MicroRNAs是小分子(21–23-nt)的大家族4在整个后生动物物种中表现出高度结构和功能保守性的RNA。miRNAs最初由聚合酶II合成为长一级转录物,随后由Drosha RNase III内切酶加工成~70-nt干环前microRNAs(1)并通过exportin 5从细胞核中运输出来(2). Dicer在细胞质中进一步处理前microRNA,以产生最终的~22-nt成熟miRNA(三). miRNA与靶mRNA的结合具有完全或接近完全互补性,会导致mRNA降解,而不完全互补性通常会导致翻译抑制。人们认为,miRNAs 5′端的7-8nt(称为种子序列)对有效靶向至关重要。
miRNAs参与调节复杂的生理过程,如胚胎发生(4),器官发育(5)和肿瘤发生(6,7). 然而,绝大多数miRNAs的功能作用仍然未知。最近,一些研究小组使用了多种模型系统来识别不同的miRNAs作为转移的启动子或抑制子(8–12). 尽管这些研究明确表明这些miRNAs与转移有关,但尚不清楚这些miRNAs在多步骤转移过程中调节哪些步骤。在本研究中,我们试图确定miRNAs在调节转移起始步骤上皮间质转化(EMT)中的作用。应用在NMuMG细胞(正常小鼠乳腺上皮细胞)中诱导EMT的经典模型系统,我们发现miR-200家族的成员在EMT期间被显著抑制,这表明他们是EMT的抑制物。我们进一步发现,miR-200家族成员通过直接靶向E-钙粘蛋白的表达来抑制EMTZEB1号机组和ZEB2型此外,miR-200家族在4TO7乳腺癌细胞中的异位表达(表达这些miRNAs的低内源性水平)通过上调E-cadherin的表达诱导了间质-上皮转变,并抑制了这些肿瘤细胞的迁移。这些结果有力地表明了miR-200家族miRNA在抑制上皮-间充质转化和癌症进展中的重要作用。
实验程序
细胞系与细胞培养-NMuMG、4TO7和HeLa细胞保存在补充有10%胎牛血清的Dulbecco改良Eagle's培养基中。NMuMG培养基进一步补充10μg/ml胰岛素(Sigma)。
RNA提取和实时定量PCR-总RNA使用miR公司Vana miRNA分离试剂盒(Ambion)。对于miRNA分析,成熟的miRNA被反转录,并使用塔克人类microRNA分析(应用生物系统)。所有数据均归一化为U6表达。对于mRNA分析,在ABI 7900HT系列PCR机器Applied Biosystems上使用Power SYBR®绿色PCR主混合液(Applied生物系统)进行实时PCR,数据标准化为GAPDH公司表达并进一步归一化为阴性对照,除非另有说明。
ZEB1和ZEB2 3′-UTR荧光素酶报告分析-两者的3′-UTRZEB1号机组和ZEB2型从NMuMG细胞提取的基因组DNA中进行PCR扩增。PCR引物用于扩增斑马13′-UTR包括5′-AAAA TCCGGGTGTGCTGTGA-3′(正向)和5′-AACTGCTTTTCTACTCTG-3′(反向),而用于扩增Zeb2 3′-UTR的引物包括5′-CAGTTCAGCAAGAGAGAGT-3′(正向,5′-TTCGATCATTTC-3′(逆向)。扩增的3′-UTR被克隆到pMIR-REPORT™(Ambion)中萤火虫荧光素酶编码区的下游。转染前24小时将HeLa或4TO7细胞接种在24孔板中。第二天,200 ng报告质粒和200 ng对照雷尼利亚-荧光素酶质粒使用Lipofectamine 2000(Invitrogen)联合转染。转染24小时后收集细胞,并使用Glomax 96光度计(Promega)检测荧光素酶活性。为了评估miRNA对报告活性的影响,50 p米共转染合成前体miRNAs(前miRs)(Ambion)。所有实验均进行了三次。
