中Notch1和Notch3的差异活动是否与表达差异有关,或涉及真正不同的信号输出,仍有待直接评估。 大脑皮层RG中Notch1 ICD的异位表达降低了增殖,模拟了Notch3信号的预期效果( Chapouton等人,2010年 ). 然而,生理条件下NICD片段的表达可能绕过内源性特异性。 在小鼠中,Notch1和Notch3在NSC中共同表达( Basak等人,2012年 ),而 缺口1b 和 缺口3 斑马鱼神经干细胞仅部分重叠( 图4 ). 然而,斑马鱼Notch3和Notch1b的ICD片段缺乏抗体检测,无法精确定位这两种受体的活性。 Notch3和Notch1功能在肌肉卫星干细胞招募中的顺序意义( Conboy和Rando,2002年 )至少部分涉及两种受体的不同靶点( 北本和花冈,2010年 ),证实了先前的研究表明Notch1和Notch3信号的靶序列选择性( Ong等人,2006年 ). 最近观察到Notch信号的阈值水平用于控制斑马鱼胰腺祖细胞和再生脊髓中的特定细胞输出( Ninov等人,2012年 ; Dias等人,2012年 ),并且Notch1和Notch3的信号传导效率的定量差异也可能在成人脑NSCs中编码不同的细胞反应。
基于一致的表达式 her4.1(her4.1) 和胶质细胞标记物,有人认为通过Her4的Notch信号主要参与维持成人大脑中的胶质细胞特性( Ganz等人,2010年 ). 我们的结果强调了胶质标记物在 缺口3 长期Notch阻断后的表达域 缺口3 fh332(传真332) 变种人反对这种解释。 相反,Her4可能作为Notch3的效应物参与预防NSC激活。 鉴于 her4(her4) 然而,在幼年期RG增殖中,也有可能另一个Notch3靶点控制NSC的激活。 培养的成纤维细胞的轮廓数据表明Hes1是一种能直接对抗衰老和分化程序的静止因子,并被Notch上调( Coller等人,2006年 ; Sang等人,2008年 ; Sang和Coller,2009年 ). Hes1是否也直接影响扩散尚不清楚。 直接评估这些不同的 他/她 候选人 缺口3 废除,以及他们在控制国家安全委员会平静方面的个人职能。
我们的观察结果与小鼠RBPJk的基因无效性存在明显差异:在条件性RBPJk基因敲除小鼠中,NSC增殖是短暂的,NSC池最终耗尽( Ehm等人,2010年 ; Imayoshi等人,2010年 )而长期的Notch阻断导致斑马鱼成年大脑皮层GZ扩增( 图2 ). 延长LY治疗后,大量扩增RG的神经元分化; 然而,由活跃增殖的RG组成的组织学正常GZ保持不变,没有NSC丢失的迹象( 图3 ; 补充材料 图S4、S5)。 对这些看似不同的老鼠和斑马鱼表型有几种解释。 RBPJk突变小鼠的神经干细胞耗竭可能是由于神经干细胞的耗竭,或是在经过有限数量的分裂后进入非激活状态,这在突变小鼠中可能会过早达到。 据报道,衰老小鼠体内存在终止谱系( Encina等人,2011年 ),使这个假设成为可能。 例如,与小鼠相比,斑马鱼大脑皮层表现出更高的修复能力(由 克孜尔等人,2012年 )这可能意味着可能会有更多的NSC部门。 然而,还没有进行连续的损伤来反复挑战生发池,并且尚不清楚修复过程中主要招募哪些祖细胞亚型和分裂模式。 或者,或者另外,RBPJk-null小鼠中NSC的缺失可能是由于在没有Notch1的情况下分裂时干细胞的缺陷性维持所致( Ables等人,2010年 ; Basak等人,2012年 ). 我们的结果强调了斑马鱼Notch1b的类似功能,尽管Notch3对RG激活的抑制作用有所缓解,但LY对其的废除最终应导致NSC丢失。 我们认为,LY不太可能优先靶向斑马鱼Notch3而非Notch1b:LY的抑制活性在胚胎神经发生期间被证明是完全有效的(未显示),这在很大程度上依赖于Notch1功能。 与N3ICD相比,N1bICD具有更高的稳定性,这可能会导致Notch1b废除的延迟,考虑到 缺口1b -MO在5天内影响RG,而即使在LY治疗3周后,我们仍然观察到RG扩增( 补充材料 图S4)。 更令人信服的是,我们的LY治疗可能允许阈下Notch信号传导,这与斑马鱼成年大脑皮层GZ的大量NSC富集相结合,导致净NSC扩增反应。 此外,除Notch1b外的其他因子可能会部分补偿斑马鱼在LY治疗下保持茎干,和/或激活RG中Notch1b和Notch3的缺失可能会触发一种不同于仅Notch1b1缺失的表型。 沿着这些思路,Notch3失效的长期影响,尤其是该法规对NSC干细胞的影响,仍然是一个重要问题,应澄清NSC分割率和干细胞维持之间的联系。
总之,我们建议,维持成熟大脑中组成性神经原性NSC池的内稳态需要连续的Notch依赖性步骤,其中最上游的NSC激活和细胞周期重新进入由Notch3介导,而随后激活的NSC的维持需要Notch1。 这种分子机制与肌肉卫星干细胞的激活和卫星干细胞池大小的生理维持有关,但与表现出高细胞更新的系统,如造血、皮肤表皮或肠干细胞无关(由 Perdigoto和Bardin,2013年 ). 这种区别的一般相关性,以及Notch3与Notch1b活性的介导物,将在未来的研究中很重要。
致谢 我们感谢实验室同事的科学和技术投入,并感谢马里恩·库伦、伊莎贝拉·福彻、肖娜·卡茨和伊曼纽尔·丹·特荣对手稿的批判性阅读。 塞巴斯蒂安·贝杜提供了专业的鱼类护理。
