
摘要
在大多数文化中,饮酒是一种习惯,酒精滥用在全世界都很常见。例如,50%以上的美国人饮酒,估计有23.1%的美国人每月至少饮酒一次。尽管我们对酒精性肝病(ALD)是如何发病和进展的认识有了重大进展,但对人类酒精性肝疾病(ALD。现在很清楚,急性酗酒不仅会对肝脏产生急性毒性,还可能导致ALD的慢性化。急性酒精导致损伤的潜在机制包括脂肪变性、免疫和炎症失调以及肠道通透性改变。最近对急性酒精暴露建模的兴趣为急性损伤的潜在机制提供了新的见解,这也可能与慢性ALD有关。该小组最近关于PAI-1和纤维蛋白代谢在介导急性酒精诱导肝损伤中的作用的研究,是一个可能的新靶点的例子,这些靶点可能对急性或慢性酒精滥用有用。
介绍
在大多数文化中,饮酒是一种习惯,酒精滥用在全世界都很常见。例如,50%以上的美国人饮酒,估计有23.1%的美国人每月至少饮酒一次(药物滥用和精神健康服务管理局,2011). 饮酒的有害影响,尤其是大量饮酒和/或长期饮酒,已被公认。饮酒最常见的症状与慢性酒精中毒有关,它是60多种主要疾病的病因/风险因素。饮酒的这些和其他影响使酒精成为全球第三大疾病和残疾风险因素(世界卫生组织,2011). 1998年,美国酗酒的医疗后果预计将超过260亿美元,使其成为卫生保健系统的重大财政负担(美国卫生与公众服务部,2000). 这些估计仅考虑了直接医疗费用;据估计,1998年美国因酒精相关疾病导致的生产力损失约为900亿美元。因此,饮酒的医疗后果除了对个人的影响外,还具有重大的社会后果。
肝脏是酒精代谢的主要场所,也是酒精损伤的主要靶器官。肝脏对酒精诱导毒性的易感性是由于门静脉血中发现的高浓度酒精(与全身相比),以及乙醇代谢的代谢后果。酒精性肝病(ALD)是一系列疾病状态,包括脂肪变性(脂肪肝)、脂肪性肝炎,严重时包括纤维化和/或肝硬化。以肝细胞中脂肪堆积为特征的脂肪变性,在每天饮酒超过16克的人中有90%会发生(Crabb,1999),但决定停止饮酒(Bergheim等人。,2005). 只有少数即使是重度饮酒者,脂肪变性也会发展为脂肪性肝炎,其特征是脂肪肝持续存在并伴有炎症。在ALD的后期,胶原沉积和再生结节可分别导致纤维化和肝硬化的发展。戒酒对处于ALD所有阶段的患者都是有益的,并且对于预防早期疾病患者的肝损伤进展是必要的(Bergheim等人。,2005). 不幸的是,慢性酒精滥用者的高累犯率大大降低了通过节欲缓解疾病的机会。因此,更好地理解酒精损伤肝脏的机制可能会产生新的药理学策略,以减缓、阻止或逆转疾病的进展,甚至可能是在顽固的酗酒者中。
尽管对酒精暴露的研究已有数十年,但大多数工作都集中在慢性暴露,我们对急性酒精暴露的机制和影响缺乏了解。急性酒精接触,俗称“狂饮”,是由国家药物使用与健康调查(NSDUH)定义为在3个时间段内单次饮酒5杯或更多h.然而,急性酒精接触也可能包括持续数天的大量饮酒或间歇性、反复的大量饮酒。例如,在临床环境中,“狂饮”的一幕通常描述了一段持续2天以上的饮酒和醉酒时期(韦施勒和奥斯汀,1998). 2010年的NSDUH显示,超过5800万美国人在30天内至少参与过一次狂饮(药物滥用和心理健康服务管理局,2011). 由于18-25岁人群中狂饮的流行率越来越高,人们越来越关注急性酒精暴露的影响。虽然慢性酒精滥用是最常见的由饮酒引起的疾病来源,但急性酒精滥用也是有害的。许多与酒精相关的急性死亡是由于严重的中枢神经系统抑制,这会减缓运动协调和呼吸频率。急性酒精暴露也会损害感染和创伤的恢复,并延迟伤口愈合(Radek等人。,2005,2012). 因此,急性饮酒是入院期间发病率和死亡率的主要潜在原因(Jones等人。,1991).
急性酒精性肝损伤
目前对酗酒对肝损伤的影响的认识还不如我们对慢性酒精暴露影响的认识那么全面。然而,急性和慢性酒精暴露之间有一些相似之处。下文讨论了急性酒精导致肝损伤的潜在机制。
脂肪变性的机制
如上所述,第一个也是最常见的由饮酒引起的肝脏变化是脂肪变性或脂肪肝。脂肪积聚可以是大泡状的(每个肝细胞有一个大的脂肪滴和细胞核的侧向移位)或微泡状的(每个肝细胞有许多小的脂肪滴;Ishak等人。,1991). 酒精暴露导致脂肪变性的一个机制是直接通过酒精代谢。饮酒期间,门静脉/肝脏循环中的酒精浓度很容易达到mM范围。在将乙醇代谢为醋酸盐的过程中,每氧化一当量乙醇,就会产生两当量的还原NADH。这种新陈代谢会显著增加NADH:NAD的比率+然后抑制肝脏中脂肪酸的β-氧化。此外乙醇代谢也增加了脂肪酸的酯化速率(Ontko,1973). 净效应是有利于脂肪在肝细胞中积聚。
虽然酒精代谢可能足以导致脂肪堆积,但这并不是脂肪变性形成的唯一机制。阻断脂多糖(LPS)信号传导(Yin等人。,2001)和产生前氧化剂的酶(Kono等人。,2000; McKim等人。,2003)在啮齿类动物中,通过基因改变也可以防止脂肪变性,而不会改变乙醇代谢。脂肪积累也可以由改变脂质代谢的介质如促炎细胞因子刺激(Grunfield等人。,1990). 这些细胞因子下游的信号通过两种方式促进肝脏脂肪堆积——增加脂肪酸的沉积,减少脂质向循环中的分解和分泌。例如,TNFα增加了外周脂肪细胞释放的游离脂肪酸(Hardardottir等人。,1992),增加从头开始脂质合成(Feingold和Grunfeld,1987),并抑制脂肪酸的β-氧化(Nachiappan等人。,1994). 最终,这些变化通过增加脂肪酸供应刺激肝脏中的脂肪积累,同时损害肝脏的脂肪酸代谢和分泌能力。
即使摄入适量酒精,大多数人的肝脏脂肪变性也会加剧。在狂饮的啮齿类动物模型中也可以看到蒸汽变化(Kaiser等人。,2009; Donohue等人。,2011). 脂肪增多症被认为是一种无症状的疾病状态,一旦禁食,这种状态很容易逆转(Bergheim等人。,2005). 事实上,在机体层面上,肝脏脂肪变性可以被视为一种保护措施,因为它将脂质从血液中分离出来,并将其储存起来以备日后使用(van Ginneken,2008). 然而,尽管脂肪变性是一种惰性病理学就其本身而言,它使肝脏对第二次侮辱造成的损伤敏感(Day和James,1998). 根据这个“双重打击假说”,单靠饮酒并不会导致ALD从脂肪变性发展为脂肪性肝炎、肝硬化或纤维化;相反,第二个危险因素或侮辱是疾病后期发展所必需的。第二种侮辱可能包括炎症反应、活性氧或缺氧等。例如,众所周知,急性乙醇暴露会增强外源性物质(例如细菌细胞壁产物(例如脂多糖))诱导的肝脏病理学。此外,急性饮酒本身也可能造成第二次打击;例如,一次狂饮会引发慢性酒精中毒者发生急性酒精性肝炎(Rivara等人。,1993; Barrio等人。,2004).
