线粒体代谢为生长中的癌细胞构建大分子提供了前体1,2在正常功能的肿瘤细胞线粒体中,葡萄糖和谷氨酰胺衍生碳的氧化代谢产生柠檬酸和乙酰辅酶A,用于脂质合成,这是肿瘤发生所必需的三然而,一些肿瘤的柠檬酸循环(CAC)或电子传递链(ETC)发生突变,导致线粒体正常氧化功能丧失4–7目前尚不清楚这些肿瘤细胞是如何产生大分子合成前体的。在这里,我们发现线粒体缺陷的肿瘤细胞使用谷氨酰胺依赖性还原羧化而不是氧化代谢作为柠檬酸盐形成的主要途径。该途径使用NADP的线粒体和细胞溶质亚型+/NADPH依赖的异柠檬酸脱氢酶和谷氨酰胺衍生柠檬酸盐的后续代谢为脂质合成提供乙酰辅酶A,并提供生成剩余CAC代谢物和相关大分子前体所需的4-碳中间体。这种还原性的谷氨酰胺依赖性途径是快速生长的恶性细胞中的主要代谢模式,这些恶性细胞中含有ETC复合物I或复合物III的突变,患者衍生的肾癌细胞中含有富马酸水合酶(FH)的突变,以及正常线粒体受到急性药物ETC抑制的细胞中。我们的发现揭示了多功能谷氨酰胺依赖性途径的新诱导,该途径逆转了典型CAC的许多反应,支持肿瘤细胞生长,并解释了细胞如何在线粒体代谢受损的情况下生成大量CAC中间产物。
我们首先研究了ETC复合物III(细胞色素b-c1复合物)中含有或缺乏功能缺失突变的等基因143B人骨肉瘤细胞的代谢。这些细胞系是通过消耗143B线粒体DNA(mtDNA)并重新填充野生型mtDNA或编码细胞色素b基因中含有移码突变的mtDNA而产生的(cytb公司),一种重要的复合物III成分8尽管突变体缺乏呼吸和复合III功能8,均为野生型(143B重量)和cytb公司-突变型(143Bcytb公司)细胞在软琼脂中形成菌落9并以可比的速度增长(补充图1a)使这些细胞成为研究线粒体功能障碍期间生长的良好模型。这两种细胞系都有可检测到的CAC中间产物,尽管在143B中柠檬酸盐含量较少,琥珀酸盐含量明显较多cytb公司单元格(补充图1b). 正如预期的那样,对于氧化磷酸化缺陷的细胞,143Bcytb公司细胞的葡萄糖消耗和乳酸生成高于143B重量细胞,表明代谢向有氧糖酵解转变(图1a). 为了确定cytb公司葡萄糖代谢命运突变,我们在含有D[U的培养基中培养这两种细胞系-13C] 葡萄糖和测量值13质谱法富集细胞内代谢物。143B年重量细胞,大多数柠檬酸分子含有葡萄糖衍生的13C类(图1b). 柠檬酸m+2是葡萄糖衍生丙酮酸通过丙酮酸脱氢酶(PDH)氧化脱羧形成[1,2-13C] 乙酰辅酶A,然后与未标记的草酰乙酸(OAA)缩合。绕CAC一圈处理柠檬酸盐m+2产生柠檬酸盐m+4(补充图2). 143B年cytb公司细胞,大多数柠檬酸盐不含葡萄糖碳(m+0),表明PDH对乙酰辅酶A的贡献受到抑制(图1b). 富马酸和苹果酸m+2也降低(补充图1c、d).