脚注
基金
L.B.-C.实验室的工作得到了欧盟项目NeuroXsys[FP7/2007-2013,拨款协议编号223262]和ZF-Health[FP7/2010-2015,拨款协议号242048)的资助;以及法国国家科学院(ANR)[ANR-08-CEX-08-000-01和ANR-2012-BSV4-0004-01]、法国巴黎神经科学院(ENP)的资助 和La Fondation pour La Recherche Médicale(FRM)【FRP‘装备’DEQ20120323692】。 这个 缺口3 fh332(传真332) 突变等位基因是在美国国立卫生研究院的支持下产生的[R01 HG002995 to C.B.M.]。 存放在PMC中,12个月后发布。
作者贡献
A.A.、M.K.和L.B.-C.构思了这项研究并整理了手稿; A.A.和M.K.对成年人进行了实验; A.B.对青少年进行实验; S.G.分析 her4(her4) 成人的表达和增殖; L.P.和C.B.M.生成了 缺口3fh332 等位基因。
工具书类
Ables J.L.、Decarolis N.A.、Johnson M.A.、Rivera P.D.、Gao Z.、Cooper D.C.、Radtke F.、Hsieh J.、Eisch A.J.(2010)。 Notch1是维持成年海马干细胞库所必需的。 《神经科学杂志》。 30, 10484–10492 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Ables J.L.、Breunig J.J.、Eisch A.J.、Rakic P.(2011年)。 不仅仅是发育:成人大脑中的Notch信号。 神经科学自然评论。 12, 269–283 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Adolf B.、Chapouton P.、Lam C.S.、Topp S.、Tannhäuser B.、Strähle U.、Götz M.、Bally-Cuif L.(2006)。 斑马鱼端脑成体神经发生的保存和获得特征。 开发生物。 295, 278–293 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Andreu-AgullóC.、Morante-Redolat J.M.、Delgado A.C.、Fariñas I.(2009年)。 血管生态因子PEDF调节成人室管膜下区的Notch依赖性干细胞。 自然神经科学。 12, 1514–1523 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Basak O.、Giachino C.、Fiorini E.、Macdonald H.R.、Taylor V.(2012)。 神经源性室下区干/祖细胞在其活动状态而非静止状态下是Notch1依赖性的。 《神经科学杂志》。 32, 5654–5666 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Bernardos R.L.,Raymond P.A.(2006年)。 GFAP转基因斑马鱼。 基因表达。 模式 6, 1007–1013 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Bonaguidi M.A.、Wheeler M.A.、Shapiro J.S.、Stadel R.P.、Sun G.J.、Ming G.L.、Song H.(2011)。 体内克隆分析揭示了成体神经干细胞的自我更新和多能特性。 单元格 145、1142–1155 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Breunig J.J.、Silbereis J.、Vaccarino F.M.、Sestan N.、Rakic P.(2007年)。 Notch调节出生后齿状回新生神经元的细胞命运和树突形态。 程序。 国家。 阿卡德。 科学。 美国 104, 20558–20563 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Chapouton P.、Jagasia R.、Bally-Cuif L.(2007年)。 非哺乳动物脊椎动物的成年神经发生。 生物论文 29, 745–757 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Chapouton P.、Skupien P.、Hesl B.、Coolen M.、Moore J.C.、Madelaine R.、Kremmer E.、Faus-Kessler T.、Blader P.、Lawson N.D.等人(2010年)。 Notch活性水平控制成年神经干细胞的静止和补充之间的平衡。 《神经科学杂志》。 30, 7961–7974 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Chapouton P.、Webb K.J.、Stigloher C.