免疫和炎症失调
“启动”和“致敏”的概念在分子水平上反映了双重打击假说,而这两个概念现在被认为是酒精诱导肝损伤的基础(Tsukamoto等人。,2001). 这里,“启动”是指乙醇预暴露导致肝脏炎性细胞(例如库普弗细胞)在第二次刺激(如脂多糖)的反应下更强烈地释放促炎细胞因子的能力。酒精性肝炎患者在基础和刺激后都有大量的循环TNFα(McClain和Cohen,1989). 此外,乙醇预暴露可使库普弗细胞受到LPS刺激,导致TNF-α释放增强(Enomoto等人。,1998). 乙醇也能使细胞增敏,导致炎症细胞因子信号下游的细胞群反应更强烈。脂肪肝患者对脂多糖诱导的肝损伤更敏感(Yang等人。,1997). 在急性酒精暴露的啮齿动物模型中,对脂多糖的敏感性被重新描述(Bergheim等人。,2006; Beier等人。,2009).
尽管一些研究表明,酒精暴露会使巨噬细胞产生更多炎症组织损伤,但其他研究表明,急性酒精中毒(即系统中存在酒精时)实际上会损害免疫反应。例如,酗酒会增加感染的风险(例如Gentilello等人。,1993; Gamble等人。,2006; Griffin等人。,2009). 此外,急性乙醇摄入会恶化脓毒症的预后(Lin等人。,2009). 酒精能够削弱免疫反应的一个主要机制是通过改变Toll样受体(TLR)信号,TLR信号作为巨噬细胞和先天免疫系统其他细胞的模式识别受体(Beutler,2009). TLR参与巨噬细胞的直接细胞毒性和效应性反应,因此是适当免疫反应的关键早期启动因子。重要的是,乙醇中毒已被证明可以减弱许多TLR配体对巨噬细胞的刺激,包括酵母多糖a(TLR2)和脂多糖(TLR4;Pruett等人。,2004; 戈尔和科瓦茨,2005).
尽管急性乙醇对巨噬细胞反应性的这些影响似乎是矛盾的(即损害和启动巨噬细胞),但与乙醇暴露相关的炎症刺激的时间似乎很重要。例如,急性乙醇(4-6 g/kg)在乙醇暴露3小时后(中毒期间)注射LPS,可以减轻LPS引起的炎症性肝损伤,但如果在乙醇暴露24小时后注射LPS则会加重损伤(Enomoto等人。,1998). 急性乙醇导致先天免疫系统耐受和启动的机制尚不完全清楚。然而,很明显,急性酒精暴露会产生一种危险情况,即在急性中毒期间感染的可能性增加,但对该感染的炎症反应会加剧肝脏损伤。
肠道通透性和内毒素
即使没有感染,酒精也会改变对炎症刺激的反应。自20世纪70年代以来,循环细菌内毒素(即LPS)增加与肝损伤之间的关系就已经确立(诺兰,2010). ALD患者的循环LPS水平增加(Bode等人。,1987; 福井等人。,1991)以及在两种急性病的实验模型中(Lambert等人。,2003)和慢性(Nanji等人。,1994; Keshavarzian等人。,2001)饮酒。饮酒导致循环LPS增加,至少部分是由于肠道通透性增加,从而使LPS转移到门静脉血中。事实上,消除肠道细菌可以通过降低小鼠循环内毒素水平来减轻酒精诱导的肝损伤(Nanji等人。,1994). 两种慢性病的实验模型(Mathurin等人。,2000; Tamai等人。,2002)和急性(Tamai等人。,2000; Lambert等人。,2003)酒精暴露表明,酒精增加了LPS的渗透性。急性饮酒增加了健康志愿者和ALD患者的LPS循环水平(Bode等人。,1987; 福井等人。,1991).
在正常情况下,构成肠屏障的上皮细胞之间的间隙被紧密连接所封闭。这些紧密连接防止有毒化合物(如LPS)转移到循环中(Purohit等人。,2008). 乙醇引起的肠道通透性变化可能部分是由于小肠顶端连接蛋白的破坏。体外使用Caco-2肠细胞进行的研究表明,酒精可以破坏ZO-1紧密连接,最终导致细胞旁间隙出现间隙(Ma等人。,1999). 此外,慢性酒精暴露降低了小鼠回肠中的ZO-1和occludin(Zhong等人。,2010). 同样的研究也表明,紧密连接蛋白的减少与肠道中的氧化应激有关。酒精暴露后胃肠道氧化应激的潜在来源包括乙醛和一氧化氮(Purohit等人。,2008). 急性酒精暴露导致的肠道通透性可能与类似机制有关。
总结
总之,急性酒精暴露可导致“完美风暴”,有利于炎症性肝损伤。急性酒精暴露会增加感染和胃肠道通透性的风险。其中一种或两种机制都会增加TLR配体向肝脏巨噬细胞的递送。在酒精暴露后的稍后时间,巨噬细胞对TLR配体的炎症反应被激发,产生更多的细胞毒性细胞因子(例如TNFα)。此外,实质细胞中的脂肪变性使其对巨噬细胞释放的细胞因子的细胞毒性杀伤敏感。
急性乙醇暴露模型
大多数关于酒精性肝损伤的研究都调查了长期饮酒的影响。然而,强调急性饮酒有害影响的临床证据,最近激发了人们对狂饮、间歇大量饮酒和其他急性酒精暴露的实验模型的兴趣。这些模型包括在体外和体内范式,后者涵盖许多物种,包括啮齿动物、狗、灵长类动物和微型动物(Dolganiuc和Szabo,2009). 大多数急性酒精研究都是在啮齿动物模型中进行的。这种模型实现了与人类早期肝损伤相似的病理状态(例如脂肪变性、炎症)。自从开发出各种敲除鼠和其他转基因鼠种以来,研究界对啮齿动物的使用有所增加,这些转基因鼠种可以进行更方便和具体的机理研究。