图1。缺乏ETC复合物III活性的癌细胞谷氨酰胺代谢的还原途径。
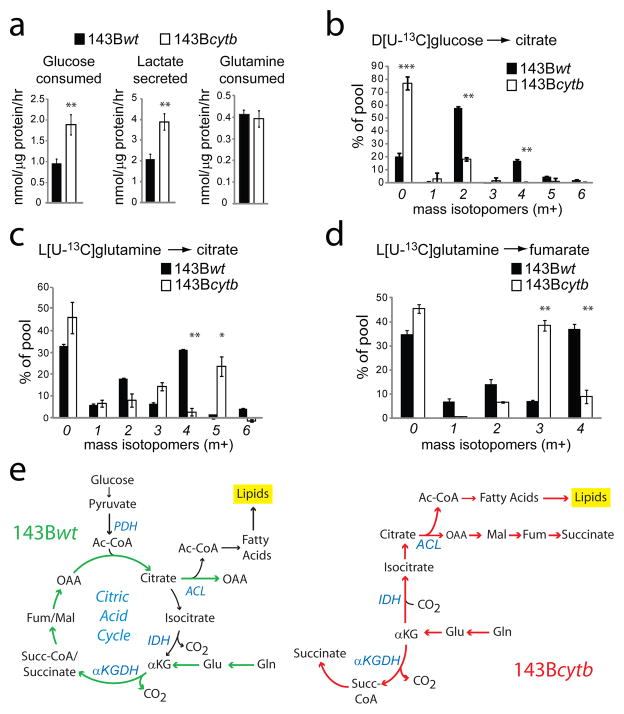
一143B年的葡萄糖利用、乳酸分泌和谷氨酰胺利用重量和143Bcytb公司细胞。b、,用D[U培养的细胞中柠檬酸盐的质量等位异构体分析-13C] 葡萄糖和未标记的谷氨酰胺。c、 日期:,L[U培养细胞中柠檬酸和富马酸的质量同位素分析-13C] 谷氨酰胺和未标记葡萄糖。数据是三种独立培养物的平均±S.D.。*p<0.05;**p<0.005,学生t检验。e、,143B年谷氨酰胺代谢示意图重量和143Bcytb公司细胞。彩色箭头沿着谷氨酰胺衍生碳的路径。缩写:Ac-CoA,乙酰-CoA;草酰乙酸;谷氨酰胺;谷氨酸;αKG,α-酮戊二酸;琥珀酰辅酶A;富马酸;苹果酸;PDH、丙酮酸脱氢酶;ACL、ATP-柠檬酸裂解酶;异柠檬酸脱氢酶;α-KGDH,α-酮戊二酸脱氢酶。
谷氨酰胺是癌细胞的主要呼吸底物,为生长提供能量和补充碳三,10.均为143B重量和143Bcytb公司细胞需要谷氨酰胺来形成菌落,这意味着谷氨酰胺在细胞生长中具有呼吸依赖性功能9.143亿cytb公司和143B重量细胞以相似的速度消耗谷氨酰胺(图1a). 我们用L[U培养了两种细胞系-13C] 谷氨酰胺定义谷氨酰胺的代谢命运。类似于其他谷氨酰胺依赖性癌细胞11,143B年重量细胞使用谷氨酰胺作为主要的回补前体,产生大量富马酸、苹果酸和柠檬酸m+4(图1c、d;补充图1e和3,顶部)。相比之下,143Bcytb公司细胞只产生微量柠檬酸盐m+4。相反,他们通过谷氨酰胺衍生的α-酮戊二酸的还原羧基化生成柠檬酸盐m+5(图1c). 该反应涉及通过异柠檬酸脱氢酶(IDH)添加未标记的碳,与典型氧化CAC相反12.通过ATP-柠檬酸裂解酶裂解柠檬酸m+5,然后生成乙酰CoA m+2和OAA m+3,未标记的碳保留在OAA上(补充图3,底部)。143B中其他CAC代谢物的检测cytb公司细胞显示,它们在柠檬酸盐m+5和OAA m+3的下游形成。这些细胞含有丰富的苹果酸和富马酸m+3,这在143B年基本上不存在重量单元格(图1d和补充图1e). 因此,这些代谢物是由还原性而非氧化性谷氨酰胺代谢产生的(图1e). 丁二酸是在143B年通过还原和氧化谷氨酰胺代谢形成的cytb公司单元格(补充图1f). 143B也观察到谷氨酰胺依赖性还原羧基化cytb公司在含有完整氨基酸的培养基中培养的细胞(补充图4)和ETC复合物I基因缺陷细胞(补充图5).