、Alunni A.、Adolf B.、Hesl B.、Topp S.、Kremmer E.、Bally-Cuif L.(2011)。 成年斑马鱼大脑神经祖细胞和神经发生域中分裂基因毛/增强子的表达。 J.公司。 神经醇。 519, 1748–1769 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Coller H.A.、Sang L.和Roberts J.M.(2006年)。 细胞静止的新描述。 《公共科学图书馆·生物》。 4,e83 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Conboy I.M.,Rando T.A.(2002年)。 Notch信号的调节控制出生后肌发生中卫星细胞的激活和细胞命运的决定。 开发单元 3, 397–409 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Costa M.R.、Ortega F.、Brill M.S.、Beckervordersandforth R.、Petrone C.、Schroeder T.、Götz M.、Berninger B.(2011)。 成人神经干细胞体外分裂和谱系进展的连续实时成像。 开发 138, 1057–1068 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Dias T.B.、Yang Y.J.、Ogai K.、Becker T.和Becker C.G.(2012年)。 Notch信号控制成年斑马鱼受损脊髓中运动神经元的生成。 《神经科学杂志》。 32, 3245–3252 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Doetsch F.、García-Verdugo J.M.、Alvarez-Buylla a.(1999)。 成年哺乳动物大脑中生发层的再生。 程序。 国家。 阿卡德。 科学。 美国 96, 11619–11624 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
董政、杨N.、Yeo S.Y.、Chitnis A.、Guo S.(2012)。 谱系内定向Notch信号调节不对称分裂的放射状胶质细胞的自我更新和分化。 神经元 74, 65–78 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Draper B.W.、McCallum C.M.、Stout J.L.、Slade A.J.、Moens C.B.(2004)。 一种高通量检测斑马鱼中N-乙基-N-亚硝基脲(ENU)诱导的点突变的方法。 方法细胞生物学。 77, 91–112 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Ehm O.、Göritz C.、Covic M.、Schäffner I.、Schwarz T.J.、Karaca E.、Kempkes B.、Kremmer E.、Pfrieger F.W.、Espinosa L.等人(2010年)。 RBPJkappa依赖性信号对成人海马神经干细胞的长期维持至关重要。 《神经科学杂志》。 30, 13794–13807 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Encinas J.M.、Michurina T.V.、Peunova N.、Park J.H.、Tordo J.、Peterson D.A.、Fishell G.、Koulakov A.、Enikolopov G.(2011年)。 成年海马中分裂偶联星形细胞分化和与年龄相关的神经干细胞耗竭。 细胞干细胞 8, 566–579 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Fauq A.H.、Simpson K.、Maharvi G.M.、Golde T.、Das P.(2007年)。 γ-分泌酶抑制剂LY411575及其非对映异构体的多图化学合成。 生物有机医药化学。 莱特。 17, 6392–6395 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Folgueira M.、Bayley P.、Navratilova P.、Becker T.S.、Wilson S.W.、Clarke J.D.(2012)。 外翻硬骨端脑发育的形态发生。 神经发育。 7, 32 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Ganz J.、Kaslin J.、Hochmann S.、Freudenreich D.和Brand M.(2010年)。 斑马鱼端脑中成年神经祖细胞的异质性和Fgf依赖性。 格利亚 58, 1345–1363 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Imayoshi I.、Sakamoto M.、Yamaguchi M.、Mori K.