啮齿动物接触酒精模型的主要障碍之一是它们对乙醇的厌恶。啮齿类动物不会自愿饮用浓度足以重现人类肝脏疾病的酒精。因此,通过胃内灌胃强制给药是模拟急性酒精滥用的最常见方法(Siegmund等人。,2005). 急性酒精暴露的啮齿动物模型通常分为三类:单剂量模型、多剂量模型和“二次打击”模型。乙醇以高达6 g/kg的剂量给药,这考虑到啮齿动物与人类乙醇动力学的差异(格什曼和斯蒂珀,1991; 卡森和普鲁特,1996). 啮齿类动物体内的酒精峰值浓度与人类在急性饮酒狂欢期间所能达到的浓度类似。
慢性模型的成本很高,而且在慢性肝损伤模型中通常很难区分影响和近因。因此,除了研究急性肝毒性外,急性模型还可用作筛选工具和/或机制分析,以补充慢性研究。这一方法得到了以下观察结果的支持:急性饮酒的动物与慢性乙醇研究中的动物相似,会发生脂肪变性和炎症。在急性和慢性酒精暴露模型中,保护剂的机械重叠进一步强化了“模型模型”的概念。例如,Enomoto等人(2000)证明在慢性接触模型中已知可保护肝脏免受损伤的化合物[例如GdCl三(阿达奇等人。,1994)、抗生素(Adachi等人。,1995)尼莫地平(Iimuro等人。,1996)]在急性乙醇暴露的大鼠模型中也能防止脂肪变性。由于急性酒精暴露模型被用作慢性酒精暴露的模型,因此对于筛选可能提供治疗益处的化合物以及研究治疗化合物下游的新机制特别有价值。到目前为止,急性酒精暴露模型在用于筛选慢性研究的治疗分子的疗效时几乎没有出现假阳性。
PAI-1和纤维蛋白——急性酒精性肝损伤的新介质
纤溶酶原激活物抑制剂-1
纤溶酶原激活物抑制物-1(PAI-1)是一种急性期蛋白,至少部分通过调节纤维蛋白溶解在损伤和炎症中发挥重要作用。PAI-1通过抑制尿激酶型和组织型纤溶酶原激活剂(uPA和tPA;图1; 克鲁索夫,1988). PAI-1在酒精诱导肝损伤中的作用首次见于PA:PAI-1比率升高的肝硬化患者(Violi等人。,1992; Hu等人。,2001)导致高纤溶。相反,在ALD的发展过程中,循环PAI-1水平升高,PA:PAI-1较低,有利于低纤维蛋白溶解(Marques-Vidal等人。,1995; Dimmitt等人。,1998; Mukamal等人。,2001). 在酒精诱导肝脂肪变性的实验模型中,PAI-1水平与脂质积聚程度相关(Alessi等人。,2003),并可用作ALD后期严重程度的指标(Tran-Tang等人。,1989).
图1。
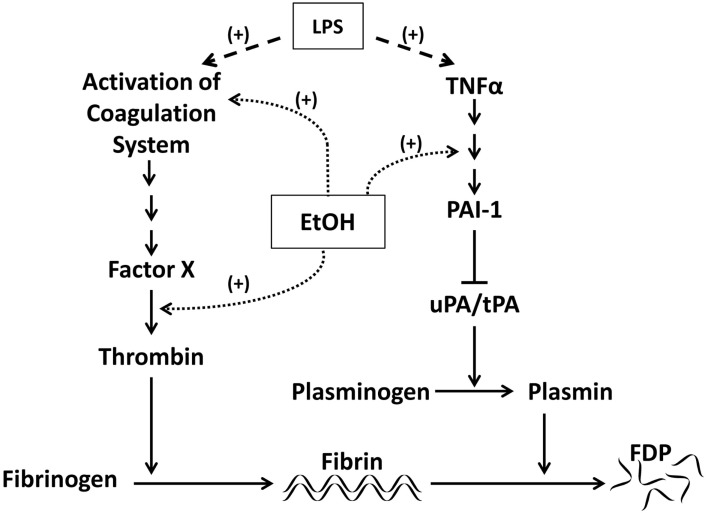
脂多糖和乙醇对PAI-1和纤溶的影响LPS通过凝血酶激活凝血级联反应导致交联纤维蛋白的积累。交联纤维蛋白被纤溶酶降解。PAI-1通过抑制纤溶酶原激活剂uPA和tPA从而抑制纤溶酶的激活来阻断纤维蛋白溶解。乙醇和LPS都可以调节凝血级联下游的信号传导,增加纤维蛋白沉积。乙醇还可以通过改变TNFα下游的信号传导来减缓纤维蛋白降解。LPS通过刺激TNFα的释放,对纤溶有类似的作用。
虽然PAI-1在某些疾病(如动脉粥样硬化)中的重要性已经确定,但PAI-1对ALD的作用仍不清楚。使用敲除小鼠菌株,特别是PAI-1的研究−/−小鼠已经开始阐明PAI-1在脂质积累中的作用。Ma等人(2004)表明PAI-1−/−在高脂肪/高碳水化合物饮食诱导的肥胖模型中,小鼠可以避免肝脏脂质积聚。此外,我们的研究小组已经表明,急性和慢性乙醇诱导的脂肪变性都与肝脏PAI-1的表达相关,阻断PAI-1,无论是在药理学上还是遗传学上,都可以防止脂肪变性(Bergheim等人。,2006). 在本研究中,TNFR1−/−同时,PAI-1钝化也能保护小鼠免受脂肪变性的影响(Bergheim等人。,2006)这表明酒精暴露后PAI-1的诱导机制是TNFα下游,可能通过MAPK信号传导(Fearns和Loskutoff,1997). PAI-1似乎在介导器官炎症方面也很关键。例如,在肾小球肾炎模型中,缺乏PAI-1的小鼠可以防止白细胞浸润和组织损伤(Kitching等人。,2003). 该实验室的工作表明,PAI-1−/−小鼠几乎完全免受慢性酒精引起的炎症(Arteel,2008). PAI-1如何介导炎症,尤其是在饮酒后,尚不清楚。一种潜在的机制是PAI-1介导炎症反应中重要细胞因子的蛋白水解激活/失活的变化。这些影响可能包括阻止细胞因子的激活,包括转化生长因子-β(Espevik等人。,1987; 佐藤等人。,1990)或稳定促炎细胞因子,如趋化剂IL-8(Marshall等人。,2003).
纤维蛋白
PAI-1炎症效应的另一潜在机制涉及纤溶受损和随后的纤维蛋白积聚(Holdsworth等人。,1979; Loike等人。,1995). 纤维蛋白(原)是细胞外基质(ECM)的一种结构成分。凝血级联激活后,潜在形式纤维蛋白原被凝血酶分解为活性形式纤维蛋白(图1). 纤维蛋白可以交联形成血栓或被纤溶酶降解为纤维蛋白原降解产物。除了在止血方面的作用外,纤维蛋白还可以通过与细胞表面受体的相互作用发挥信号分子的作用。纤维蛋白在炎症中的作用因其能够发挥多种促炎症作用而得到强调。例如,Qi等人(1997)显示纤维蛋白诱导血管内皮细胞中IL-8的表达。纤维蛋白基质可以调节炎症细胞的迁移在体外包括IL-8刺激的中性粒细胞(Loike等人。,1995). 此外,除颤可减少肾脏中巨噬细胞的迁移(Holdsworth等人。,1979).