为了确定三种IDH亚型中哪一种有助于还原羧基化,我们使用RNA干扰(RNAi)来沉默印尼盾1,IDH2公司和印尼盾3143B年cytb公司细胞和测量的谷氨酰胺来源13柠檬酸盐中的C标记。沉默任何一个NADP+/NADPH依赖性IDH亚型,印尼盾1或IDH2公司,在用L[U培养的细胞中减少柠檬酸盐m+5-13C] 谷氨酰胺和沉默同时进一步降低柠檬酸盐m+5(图2a、b). 谷氨酰胺转化为谷氨酸不受影响(图2b). 沉默印尼盾3,线粒体NAD+/NADH依赖性亚型,对标记无影响(图2a,b). 因此143B中的还原羧基化cytb公司单元格涉及印尼盾1和IDH2公司,但不是印尼盾3因为IDH1和IDH2蛋白主要定位于两种细胞系的细胞质和线粒体(补充图6),数据表明143B年的还原羧基化cytb公司细胞可能出现在任何一个隔室中。
图2。NADP公司+/异柠檬酸脱氢酶依赖NADPH的亚型有助于还原羧基化。
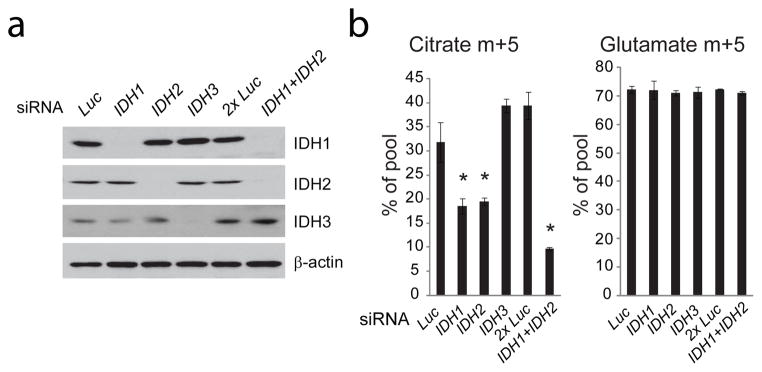
a、,143B异柠檬酸脱氢酶(IDH)蛋白的瞬时沉默cytb公司针对具有短干扰RNA(siRNAs)的细胞印尼盾1,IDH2公司或印尼盾3.一种针对荧光素酶的siRNA(吕克)用作阴性对照。什么时候?印尼盾1和IDH2公司siRNA同时转染(印尼盾1+IDH2公司),转染相同数量的纳米Luc siRNA(2倍吕克)用作阴性对照。b、,143B年柠檬酸和谷氨酸的质量同位素分析cytb公司L[U培养的细胞-13C] IDH亚型沉默后的谷氨酰胺。数据是三种独立文化的平均±S.D*p<0.05,学生t检验。
为了确定生长是否需要IDH1和IDH2,我们使用慢病毒RNAi稳定地沉默了每种酶。143B年重量细胞,沉默任一亚型降低细胞生长(补充图7),与其他线粒体正常的癌细胞的研究结果一致13143B年cytb公司细胞,当任一亚型沉默时,也会出现类似的生长减少(补充图7). 因此,IDH1和IDH2都支持使用谷氨酰胺依赖性还原羧基化的细胞生长。
接下来,我们确定还原性谷氨酰胺代谢是否有助于脂肪生成,这是癌细胞生长所需的生物合成活动。都是143B重量和143B胞苷细胞产生脂肪酸从头开始,如公司所示三H(H)2O转化为脂质(图3a). 然后用14C-标记的脂肪生成底物用于检查碳对脂肪生成的偏好(图3a). 这两种细胞系都有来自外源性乙酸盐的完整脂肪生成途径。143B中的增强标签cytb公司细胞可能是由于PDH抑制,细胞内2-碳池的比活性增加所致。来自143B的脂质cytb公司细胞中葡萄糖的放射性很低,但谷氨酰胺的放射性很高,这表明这些营养物质提供脂肪生成的程度发生了变化。为了量化葡萄糖和谷氨酰胺对脂肪生成乙酰辅酶A的贡献,将细胞培养在含有D[U-13C] 葡萄糖和未标记的谷氨酰胺,或L[U-13C] 谷氨酰胺和未标记葡萄糖。提取的脂质通过13C核磁共振。因为脂肪酸是通过连续添加2个碳单位合成的13C类-13ω-1和ω-2碳(进入不同2-碳单位的脂肪酰基链)之间的C耦合可以计算13产脂乙酰辅酶A中的C富集三.在ω-1多重态中,13C类-13C联轴器在两个13连续添加C-标记的2-碳单元,而一个标记的单元和一个未标记的单元产生双重(图3b). 因此13脂肪生成乙酰辅酶A中的C标记与三重态对ω-1多重态总面积的贡献成正比。143B年重量用D[U培养的细胞-13C] ω-1共振以三联体为主,占总面积的65%(图3c,左)。谷氨酰胺是脂肪酰基碳的次要来源,当细胞与L[U培养时,三联体只占15%的面积-13C] 谷氨酰胺(图3d,左)。这种模式在143B年被逆转cytb公司细胞,谷氨酰胺提供67%的脂肪生成碳(图3c、d,右;补充图8a). 因此,还原性谷氨酰胺代谢是143B年新合成脂质中脂肪酸生成的主要途径cytb公司增长。