、Kageyama R.(2010年)。 Notch信号在发育中和成人大脑中维持神经干细胞的基本作用。 《神经科学杂志》。 30, 3489–3498 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Kaslin J.、Ganz J.、Brand M.(2008年)。 非哺乳动物脊椎动物大脑的增殖、神经发生和再生。 菲洛斯。 事务处理。 R.Soc.B公司 . 363, 101–122 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Kazanis I.、Lathia J.D.、Vadakkan T.J.、Raborn E.、Wan R.、Mughal M.R.、Eckley D.M.、Sasaki T.、Patton B.、Mattson M.P.等人(2010年)。 哺乳动物大脑室管膜下区干细胞和前体细胞群的静止和激活与不同的细胞和细胞外基质信号有关。 《神经科学杂志》。 30, 9771–9781 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Kitamoto T.、Hanaoka K.(2010年)。 小鼠Notch3缺失突变通过重复肌肉再生导致肌肉增生。 干细胞 28, 2205–2216 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
克孜尔·C、卡斯林·J、克罗恩·V、布兰德·M(2012)。 斑马鱼成体神经发生和大脑再生。 神经生物学发展。 72, 429–461 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Kriegstein A.、Alvarez-Buylla A.(2009年)。 胚胎和成人神经干细胞的胶质性质。 每年。 神经科学版。 32, 149–184 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Lindsey B.W.,Tropepe V.(2006年)。 了解成人神经发生生物学原理的比较框架。 掠夺。 神经生物学。 80, 281–307 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Louvi A.,Artavanis-Tsakonas S.(2006年)。 脊椎动物神经发育中的Notch信号传导。 神经科学自然评论。 93–102年7月 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Lugert S.、Basak O.、Knuckles P.、Haussler U.、Fabel K.、Götz M.、Haas C.A.、Kempermann G.、Taylor V.、Giachino C.(2010)。 具有不同形态的静止和活跃海马神经干细胞对生理和病理刺激及衰老有选择性反应。 细胞干细胞 6, 445–456 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
马M.,江Y.J.(2007)。 Jagged2a-notch信号调节斑马鱼前肾管中的细胞命运选择。 公共科学图书馆-遗传学。 3,e18 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
März M.、Chapouton P.、Diotel N.、Vaillant C.、Hesl B.、Takamiya M.、Lam C.S.、Kah O.、Bally-Cuif L.、Strähle U.(2010年)。 斑马鱼成年端脑心室区祖细胞亚型的异质性。 格利亚 58, 870–888 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Milan D.J.、Giokas A.C.、Serluca F.C.、Peterson R.T.、MacRae C.A.(2006年)。 Notch1b和神经调节蛋白是心脏中央传导组织规范所必需的。 开发 133, 1125–1132 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Mira H.、Andreu Z.、Suh H.、Lie D.C.、Jesserger S.、Consiglio A.、San Emeterio J.、Hortigüela R.、MarquéS-Torrejón M.A.、Nakashima K.等人(2010年)。 通过BMPR-IA的信号调节成人海马神经干细胞的静止和长期活动。 细胞干细胞 7, 78–89 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Morshead C.M.、Craig C.G.、van der Kooy D.(1998)。 体内克隆分析揭示了成年哺乳动物前脑内源性神经干细胞增殖的特性。 开发 125, 2251–2261 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Mueller T.、Dong Z.