纤维蛋白在实验性ALD中也有直接作用。我们的研究小组已经表明,慢性乙醇摄入会增加肝窦中的纤维蛋白沉积(图2)在酒精诱导的急性肝损伤模型中,纤维蛋白积聚与炎症相关。此外,使用凝血酶抑制剂水蛭素直接阻断纤维蛋白沉积,或使用MEK抑制剂U0126间接阻断纤维蛋白沉积,可防止酒精预暴露引起的肝损伤增强(Beier等人。,2009). 阻止纤维蛋白积累也可以防止炎症(Beier等人。,2009). 这些数据表明纤维蛋白在酒精暴露后PAI-1介导的炎症和损伤增强中起重要作用。
图2。
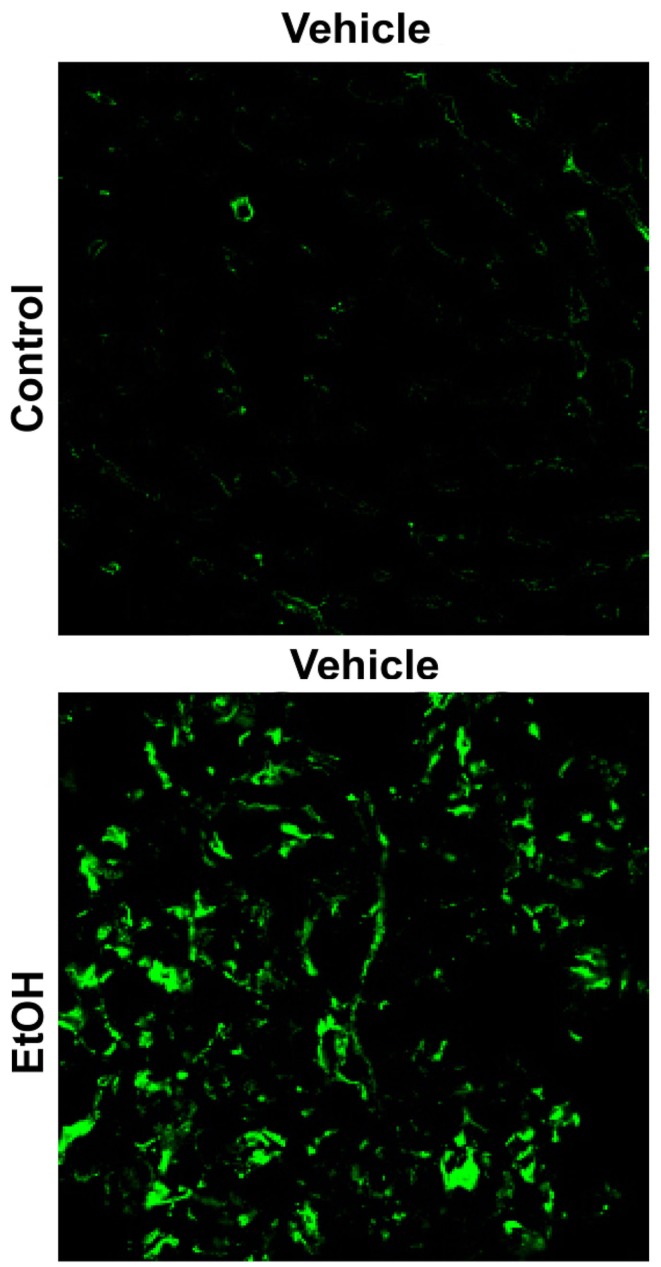
慢性乙醇增加肝纤维蛋白(原)沉积显示了描述纤维蛋白(原)ECM免疫荧光检测的代表性显微照片(40×)。肝组织取自连续4周喂食控制饮食或肠内乙醇饮食的小鼠。
整合素–细胞与细胞间通信的介质
虽然该组显示了酒精诱导的纤维蛋白沉积与肝脏炎症和损伤之间的密切关系,但纤维蛋白介导其炎症效应的机制尚不清楚。纤维蛋白积聚介导炎症效应的一种机制是通过与细胞表面受体(如整合素)的相互作用。整合素是多种细胞类型上表达的异二聚体受体,包括内皮细胞、炎症细胞(包括巨噬细胞和中性粒细胞)和血小板。在与细胞外成分(包括周围基质和细胞)相互作用后,整合素的β亚单位发生快速构象变化。整合素结构的这种变化允许通过细胞内信号通路的激活跨膜传递信息。包括整合素连接激酶(ILK)在内的适配器蛋白是整合素激活下游信号转导的重要介质,并在确定细胞内激活哪些信号级联中发挥作用。整合素是一个具有至少24种不同异二聚体的多样超家族,在包括增殖、血管生成和炎症在内的多种细胞过程中发挥重要作用。
纤维蛋白(原)ECM能够与细胞表面粘附受体的整合素家族相互作用,例如整合素αv(v)β三(Cheresh等人。,1989). 整合素α的作用v(v)β三肿瘤中的血管生成和转移已被充分记录,αv(v)β三整合素已被证明有助于炎症(Zhou等人。,2009). 整合素αv(v)β三内皮细胞强烈表达(Carloni等人。,1996)包括排列在肝窦(HSEC)的区域,这是一个在酒精暴露后与纤维蛋白积聚共存的区域(图2). 整合素αv(v)β三通过蛋白质上的RGD序列结合纤维蛋白(原)(Cheresh等人。,1989),在药理学上可与小肽拮抗剂结合。该小组的研究表明v(v)β三在急性酒精性肝损伤的“2击”模型中,使用小肽拮抗剂CycloRGDfV可以防止急性酒精引起的LPS诱导的肝损伤加重(图三)对纤维蛋白积累没有任何影响。此外,在该模型中,CycloRGDfV对肝脏损伤的保护与LPS诱导的中性粒细胞浸润的减弱同时发生(图三). 这些数据表明,急性酒精暴露后LPS引起的炎症和肝损伤可能是由于正弦纤维蛋白和整合素α之间的相互作用所致v(v)β三.