图3。谷氨酰胺是缺乏氧化CAC功能的细胞中主要的成脂前体。
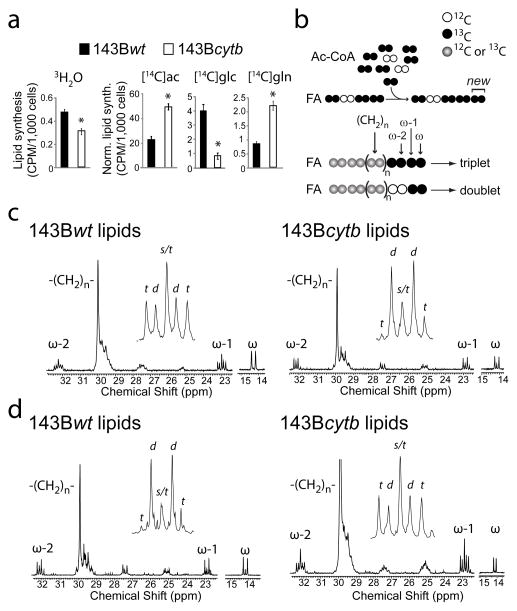
a、,细胞在含有三H(H)2O(21%体积),或14醋酸盐(ac,5μCi)、葡萄糖(glc,10μCi。对脂质进行分析三H或14闪烁计数法测定C含量。在14C实验,将每千个细胞的原始计数标准化为使用三H(H)2O.*p<0.05,学生t检验。b、,示意图概述了从乙酰辅酶A(Ac-CoA)合成脂肪酸(FA)以及ω-1处三重态和双态的来源13C核磁共振波谱。c、 日期:,
13D[U标记脂质的C NMR-13C] 葡萄糖(c(c))或L[U-13C] 谷氨酰胺(d日). 插图是ω-1共振的展开式。缩写:s,singlet;d、 双股;t、 三胞胎。
虽然大多数癌细胞都有功能性线粒体,但人类肿瘤的一个子集含有损害线粒体代谢的突变。CAC酶琥珀酸脱氢酶功能所需的基因发生两类主要突变(SDHA-D公司,SDHAF2型)或富马酸水合酶(跳频)14为了检测含有这些突变的肿瘤细胞中是否发生谷氨酰胺依赖性还原羧基化,我们研究了遗传性平滑肌瘤病肾细胞癌综合征患者肾肿瘤中的UOK262细胞。在这种综合征中,受影响的个体会遗传一种功能丧失跳频等位基因及其肿瘤显示杂合性缺失跳频轨迹15.UOK262细胞呼吸缺陷,缺乏FH活性15在含有半乳糖和谷氨酰胺的无葡萄糖培养基中培养失败(图4a),一种迫使细胞利用线粒体代谢产生ATP的情况16缺乏FH活性使谷氨酰胺无法用作传统氧化CAC中的碳源。然而,UOK262细胞需要谷氨酰胺才能增殖(图4b). 用D[U培养这些细胞-13C] 葡萄糖产生的质量同位素数据与143B相似cytb公司细胞,柠檬酸盐m+2的形成最少(图4c,左)。在用L[U培养的细胞中-13C] 柠檬酸最丰富的质量同位素谷氨酰胺是由还原羧基化形成的m+5(图4c,左)。富马酸主要通过氧化谷氨酰胺代谢形成,产生富马酸m+4(图4c,中间)。然而,大部分苹果酸池(m+3)是通过柠檬酸盐分解和随后的还原代谢形成的(图4c,右侧)。使用D[U标记脂质-13C] 葡萄糖或L[U-13C] 谷氨酰胺显示,就像143Bcytb公司细胞,谷氨酰胺是脂肪酸合成的主要碳源(补充图8b). 因此,这些癌细胞通过两条不同的途径产生CAC中间体,以补偿FH酶活性的损失:谷氨酰胺氧化代谢为富马酸,谷氨酰胺依赖性还原羧基化生成柠檬酸盐,乙酰辅酶A生成脂肪,以及剩余的4-碳CAC中间体。
图4。缺乏富马酸水合酶的肾癌细胞中谷氨酰胺依赖性还原羧基化,以及线粒体正常的肿瘤细胞中ETC抑制期间。
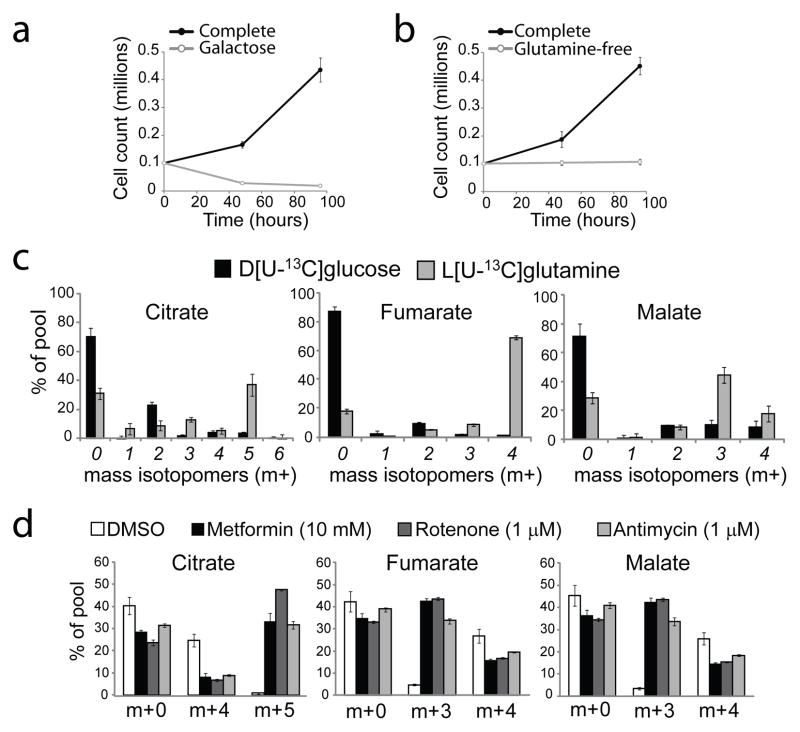
UOK262肾癌细胞在葡萄糖被半乳糖取代的完全培养基中的生长(一)在缺乏谷氨酰胺的中等水平(b条).c、,在含有D[U的培养基中培养的UOK262细胞柠檬酸、富马酸和苹果酸的质量同位素分析-13C] 葡萄糖和未标记的谷氨酰胺,或L[U-13C] 谷氨酰胺和未标记葡萄糖。日期:,143B年柠檬酸、富马酸和苹果酸的质量同位素分析重量在含有未标记葡萄糖和L[U的培养基中培养的细胞-13C] 谷氨酰胺。在用载体(DMSO)、二甲双胍、鱼藤酮或抗霉素处理细胞的同时,引入标记的谷氨酰胺。6小时后提取代谢物。在所有实验中,数据均为三种独立培养物的平均±S.D。