、Berberoglu M.A.、Guo S.(2011)。 斑马鱼的背侧大脑皮层,斑马鱼(鲤科,真骨鱼类)。 大脑研究。 1381, 95–105 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Ninov N.、Borius M.和Stainier D.Y.(2012年)。 不同水平的Notch信号调节胰腺内分泌祖细胞的静止、更新和分化。 开发 139, 1557–1567 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Ong C.T.、Cheng H.T.、Chang L.W.、Ohtsuka T.、Kageyama R.、Stormo G.D.、Kopan R.(2006)。 脊椎动物缺口蛋白的靶选择性。 离散域和CSL绑定站点架构之间的协作决定了激活概率。 生物学杂志。 化学。 281, 5106–5119 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Perdigoto C.N.、Bardin A.J.(2013年)。 发送正确的信号:缺口和干细胞。 生物化学。 生物物理学。 学报 1830, 2307–2322 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Pierfelice T.、Alberi L.、Gaiano N.(2011年)。 脊椎动物神经系统的缺口:一只老狗有新把戏。 神经元 69, 840–855 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Rothenaigner I.、Krecsmarik M.、Hayes J.A.、Bahn B.、Lepier A.、Fortin G.、Götz M.、Jagasia R.、Bally-Cuif L.(2011)。 通过不同病毒载体的克隆分析,确定成年斑马鱼端脑中真正的神经干细胞,并表征其分裂特性和命运。 开发 1381459-1469年 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Ryu S.、Holzschuh J.、Erhardt S.、Ettl A.K.、Driever W.(2005年)。 斑马鱼视网膜中微小染色体维持蛋白5的耗竭导致细胞周期缺陷和凋亡。 程序。 国家。 阿卡德。 科学。 美国 102, 18467–18472 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Salas C.、Broglio C.、Durán E.、Gómez A.、OcañA F.M.、Jiménez-Moya F.、Rodríguez F.(2006)。 硬骨鱼类学习和记忆的神经心理学。 斑马鱼 3, 157–171 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Sang L.,Coller H.A.(2009年)。 害怕承诺:Hes1保护静止的成纤维细胞免受不可逆转的细胞命运。 细胞周期 8, 2161–2167 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Sang L.、Coller H.A.和Roberts J.M.(2008年)。 通过转录抑制因子HES1控制细胞静止的可逆性。 科学类 321, 1095–1100 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Seri B.、García-Verdugo J.M.、McEwen B.S.、Alvarez-Buylla a.(2001年)。 星形胶质细胞在成年哺乳动物海马体中产生新的神经元。 《神经科学杂志》。 21, 7153–7160 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Suh H.、Consiglio A.、Ray J.、Sawai T.、D'Amour K.A.、Gage F.H.(2007年)。 体内命运分析揭示了成年海马中Sox2+神经干细胞的多潜能和自我更新能力。 细胞干细胞 1, 515–528 [ 内政部 ][ PMC免费文章 ][ 公共医学 ][ 谷歌学者 ]
Takke C.、Dornseifer P.、v Weizsäcker E.、Campos-Ortega J.A.(1999年)。 her4是果蝇神经原基因E(spl)的斑马鱼同源物,是NOTCH信号的靶点。 开发 126, 1811–1821 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
Yoon K.,Gaiano N.(2005)。 哺乳动物中枢神经系统中的Notch信号:来自小鼠突变体的见解。 自然神经科学。 8, 709–715 [ 内政部 ][ 公共医学 ][ 谷歌学者 ]
关联数据 本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。
来自Development(英国剑桥)的文章由 生物学家公司