免疫印迹分析-对于EMT分析,将NMuMG细胞接种在6孔板中,第二天,将合成的miRNAs单独或成簇转染细胞。转染24h后,用200p处理细胞米TGFβ1(R&D系统)48 h。细胞在预冷裂解缓冲液中裂解,每道装载10μg上清液蛋白,并通过SDS-聚丙烯酰胺电泳进行分离。将蛋白质转移到硝化纤维素膜上,用小鼠抗E-钙粘蛋白(BD Biosciences)、小鼠抗N-钙粘蛋白或小鼠抗β-肌动蛋白(AbCam)进行阻断和探测。
免疫荧光-将NMuMG细胞转染到带有合成前microRNA(pre-miRs)的6孔板中。转染后4–6 h,将细胞分离并接种到放置在24孔板中的明胶涂层玻璃盖玻片上,并用重组TGFβ刺激48 h。4TO7细胞也进行了类似处理,但没有用TGFβ进行刺激。转染72h后,抽吸培养基,用冰镇甲醇固定10min,用0.2%Triton渗透3min,室温下用10%山羊血清封闭1h。在室温下用小鼠抗E-钙粘附素探针探测E-钙黏附素1h,然后在室温下使用罗丹明结合的山羊抗鼠二级抗体检测1h。随后使用Hoechst染料(1μg/ml)对细胞核进行染色。在蔡司显微镜上观察细胞,并使用Axiocam Icc3相机拍摄照片。
Transwell迁移分析-将前miRs转染4TO7细胞48小时,然后分离。1 × 105细胞重新悬浮在无血清培养基中,并放置在含有8-μm孔的插入物中。这些插入物被放置在含有血清的培养基中。播种后12h,抽吸含血清的培养基,将胰蛋白酶放入孔中,对通过孔的细胞进行胰蛋白酶消化,并使用血细胞仪进行计数。
结果
EMT期间miR-200家族miRNA的下调-为了研究EMT,我们使用了一个经典的模型系统:TGFβ1在NMuMG小鼠乳腺上皮细胞系中诱导EMT。在TGFβ治疗72小时内,NMuMG细胞经历了巨大的形态学变化,从致密的鹅卵石样上皮结构变为纤维母细胞样纺锤状细胞,同时细胞间黏附显著解体(图1A类). 这种形态转变伴随着E-cadherin的下调和膜定位的减少(图1,A类和B类)和N-钙粘蛋白上调(图1B类)增加了对膜的定位(图1A类). 这些形态和分子水平上的特征性变化表明NMuMG细胞中的EMT程序是成功的。
图1。
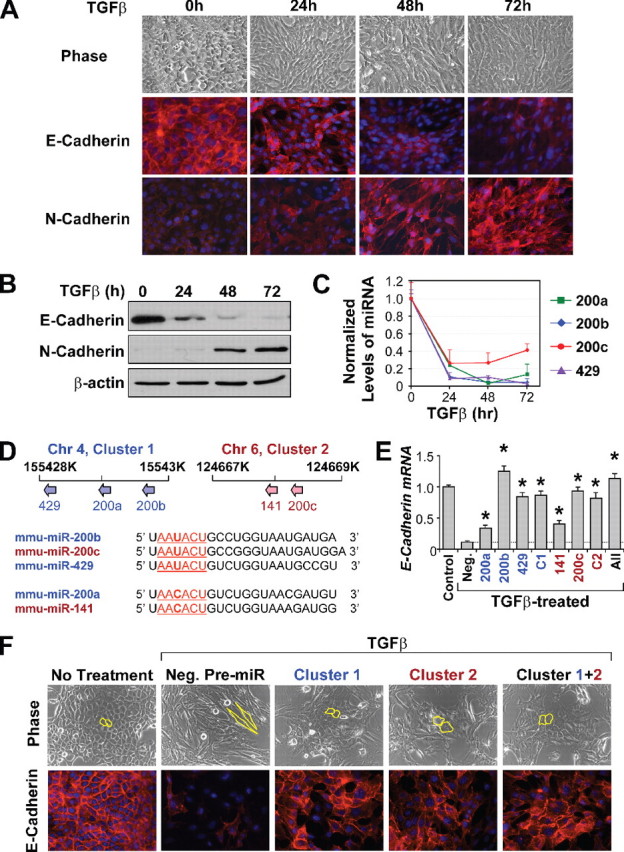
miR-200家族的过度表达阻碍EMT并上调E-cadherin的表达。 A类进行EMT的NMuMG细胞中E-cadherin和N-cadherin的相位对比图像和免疫荧光染色。B类Western blot分析证实了EMT期间E-cadherin的下调和N-cadherin上调。C类TGFβ处理的NMuMG细胞中miR-200家族水平的变化塔克Man-qRT-PCR并归一化为U6表达。数据为代表性时间过程实验的平均值,一式三份,以平均值±标准误差表示。天,上部面板,miR-200家族成员在小鼠基因组中的染色体位置示意图。下部面板miR-200家族成员的序列比对。