最后,我们测试了谷氨酰胺依赖性还原羧基化是否是线粒体代谢永久性异常细胞特有的。1430亿重量对细胞和转化的小鼠胚胎成纤维细胞(MEFs)进行急性ETC抑制。暴露于复合物III(抗霉素)或复合物I(鱼藤酮)抑制剂诱导谷氨酰胺从氧化代谢转变为还原代谢(图4d,补充图9a). ETC抑制一小时后,还原羧基化增强明显,并且不能通过蛋白质合成抑制剂环己酰亚胺的预处理消除(补充图9b,c). 二甲双胍,一种通过对复合物I的作用抑制呼吸的抗糖尿病药物17高剂量时还诱导谷氨酰胺依赖性还原羧基化(图4d,补充图9a). 有趣的是,模拟急性二甲双胍中毒和乳酸酸中毒患者血浆浓度的剂量可诱导谷氨酰胺还原代谢(补充图10). 这些数据表明,谷氨酰胺依赖性还原羧基化是线粒体代谢受损的常见细胞反应。
谷氨酰胺依赖性还原羧基化以前被描述为哺乳动物细胞中异柠檬酸盐/柠檬酸盐和脂肪碳的次要来源12,13,18–20.英寸恶性疟原虫是一种呼吸受限的单细胞原生动物,该途径是柠檬酸和乙酰基单位的主要来源21我们表明,该途径也可以作为主要的代谢策略来产生用于癌细胞生长的脂质,并且可以在ETC抑制期间被急性诱导。我们推测,ETC损伤导致线粒体氧化还原比率紊乱,从而降低NAD,从而刺激还原羧基化+/NADH比率和使CAC的氧化功能效率降低。NAD(P)-转氢酶可以部分消散这一比例,将还原当量从NADH转移到NADPH,而这反过来又可能通过IDH1和IDH2驱动NADPH依赖的还原羧基化。CAC的这种重定向扩展了谷氨酰胺代谢在癌细胞生长中的多功能性。在线粒体正常的细胞中,谷氨酰胺氧化常常是回补的主要来源,为CAC中间产物库提供大部分碳,并促进线粒体中大分子前体的生成22谷氨酰胺依赖性还原羧基化使谷氨酰胺即使在线粒体代谢受损时也能保持其作为促生长营养素的地位。因此,在恶性转化过程中,获得损害线粒体功能并刺激有氧糖酵解的体细胞突变(Warburg效应)并不会降低谷氨酰胺代谢或线粒体本身对肿瘤细胞增殖的重要性。
方法总结
1430亿重量和143Bcytb公司和UOK262细胞按所述进行培养9,15MEF在含有10%胎牛血清(FBS,Hyclone)、青霉素/链霉素和6mM L-谷氨酰胺的Dulbecco改良鹰培养基(DMEM)中培养。通过胰酶消化和血细胞仪计数监测亚融合培养物中的细胞生长。如前所述测量葡萄糖、乳酸和谷氨酰胺23在DMEM中用10%透析的FBS补充15mM D-[U进行同位素标记-13C] 葡萄糖或2mM L-[U-13C] 谷氨酰胺。使用安捷伦6970气相色谱仪和安捷伦5973质量选择检测器分析水代谢物。为了确定样品中的相对代谢物丰度,将相关代谢物的总离子电流峰值面积与内标物(2-氧代丁酸)的总离子流峰值面积进行比较,并对蛋白质含量进行归一化。分析13碳富集和质量同位素分布按照公布的方法进行11,24对于放射性同位素脂质合成分析,细胞在补充有三H(H)2O或14C示踪剂(Perkin Elmer)。按照说明提取并分析脂质三对于使用NMR的脂质合成实验,将细胞接种在10cm培养皿中,并使其在含有D[U的培养基中指数增殖-13C] 葡萄糖和未标记的谷氨酰胺,或未标记的葡萄糖和L[U-13C] 直到得到两个15厘米的盘子。然后通过以下方法提取和分析脂质13如上所述的核磁共振三使用商业化小干扰RNA(Thermo)进行基因沉默印尼盾1,IDH2公司或印尼盾3用DharmaFECT转染试剂(Thermo)转染细胞,72小时后用带有商业抗体的western blots检测蛋白质丰度(Santa Cruz Biotechnology)。细胞与L[U孵育-13C] 谷氨酰胺提取前两小时进行代谢物的质量同位素分布分析。稳定沉默使用来自Mission shRNA pLKO.1-puro文库(Sigma)的慢病毒shRNA。更多详细信息请参阅在线方法。