图3。
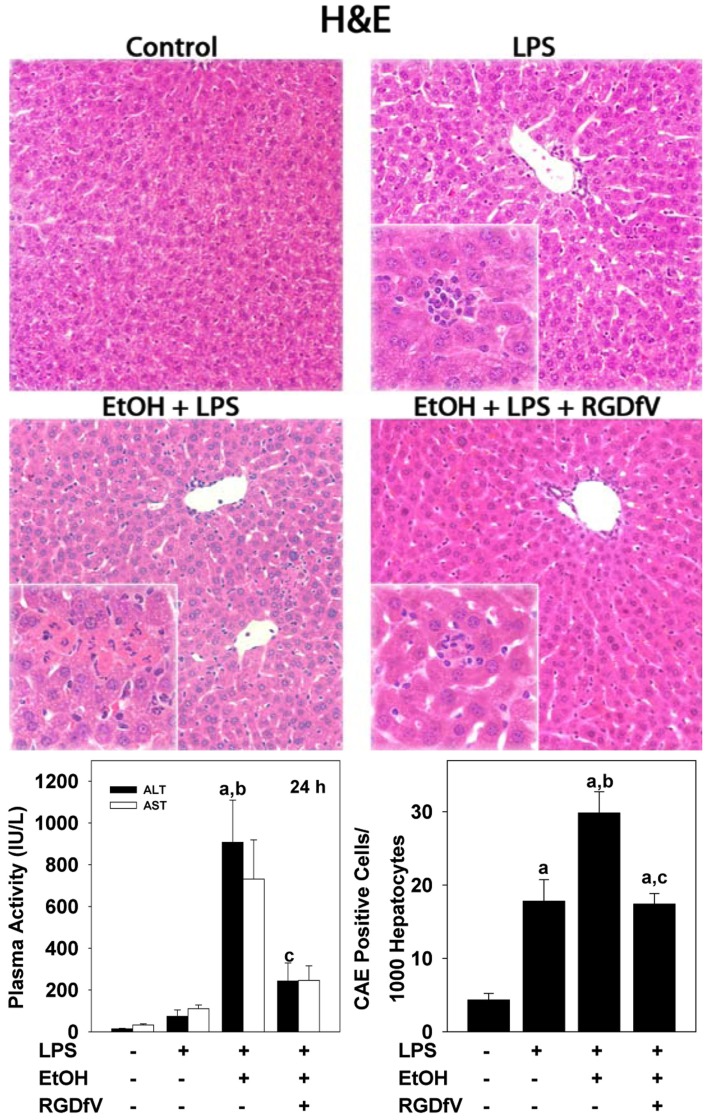
整合素α的抑制作用v(v)β三防止酒精预暴露导致LPS诱导的肝损伤加重苏木精和伊红染色的代表性显微照片(100×;插图400×)显示在上面板中。循环ALT水平和中性粒细胞迁移(由CAE染色确定)分别位于左下方和右下方。将动物暴露于酒精(6 g/kg i.g)中3天,然后给予LPS(10 mg/kg;i.p)最后一次给药后24h,给药24h后处死。一些动物在LPS前1小时服用CycloRGDfV(3 mg/kg i.p.)。
总结与结论
尽管我们对该病如何发生和发展的认识有了重大进展,但对人类ALD的安全有效治疗仍然难以捉摸。很明显,酗酒不仅会对肝脏产生急性毒性,还可能导致ALD的慢性化。急性酒精导致损伤的潜在机制包括脂肪变性、免疫和炎症失调以及肠道通透性改变。最近对急性酒精暴露建模的兴趣为急性损伤的潜在机制提供了新的见解,这也可能与慢性ALD有关。该小组最近关于PAI-1和纤维蛋白代谢在介导急性酒精诱导肝损伤中的作用的研究,是一个可能的新靶点的例子,这些靶点可能对急性或慢性酒精滥用有用。
利益冲突声明
作者声明,该研究是在没有任何可能被解释为潜在利益冲突的商业或金融关系的情况下进行的。
致谢
这项工作部分得到了国家酒精滥用和酗酒研究所(AA003624)的资助。Veronica L.Massey得到了美国国家环境健康科学研究所(ES011564)博士前(T32)奖学金的支持。
工具书类
-
Adachi Y.、Bradford B.U.、Gao W.、Bojes H.K.、Thurman R.G.(1994)。Kupffer细胞失活可预防早期酒精诱导的肝损伤。肝病学20、453–46010.1002/hep.184020227[内政部] [公共医学] [谷歌学者]
-
Adachi Y.、Moore L.E.、Bradford B.U.、Gao W.、Thurman R.G.(1995)。抗生素可预防大鼠长期接触乙醇后的肝损伤。胃肠病学108, 218–22410.1016/0016-5085(95)90027-6[内政部] [公共医学] [谷歌学者]
-
Alessi M.C.、Bastelica D.、Mavri A.、Morange P.、Berthet B.、Grino M.、Juhan-Vague I.(2003)。血浆PAI-1水平与肝脂肪变性的关系比与脂肪组织积聚的关系更密切。动脉硬化。血栓。瓦斯克。生物。231262–126810.1161/01.ATV.0000077401.36885.BB[内政部] [公共医学] [谷歌学者]
-
Arteel G.E.(2008)。纤溶酶原激活物抑制剂-1在酒精诱导肝损伤中的新作用。《肠胃病学杂志》。肝素。23,S54–S5910.1111/j.1440-1746.2007.05285.x[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Barrio E.、Tome S.、Rodriguez I.、Gude F.、Sanchex-Leira J.、Perez-Becerra E.、Gonzalez-Quintela A.(2004)。伴有或不伴有酒精戒断综合征的酗酒者的肝病。酒精。临床。实验结果。28、131–13610.1097/01.ALC.000106301.39746.EB[内政部] [公共医学] [谷歌学者]
-
Beier J.I.、Luyendyk J.P.、Guo L.、von Montfort C.、Staunton D.E.、Arteel G.E.(2009)。纤维蛋白的积累在小鼠对乙醇引起的脂多糖诱导的肝损伤的致敏性中起着关键作用。肝病学1545年9月49日至155310.1002年9月22847日[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Bergheim I.、Guo L.、Davis M.A.、Lambert J.C.、Beier J.I.、Duveau I.、Luyendyk J.P.、Roth R.A.、Arteel G.E.(2006)。二甲双胍预防小鼠酒精诱导的肝损伤:纤溶酶原激活物抑制剂-1的关键作用。胃肠病学1302099–211210.1053/j.gastro.2006.03.020[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Bergheim I.、McClain C.J.、Arteel G.E.(2005)。酒精性肝病的治疗。挖掘。数字化信息系统。23275–28410.1159/000090175[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Beutler B.(2009)。微生物传感、正反馈回路和炎症疾病的发病机制。免疫学。版次。227248–26310.1111/j.1600-065X.2008.00733.x[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Bode C.H.、Kugler V.、Bode J.C.H.(1987)。酒精性和非酒精性肝硬化患者以及急性酒精过量后无慢性肝病证据的受试者的内毒素血症。《肝素杂志》。4、8–1410.1016/S0168-8278(87)80003-X[内政部] [公共医学] [谷歌学者]
-
Carloni V.、Romanelli R.G.、Pinzani M.、Laffi G.、Gentilini P.(1996年)。胶原和层粘连蛋白整合素受体在培养的人肝星状细胞中的表达和功能。胃肠病学110、1127–113610.1053/gast.1996.v110.pm8613002[内政部] [公共医学] [谷歌学者]
-
Carson E.J.、Pruett S.B.(1996年)。小鼠狂饮模型的开发和表征,用于评估乙醇的免疫效果。酒精。临床。实验结果。20、132–13810.1111/j.1530-0277.1996.tb01055.x[内政部] [公共医学] [谷歌学者]
-
Cheresh D.A.、Berliner S.A.、Vicente V.、Ruggeri Z.M.(1989年)。血小板和内皮细胞上相关整合素对纤维蛋白原上不同粘附位点的识别。单元格58945–95310.1016/0092-8674(89)90946-X[内政部] [公共医学] [谷歌学者]
-
Crabb D.W.(1999)。酒精性肝病的发病机制:新的损伤机制。庆应。医学杂志。48、184–18810.2302/kjm.48.184[内政部] [公共医学] [谷歌学者]
-
Day C.P.,James O.F.(1998)。脂肪性肝炎:两次“袭击”的故事胃肠病学114、842–84510.1016/S0016-5085(98)83433-1[内政部] [公共医学] [谷歌学者]
-
Dimmitt S.B.、Rakic V.、Puddey I.B.、Baker R.、Oostryck R.、Adams M.J.、Chesterman C.N.、Burke V.、Beilin L.J.(1998)。酒精对凝血和纤溶因子的影响:一项对照试验。血液凝固。纤维蛋白溶解9, 39–4510.1097/00001721-199801000-00005[内政部] [公共医学] [谷歌学者]
-