代表其种子序列的核苷酸2–7为下划线的集群1中嵌入的.miR-200成员位于蓝色,而嵌入到集群2中的是红色.第4阶段,第4染色体。电子转化生长因子β处理的NMuMG细胞中E-钙粘蛋白表达的变化,以及单独转染miR-200成员的NMuMG细胞中E-cadherin的表达变化(C1类或指挥与控制),或全部(全部),通过实时PCR测量。将表达水平与未经TGFβ(对照)处理的细胞或转染阴性对照前miR的TGFβ处理细胞进行比较(阴性。). * 代表对与miR前对照组相比<0.05。F类在转染阴性对照pre-miR、簇1、簇2或同时转染这两个簇后,未经处理或经TGFβ处理的NMuMG细胞的相差显微镜和E-cadherin染色(集群1+2). 细胞形态概述如下黄色的.
两个miR-200家族miRNAs,miR-200b和miR-200c,以前与上皮细胞中E-cadherin的表达相关(13,14). 另外三种miRNAs,miR-200a、miR-141和miR-429,也属于miR-200家族。qRT-PCR分析显示,在TGFβ诱导的EMT过程中,NMuMG细胞中除miR-141外,所有miR-200家族miRNAs的表达均显著下调(图1C类). 在TGFβ暴露24小时内,每个miRNA的表达降低到预处理水平的30%以下。我们没有观察到NMuMG细胞中miR-141的可检测表达。
miR-200家族miRNAs被映射到小鼠基因组中两个单独的小于2000 bp的簇(图2天,上部面板). 第一个集群(集群1)包含miR-200a、miR-200b和miR-429,位于小鼠4号染色体上。对几个细胞系中miR-200a、miR-200b和miR-429的表达水平的分析表明,该簇的三个成员共同表达(第页=0.97,数据未显示),表明一个共同的启动子可能驱动所有三个miRNA的表达。第二个集群(集群2)由miR-200c和miR-141组成,位于6号染色体的500-bp区域。5个miR-200家族miRNAs包含非常相似的种子序列(图1天,下部面板). miR-200b、miR-200c和miR-429、AA的种子序列U型ACU与miR-200a和miR-141,AA的种子序列仅存在一个核苷酸的差异C类ACU公司。
图2。
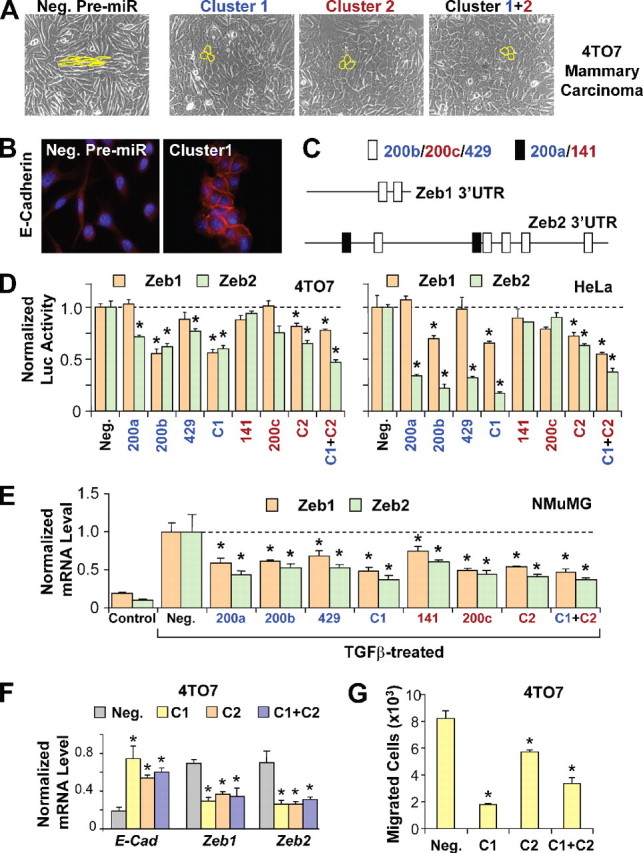
miR-200家族靶向转录抑制因子泽布1和ZEB2型增强E-cadherin表达并抑制迁移。 A类,转染阴性对照前miR的4TO7细胞的相差显微镜(阴性。)、集群1、集群2或同时存在两个集群(集群1+2). 细胞形态概述如下黄色的.B类转染阴性对照前miR或簇1的4TO7细胞的E-cadherin染色。C类,小鼠中推测的miR-200靶点示意图ZEB1号机组和ZEB2型3′-UTR。白色方框代表miR-200b/200c/429的目标位点,而黑匣子代表miR-200a/141的目标站点。天,荧光素酶报告子的归一化活性泽布1(棕色条)或ZEB2型(绿色条)4TO7中的3′-UTR(左侧面板)或HeLa细胞(右侧面板)在联合转染阴性对照pre-miR或miR-200成员单独存在的情况下,作为簇(C1类或指挥与控制)或两个群集(C1类+指挥与控制). 24小时后测量荧光素酶活性。数据为三倍的平均值±S.E.,显示为萤火虫与雷尼利亚荧光素酶活性。