致谢
Kosaku Uyeda和DeBerardinis和Chandel实验室的成员对数据进行了关键评估,Jessica Sudderth和Chendong Yang提供了实验援助。我们感谢卡洛斯·莫莱斯和I.F.M.de Coo为143B重量和143Bcytb公司CCL16-B2细胞系为Immo E.Schefler,CCL16-NDI1细胞系为Takao Yagi。这项工作得到了N.I.H.(K08DK072565、R01CA157996和RR02584)、德克萨斯州癌症预防与研究所(CPRIT,HIRP100437)和罗伯特·韦尔奇基金会(1733)向R.J.D.提供的资助;由N.I.H.(R01CA123067)、LUNGevity基金会和芝加哥大都会呼吸健康协会召集的独立肺部健康组织联盟向N.S.C.提交;并通过N.I.H.拨款(DK078933)支付给E.S.J。这项工作还得到了国家癌症研究中心N.I.H.的内部研究计划的支持。N.I.H.培训拨款支持A.R.M.(5T32GM083831)、W.W.W.(T32CA009560)和L.B.S.(T32GM008061)。T.C.得到了CPRIT培训拨款的支持。
脚注
作者贡献。A.R.M.、W.W.W.、N.S.C.和R.J.D.设计了这项研究。A.R.M.、W.W.W.、L.B.S.、E.S.J.、T.C.和P-H.C.进行了实验。A.R.M.、W.W.W.、L.B.S.、E.S.J.、P-S.C.、T.C.、N.S.C.和R.J.D.分析了数据。Y.Y.和W.M.L.提供了FH缺陷(UOK262)细胞。A.R.M.、N.S.C.和R.J.D.撰写了这篇论文。
工具书类
-
1Vander Heiden MG、Cantley LC、Thompson CB。了解Warburg效应:细胞增殖的代谢需求。科学。2009;324:1029–1033. doi:10.1126/science.1160809。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
2Gao P等。c-Myc抑制miR-23a/b可增强线粒体谷氨酰胺酶的表达和谷氨酰胺代谢。自然。2009;458:762–765. doi:10.1038/nature07823。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
三。DeBerardinis RJ等。除需氧糖酵解外:转化细胞可以参与谷氨酰胺代谢,超过蛋白质和核苷酸合成的需要。美国国家科学院院刊2007;104:19345–19350. doi:10.1073/pnas.0709747104。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
4Linehan WM,Srinivasan R,Schmidt LS。肾癌的遗传基础:一种代谢性疾病。Nat Rev Urol公司。2010;7:277–285. doi:10.1038/nurol.2010.47。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