Dolganiuc A.、Szabo G.(2009年)。急性酒精暴露的体外和体内模型。世界胃肠病学杂志。邮编:151168–117710.3748[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Donohue T.M.,Jr.、Osna N.、Trambly C.、Whitaker N.、Thomes P.、Todero S.、Davis J.S.(2011)。早期生长反应-1有助于急性乙醇摄入后脂肪变性的发展。酒精。临床。实验结果。36、759–76710.1111/j.1530-0277.2011.01681.x[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Enomoto N.、Ikejima K.、Bradford B.U.、Rivera C.A.、Kono H.、Brenner D.A.、Thurman R.G.(1998)。酒精通过依赖内毒素的机制导致大鼠Kupffer细胞的耐受性和致敏性。胃肠病学115443–45110.1016/s016-5085(98)70211-2[内政部] [公共医学] [谷歌学者]
-
Enomoto N.、Ikejima K.、Yamashina S.、Enomota A.、Nishiura T.、Nishimura T.、Brenner D.A.、Schemmer P.、Bradford B.U.、Rivera C.A.、Zhong Z.、Thurman R.G.(2000)。库普弗细胞衍生前列腺素E(2)参与酒精诱导的大鼠肝脏脂肪堆积。美国生理学杂志。279,G100–G106[内政部] [公共医学] [谷歌学者]
-
Espevik T.、Figari I.S.、Shalaby M.R.、Lackides G.A.、Lewis G.D.、Shepard H.M.、Palladino M.A.,Jr.(1987)。环孢菌素A和转化生长因子β对细胞因子产生的抑制。实验医学学报166、571–57610.1084/jem.166.2.571[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Fearns C.,Loskutoff D.J.(1997年)。脂多糖诱导小鼠肝脏纤溶酶原激活物抑制剂1基因表达。内源性肿瘤坏死因子α的细胞定位和作用。美国病理学杂志。150, 579–590[PMC免费文章] [公共医学] [谷歌学者]
-
Feingold K.R.、Grunfeld C.(1987)。肿瘤坏死因子α在体内刺激大鼠肝脏脂肪生成。临床杂志。投资。80、184–19010.1172/JCI113046[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Fukui H.、Brauner B.、Bode J.C.和Bode C.(1991年)。酒精性和非酒精性肝病患者的血浆内毒素浓度:用改进的显色分析法重新评估。《肝素杂志》。12, 162–16910.1016/0168-8278(91)90933-3[内政部] [公共医学] [谷歌学者]
-
Gamble L.、Mason C.M.和Nelson S.(2006年)。酒精对免疫和肺部细菌感染的影响。Med.Mal.感染。36、72–7710.1016/j.medmal.2005.08.010[内政部] [公共医学] [谷歌学者]
-
Gentilello L.M.、Cobean R.A.、Walker A.P.、Moore E.、Wertz M.J.、Dellinger E.P.(1993)。急性乙醇中毒会增加腹部穿透伤后感染的风险。J.创伤。35, 669–67410.1097/00005373-199309000-00011[内政部] [公共医学] [谷歌学者]
-
Gershman H.、Steeper J.(1991)。急诊室醉酒患者血液中乙醇的清除率。急诊医学杂志。9307–31110.1016/0736-4679(91)90371-L[内政部] [公共医学] [谷歌学者]
-
Goral J.、Kovacs E.J.(2005年)。体内乙醇暴露通过限制p38和ERK1/2的激活,下调TLR2-、TLR4-和TLR9-介导的巨噬细胞炎症反应。免疫学杂志。174, 456–463[内政部] [公共医学] [谷歌学者]
-
Griffin R.、Poe A.M.、Cross J.M.、Rue L.W.、McGwin G.(2009年)。烧伤患者血液酒精水平与感染并发症的关系。J.烧伤护理康复。30395–39910.1097/BCR.0b013e3181a28966[内政部] [公共医学] [谷歌学者]
-
Grunfield C.、Soued M.、Adi S.、Moser A.H.、Dinarello C.A.、Feingold K.R.(1990)。两类刺激肝脏脂肪生成的细胞因子的证据:肿瘤坏死因子、白细胞介素-1和干扰素-α之间的关系。内分泌学127,46–5410.1210/内-127-1-46[内政部] [公共医学] [谷歌学者]
-
Hardardottir I.、Doerrler W.、Feingold K.R.、Grunfeld C.(1992年)。细胞因子通过前列腺素非依赖性机制刺激脂肪细胞中的脂肪分解并降低脂蛋白脂肪酶活性。生物化学。生物物理学。Res.Commun公司。186237–24310.1016/S0006-291X(05)80798-3[内政部] [公共医学] [谷歌学者]
-
Holdsworth S.R.、Thomson N.M.、Glasgow E.F.、Atkins R.C.(1979年)。去纤维蛋白对兔肾毒性肾炎巨噬细胞参与的影响:采用肾小球培养和电镜的研究。临床。实验免疫学。37、38–43[PMC免费文章] [公共医学] [谷歌学者]
-
胡克强、于亚瑟、蒂亚古拉、雷德克·A·G、雷诺兹·T·B(2001)。转诊肝单位住院肝硬化患者的高纤溶活性。美国胃肠病杂志。96、1581–158610.1111/j.1572-0241.2001.03781.x[内政部] [公共医学] [谷歌学者]
-
Iimuro Y.、Ikejima K.、Rose M.L.、Bradford B.U.、Thurman R.G.(1996)。尼莫地平是一种二氢吡啶型钙通道阻滞剂,可预防大鼠慢性胃内乙醇暴露导致的酒精性肝炎。肝病学24、391–39710.1016/S0168-8278(96)80158-9[内政部] [公共医学] [谷歌学者]
-
Ishak K.G.、Zimmerman H.J.、Ray M.B.(1991年)。酒精性肝病:病理、病因和临床方面。酒精。临床。实验结果。15、45–6610.1111/j.1530-0277.1991.tb00518.x[内政部] [公共医学] [谷歌学者]
-
Jones J.D.、Barber B.、Engrav L.、Heimback D.(1991年)。饮酒和烧伤。J.烧伤护理康复。12, 148–15210.1097/00004630-199103000-00012[内政部] [公共医学] [谷歌学者]
-
Kaiser J.P.、Beier J.I.、Zhang J.、David H.J.、von Montfort C.、Guo L.、Zheng Y.、Monia B.P.、Bhatnagar A.、Arteel G.E.(2009)。PKCepsilon在急性乙醇诱导的脂肪变性中起着因果作用。架构(architecture)。生物化学。生物物理学。482,104–11110.1016/j.abb.2008.11.004[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Keshavarzian A.、Choudhary S.、Holems E.W.、Young S.、Banan A.、Jakate S.、Fields J.Z.(2001)。通过补充燕麦预防肠道渗漏可以改善大鼠酒精诱导的肝损伤。《药理学杂志》。实验治疗师。299, 442–448[公共医学] [谷歌学者]