电子,的表达式级别ZEB1号机组和ZEB2型未经处理的NMuMG细胞(控制)或用TGFβ治疗,转染阴性对照pre-miR或miR-200家族成员,作为集群(C1类或指挥与控制),或两个群集(C1类+指挥与控制).F类E-cadherin表达的变化(钙黏蛋白),ZEB1号机组、和ZEB2型(标准化为GAPDH公司转染阴性对照前miR或簇1、簇2或两个簇的4TO7细胞。G公司,转染阴性对照前miR、簇1、簇2或这两个簇的4TO7细胞向含血清培养基迁移。数据为代表性实验中迁移的平均细胞数,一式三份,以平均值+S.E.*表示对< 0.05.
miR-200成员单独或联合过度表达抑制EMT并增强E-cadherin表达-由于miR-200家族中的miRNA在EMT期间同样下调,我们通过合成前miRs的短暂过度表达来测试每个miRNA在调节E-cadherin表达和控制NMuMG细胞EMT中的功能作用。通过对cy3标记的对照前miR进行荧光激活细胞分选分析(数据未显示),在长达3天的时间内,90%以上的细胞群实现了成功转染。转染阴性对照前miR的NMuMG细胞对TGFβ处理有反应,E-钙粘蛋白表达减少90%以上(图1电子)EMT的形态变化特征(图1F类). 相反,与五个miR-200 miRNAs中的每一个相对应的前miRs的过度表达导致E-cadherin在EMT期间对转录抑制产生显著抵抗(图1电子). miR-200b、miR-200c和miR-429在维持E-cadherin表达方面具有最强大的作用,而miR-200a和miR-141的作用相对较小(图1电子). 所有五种miRNA或两个单独簇中的miRNA的转染对E-cadherin表达的影响与miR-200b、miR-200c或miR-429的单独转染相似。此外,miRNAs作为簇的表达(图1F类)或单独(数据未显示)显著减少TGFβ诱导NMuMG细胞EMT期间间充质特征的获得(图1F类). 在miR-200转染的细胞中,尽管细胞伸长和致密细胞-细胞粘附的丧失是明显的,但向纤维母细胞形态的转变是不完整的,许多细胞簇保持着鹅卵石样的上皮特征(图1F类). miR-200转染细胞的免疫荧光分析显示E-cadherin表达水平显著,粘附连接维持。然而,TGFβ处理细胞中N-cadherin表达的激活不受miR-200过度表达的影响(数据未显示)。总的来说,这些结果表明miR-200家族的所有五个成员都能够通过维持E-cadherin的高水平表达,在功能上破坏NMuMG细胞中的上皮-间充质转换。
miR-200家族的过度表达逆转4TO7乳腺癌细胞系的间充质特征-接下来,我们测试了miR-200家族miRNAs逆转转移性乳腺癌细胞间充质表型的能力。4TO7是一种来源于野生型BALB/c小鼠自发乳腺肿瘤的乳腺癌细胞系(15). 4TO7细胞系具有高度侵袭性,能够从BALB/c受体动物的原位乳腺肿瘤中植入大量微转移(16). 显微镜下,4TO7细胞显示出典型的成纤维细胞形态(图2A类)与E-cadherin表达水平极低一致(图2B类)和miR-200家族miRNA表达(数据未显示)。两个miR-200簇分别或联合在4TO7细胞中过度表达3天,导致形态学发生了巨大变化,从纺锤状间充质细胞群转变为鹅卵石状上皮细胞群(图2A类). miR-200转染4TO7细胞的上皮表型在48小时后明显,72小时达到峰值,转染后约84小时恢复为间充质表型,可能是由于转染前miRs降解所致。免疫荧光分析显示E-cadherin表达显著增加。重要的是,E-cadherin在miR-200过表达细胞的质膜上富集,并在相邻细胞之间形成粘附连接,这表明E-cadherin在功能上有助于4TO7细胞上皮表型的获得。