5Baysal BE等。遗传性副神经节瘤中线粒体复合物II基因SDHD的突变。科学。2000;287:848–851. doi:10.1126/science.287.5454.848。[内政部] [公共医学] [谷歌学者]
-
6Tomlinson IP等人。FH的种系突变易导致显性遗传的子宫肌瘤、皮肤平滑肌瘤和乳头状肾细胞癌。自然遗传学。2002;30:406–410. doi:10.1038/ng849。[内政部] [公共医学] [谷歌学者]
-
7郝HX等。琥珀酸脱氢酶黄变所需的基因SDH5在副神经节瘤中发生突变。科学。2009;325:1139–1142. doi:10.1126/science.1175689。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
8Rana M,de Coo I,Diaz F,Smeets H,Moraes CT。帕金森综合征患者胞外细胞色素b基因缺失与复合物III组装受损和自由基生成增加有关。Ann Neurol公司。2000;48:774–781.[公共医学] [谷歌学者]
-
9Weinberg F等。线粒体代谢和活性氧生成对Kras介导的致瘤性至关重要。美国国家科学院院刊2010;107:8788–8793. doi:10.1073/pnas.1003428107。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
10Reitzer LJ、Wice BM、Kennell D。谷氨酰胺而非糖是培养HeLa细胞的主要能量来源的证据。生物化学杂志。1979;254:2669–2676.[公共医学] [谷歌学者]
-
11Cheng T等。丙酮酸羧化酶是谷氨酰胺诱导肿瘤细胞依赖性生长所必需的。美国国家科学院院刊2011;108:8674–8679. doi:10.1073/pnas.1016627108。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
12Des Rosiers C,Fernandez CA,David F,Brunengraber H。灌注大鼠肝脏线粒体异柠檬酸脱氢酶反应的可逆性。柠檬酸循环中间产物同位素分析的证据。生物化学杂志。1994;269:27179–27182.[公共医学] [谷歌学者]
-
13.Ward PS等。白血病相关IDH1和IDH2突变的共同特征是将α-酮戊二酸转化为2-羟基戊二酸的新形态酶活性。癌细胞。2010;17:225–234. doi:10.1016/j.ccr.2010.01.020。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
14Frezza C、Pollard PJ、Gottlieb E.癌症中的先天性和后天性代谢缺陷。《分子医学杂志》2011;89:213–220. doi:10.1007/s00109-011-0728-4。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
15Yang Y,等。UOK 262细胞系,富马酸水合酶缺乏(FH-/FH-)遗传性平滑肌瘤病肾细胞癌:人类癌症中异常能量代谢途径的体内外模型。癌症基因-细胞基因。2010;196:45–55. doi:10.1016/j.cancergencyto.2009.08.018。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