-
Kitching A.R.、Kong Y.Z.、Huang X.R.、Davenport P.、Edgton K.L.、Carmeliet P.、Holdsworth S.R.、Tipping P.G.(2003)。纤溶酶原激活物抑制剂-1是实验性新月体肾炎肾损伤的重要决定因素。《美国肾脏学会杂志》。14,1487–149510.1097/01.ASN.0000065550.13931.00[内政部] [公共医学] [谷歌学者]
-
Kono H.、Rusyn I.、Yin M.、Gabele E.、Yamashina S.、Dikalova A.、Kadiiska M.B.、Connor H.D.、Mason R.P.、Segal B.H.、Bradford B.U.、Holland S.M.、Thurman R.G.(2000)。NADPH氧化酶衍生自由基是酒精诱导肝病的关键氧化剂。临床杂志。投资。106、867–87210.1172/JCI9020[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Kruithof E.K.(1988)。纤溶酶原激活物抑制剂——综述。酶40, 113–121[内政部] [公共医学] [谷歌学者]
-
Lambert J.C.、Zhou Z.、Wang L.、Song Z.、McClain C.J.、Kang Y.J.(2003)。锌对小鼠急性乙醇诱导肝损伤的抑制作用与防止肠道通透性改变有关。《药理学杂志》。实验治疗师。305880–88610.1124/日元102.047852[内政部] [公共医学] [谷歌学者]
-
Lin J.N.、Tsai Y.S.、Lai C.H.、Chen Y.H.、Tasi S.S.、Lin H.L.、Huang C.K.、Lin H H.(2009)。急诊部菌血症患者死亡的危险因素。阿卡德。急救医学。16,749–75510.1111/j.1553-2712.2009.00468.x[内政部] [公共医学] [谷歌学者]
-
Loike J.D.、el Khoury J.、Cao L.、Richards C.P.、Rascoff H.、Mandeville J.T.、Maxfield F.R.、Silverstein S.C.(1995)。纤维蛋白调节中性粒细胞迁移,以响应白细胞介素8、白三烯B4、肿瘤坏死因子和甲酰-甲亚砜基-亮氨酸-苯丙氨酸。实验医学学报1811763–177210.1084/jem.181.5.1763[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Ma L.J.、Mao S.L.、Taylor K.L.、Kanjanabuch T.、Guan Y.、Zhang Y.、Brown N.J.,Swift L.、McGuinness O.P.、Wasserman D.H.、Vaughan D.E.、Fogo A.B.(2004)。预防缺乏纤溶酶原激活物抑制剂1的小鼠肥胖和胰岛素抵抗。糖尿病53、336–34610.2337/糖尿病。53.2.336[内政部] [公共医学] [谷歌学者]
-
Ma T.Y.、Nguyen D.、Bui V.、Ngueen H.、Hoa N.(1999)。乙醇对肠上皮紧密连接屏障的调节。美国生理学杂志。276、G965–G974[内政部] [公共医学] [谷歌学者]
-
Marques-Vida P.、Cambou J.P.、Nicaud V.、Luc G.、Evans A.、Arveier D.、Bingham A.、Cambien F.(1995)。法国和北爱尔兰的心血管危险因素和饮酒。动脉粥样硬化115、225–23210.1016/0021-9150(94)05517-M[内政部] [公共医学] [谷歌学者]
-
Marshall L.J.、Ramdin L.S.、Brooks T.、DPhil P.C.、Shute J.K.(2003)。纤溶酶原激活物抑制剂-1通过抑制内皮细胞IL-8/硫酸乙酰肝素/syndecan-1复合物的组成性脱落,支持IL-8介导的中性粒细胞跨内皮细胞迁移。免疫学杂志。171, 2057–2065[内政部] [公共医学] [谷歌学者]
-
Mathurin P.、Deng Q.G.、Keshavarzian A.、Choudhary S.、Holmes E.W.、Tsukamoto H.(2000)。肠内内毒素加重大鼠酒精性肝损伤。肝病学321008–101710.1053/jhep.2000.19621[内政部] [公共医学] [谷歌学者]
-
麦克莱恩·C·J、科恩·D·A(1989)。酒精性肝炎患者单核细胞产生肿瘤坏死因子增加。肝病学9、349–35110.1002/hep.1840090302[内政部] [公共医学] [谷歌学者]
-
McKim S.E.、Gabele E.、Isayama F.、Lambert J.C.、Tucker L.M.、Wheeler M.D.、Connor H.D.、Mason R.P.、Doll M.A.、Hein D.W.、Arteel G.E.(2003)。酒精诱导肝损伤需要诱导型一氧化氮合酶:对敲除小鼠的研究。胃肠病学125、1834–184410.1053/j.gastro.2003.08.030[内政部] [公共医学] [谷歌学者]
-
Mukamal K.J.、Jadhav P.P.、D’Agostino R.B.、Massaro J.M.、Mittleman M.A.、Lipinska I.、Sutherland P.A.、Matheney T.、Levy D.、Wilson P.W.、Ellison R.C.、Silbershatz H.、Muller J.E.、Tofler G.H.(2001)。饮酒和止血因素:弗雷明翰后代队列分析。循环104、1367–137310.1161/hc3701.096067[内政部] [公共医学] [谷歌学者]
-
Nachiappan V.、Curtiss D.、Corkey B.E.、Kilpatrick L.(1994年)。细胞因子抑制离体大鼠肝细胞中脂肪酸氧化:TNF、IL-6和IL-1之间的协同作用。休克1, 123–12910.1097/00024382-199402000-00007[内政部] [公共医学] [谷歌学者]
-
Nanji A.A.,Khettry U.,Sadrzadeh S.M.H.(1994)。乳酸杆菌喂食可以降低实验性酒精性肝病的内毒素血症和严重程度。程序。Soc.实验生物学。医学。205, 243–247[内政部] [公共医学] [谷歌学者]
-
Nolan J.P.(2010)。肠内毒素在肝损伤中的作用:一段漫长的历史。肝病学52、1829–183510.1002/hep.23917[内政部] [公共医学] [谷歌学者]
-
Ontko J.A.(1973)。乙醇对离体肝细胞中游离脂肪酸代谢的影响。《脂质研究杂志》。第14页,78–86页[公共医学] [谷歌学者]
-
Pruett S.B.、Zheng Q.、Fan R.、Matthews K.、Schwab C.(2004)。乙醇抑制通过Toll样受体诱导的细胞因子反应以及对大肠杆菌在一个狂饮的老鼠模型中。酒33、147–15510.1016/S0741-8329(04)00102-8[内政部] [公共医学] [谷歌学者]
-