16Rossignol R等人。能量底物调节癌细胞中的线粒体结构和氧化能力。癌症研究2004;64:985–993. doi:10.1158/008-5472.can-03-1101。[内政部] [公共医学] [谷歌学者]
-
17El-Mir MY等。二甲基双胍通过靶向呼吸链复合物的间接作用抑制细胞呼吸。生物化学杂志。2000;275:223–228. doi:10.1074/jbc.275.1.223。[内政部] [公共医学] [谷歌学者]
-
18Yoo H、Antoniewicz MR、Stephanopulos G、Kelleher JK。定量棕色脂肪细胞系中谷氨酰胺对脂质的还原羧化通量。生物化学杂志。2008;283:20621–20627. doi:10.1074/jbc。M706494200。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
19Comte B,Vincent G,Bouchard B,Benderdour M,Des Rosiers C.在常氧和缺血条件下通过心脏NADP(+)-异柠檬酸脱氢酶逆转流量。美国生理学杂志心脏循环生理学。2002;283:H1505–1514。doi:10.1152/ajpheart.00287.2002。[内政部] [公共医学] [谷歌学者]
-
20Lemons JM等。静止的成纤维细胞表现出高代谢活性。《公共科学图书馆·生物》。2010;8:e1000514。doi:10.1371/journal.pbio.1000514。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
21Olszewski KL等,恶性疟原虫的分支三羧酸代谢。自然。2010;466:774–778. doi:10.1038/nature09301。[内政部] [PMC免费文章] [公共医学] [谷歌学者] [已缩回]
-
22DeBerardinis RJ,Cheng T.Q的下一篇:谷氨酰胺在新陈代谢、细胞生物学和癌症中的多种功能。癌基因。2010;29:313–324. doi:10.1038/onc.2009.358。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
23.Yang C等。胶质母细胞瘤细胞需要谷氨酸脱氢酶才能在葡萄糖代谢或Akt信号受损时存活。2009年癌症研究;69:7986–7993. doi:10.1158/0008-5472.CAN-09-2266。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
24Fernandez CA、Des Rosiers C、Previs SF、David F、Brunengraber H。天然稳定同位素丰度的13C质量同位素分布校正。质谱学杂志。1996;31:255–262. doi:10.1002/(SICI)1096-9888(199603)31:3<255::AID-JMS290>3.0.CO;2-3.[内政部] [公共医学] [谷歌学者]
关联数据
本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。