Purohit V.、Bode J.C.、Bode C.、Brenner D.A.、Choudhry M.A.、Hamilton F.、Kang Y.J.、Keshavarzian A.、Rao R.、Sartor R.B.、Swanson C.、Turner J.R.(2008)。酒精、肠道细菌生长、肠道对内毒素的渗透性和医学后果:研讨会摘要。酒42、349–36110.1016/j.alcohol.2008.03.131[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Qi J.、Goralnick S.、Kreutzer D.L.(1997)。纤维蛋白对人血管内皮细胞白细胞介素-8基因表达的调节。血液90, 3595–3602[公共医学] [谷歌学者]
-
Radek K.A.、Kovacs E.J.、Dipietro L.A.(2012年)。伤口愈合期间基质蛋白水解活性:急性乙醇暴露的调节。酒精。临床。实验结果。311045–105210.1111/j.1530-0277.2007.00386.x[内政部] [公共医学] [谷歌学者]
-
Radek K.A.、Matthies A.M.、Burns A.L.、Heinrich S.A.、Kovacs E.J.、Dipietro L.A.(2005)。急性乙醇暴露会损害血管生成和伤口愈合的增殖阶段。美国生理学杂志。心脏循环。生理学。289、1084–109010.1152/ajpheart.00080.2005[内政部] [公共医学] [谷歌学者]
-
Rivara F.P.、Jurkovich G.J.、Gurney J.G.、Seguin D.、Fligner C.L.、Ries R.、Raisys V.A.、Copass M.(1993)。创伤患者急性和慢性酒精滥用的程度。架构(architecture)。外科学。128、907–91210.1001/archsurg.1993.01420200081015[内政部] [公共医学] [谷歌学者]
-
Sato Y.、Tsuboi R.、Lyons R.、Moses H.、Rifkin D.B.(1990年)。内皮细胞与周细胞或平滑肌细胞共培养对潜伏TGF-β激活的表征:自我调节系统。《细胞生物学杂志》。111、757–76310.1083/jcb.111.41713[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Siegmund S.V.、Haas S.和Singer M.V.(2005年)。动物模型及其在胃肠道酒精研究中的结果。挖掘。数字化信息系统。23, 181–19410.1159/000090165[内政部] [公共医学] [谷歌学者]
-
药物滥用和心理健康服务管理局。(2011). 2010年全国药物使用与健康调查结果:全国调查结果摘要。医学博士Rockville:药物滥用和心理健康服务管理局[谷歌学者]
-
Tamai H.、Horie Y.、Kato S.、Yokoyama H.、Ishii H.(2002年)。长期乙醇喂养增强了大鼠肝脏对口服脂多糖的易感性。酒精。临床。实验结果。26,75S–80S10.1111/j.1530-0277.2002.tb02707.x[内政部] [公共医学] [谷歌学者]
-
Tamai H.、Kato S.、Horie Y.、Ohki E.、Yokoyama H.、Ishii H.(2000)。急性乙醇给药对大鼠内毒素肠吸收的影响。酒精。临床。实验结果。24、390–39410.1111/j.1530-0277.2000.tb04629.x[内政部] [公共医学] [谷歌学者]
-
Tran-Chang C.、Fasel-Felley J.、Pralong G.、Hofstetter J.R.、Bachmann F.、Kruithof E.K.(1989)。慢性酒精中毒或感染性肝炎所致肝功能缺陷患者的纤溶酶原激活剂和纤溶酶酶原激活物抑制剂。血栓。止血。62、651–653[公共医学] [谷歌学者]
-
Tsukamoto H.、Takei Y.、McClain C.J.、Joshi-Barve S.、Hill D.、Schmidt J.、Deaciuc I.、Barve S.和Colell A.、Garcia-Ruiz C.、Kapowitz N.、Fernandez-Checa J.C.、Yokoyama H.、Okamura Y.、Nakamura Ye.、Ishii H.、Chawla R.K.、Barve S、Joshi-Barve S、Watson W.H.、Nelson W.J.、Lin M.、Ohata M.、Motomura K.,Enomoto K.、Ikejima K.、Kitamura T.、。,Oide H.、Hirose M.、Bradford B.U.、Rivera C.A.、Kono H.、Peter S.、Yamashina S.、Konno A.、石川浩、清水H.、佐藤N.、瑟曼R.(2001)。酒精性肝病的肝脏是如何启动或致敏的?酒精。临床。实验结果。25、171S–181S10.1111/j.1530-0277.2001.tb02195.x[内政部] [公共医学] [谷歌学者]
- 美国卫生与公共服务部。(2000). 美国国会关于酒精和研究的第十次特别报告。
-
van Ginneken V.J.(2008)。盛宴和饥荒期间的肝脏肥大:一个进化悖论。医学假设70924–92810.1016/j.mehy.2007.10006[内政部] [公共医学] [谷歌学者]
-
Violi F.、Ferro D.、Basili S.、Quintarelli C.、Saliola M.、Alessandri C.、Cordova C.、Balsano F.(1992)。高纤溶增加晚期肝硬化患者胃肠道出血的风险。肝病学15、672–67610.1002/hep.1840150420[内政部] [公共医学] [谷歌学者]
-
Weschsler H.,Austin S.B.(1998)。酗酒:五四衡量标准。J.Stud.酒精59, 122–124[内政部] [公共医学] [谷歌学者]
-
世界卫生组织。(2011). 全球酒精与健康状况报告。日内瓦:世卫组织出版社[谷歌学者]
-
杨世清、林海珍、莱恩医学博士、克莱门斯医学博士、迪尔医学博士(1997)。肥胖增加内毒素肝损伤的敏感性:脂肪性肝炎发病机制的意义。程序。国家。阿卡德。科学。美国。94、2557–256210.1073/pnas.94.26.14524[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Yin M.、Bradford B.U.、Wheeler M.D.、Uesugi T.、Froh M.、Goyert S.M.、Thurman R.G.(2001)。减少cd14缺陷小鼠早期酒精诱导的肝损伤。免疫学杂志。166, 4737–4742[内政部] [公共医学] [谷歌学者]
-
钟伟、麦克莱恩·C·J、凯夫·M、康永杰、周泽(2010)。锌缺乏在酒精诱导的肠屏障功能障碍中的作用。美国生理学杂志。298,G625–G63310.1152/ajprenal.00289.2009[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
周华凤、陈华伟、威克莱S.A.、兰扎G.M.、范家通(2009)。α-β3靶向纳米疗法抑制小鼠炎症性关节炎。美国财务会计准则委员会J。232978–298510.1096/fj.09-129874[内政部] [PMC免费文章] [公共医学] [谷歌学者]