miR-200家族miRNAs直接靶向E-cadherin转录抑制子ZEB1和ZEB2-为了确定miR-200 miRNA的可能直接靶点,我们搜索了TargetScan数据库(17)已知E-cadherin表达抑制因子mRNA序列中miR-200靶点,包括蜗牛、鼻涕虫、扭体蜗牛、Goosecoid蜗牛、FoxC2蜗牛、ZEB1蜗牛、和ZEB2型.E-box-binding锌指转录因子ZEB2型(SIP1/ZFXH1B型)被预测为miR-200家族最可能的靶基因,因为其3′-UTR至少包含两个miR-200a/141位点和五个miR200b/200c/429位点(图2C类).泽布1(TCF8型/δEF1/无-2α) ,与属于同一家庭ZEB2型,也预计包含至少两个miR-200b/200c/429的靶位点。测试的直接目标ZEB1号机组和ZEB2型通过miR-200,我们克隆了萤火虫荧光素酶报告基因下游的3′-UTR序列。报告质粒与miR-200a、miR-200b、miR-429以及4TO7细胞中的每个或两个簇的共转染导致ZEB2-3′-UTR-荧光素酶表达显著降低,表明这些miRNAs可能靶向ZEB2型直接(图2天,左侧面板). 在报告者分析中使用ZEB1号机组3′-UTR,只有miR-200b和两个簇能够显著降低荧光素酶报告基因的表达,这可能反映了miR-200位点在ZEB1号机组将3′-UTR片段克隆到荧光素酶报告质粒中。从HeLa细胞系中获得了类似的报告基因测定结果(图2天,右侧面板). 报告者分析结果在NMuMG细胞中不太一致(数据未显示),可能是由于该上皮细胞系中miR-200的高基础水平表达。
测试内源性药物的靶向性ZEB1号机组和ZEB2型我们分析了miR-200的转录本ZEB1号机组和ZEB2型单个、同时或成簇转染miR-200家族成员的NMuMG细胞中的mRNA水平。然后诱导转染细胞进行EMT和TGFβ1处理。尽管ZEB1号机组和ZEB2型在对照细胞EMT期间,miR-200家族每个成员的过度表达都显著减少了两者的积累ZEB1号机组和ZEB2型抄本。因此,miR-200家族成员可能通过直接降低E-cadherin转录抑制因子的表达来增强E-cadherin的表达。
测试是否直接瞄准ZEB1号机组和ZEB2型也参与4TO7乳腺肿瘤细胞中E-钙粘蛋白的上调,我们在转染miR-200簇后48小时进行了qRT-PCR分析。E-钙粘蛋白表达的急剧增加伴随着ZEB1号机组和ZEB2型mRNA水平(图2F类). 时间进程分析显示ZEB1号机组和ZEB2型在48小时后维持的4TO7细胞中在24小时内。然而,ZEB1号机组和ZEB2型转染72h后靶向性消失,与转染后~84h间充质表型的逆转一致。
miR-200家族miRNAs抑制4TO7乳腺癌细胞迁移-E-cadherin的丢失和间充质特性的获得,包括运动能力的增强,被认为是良性肿瘤向恶性肿瘤转变过程中的关键事件。鉴于转染miR-200家族miRNAs的4TO7乳腺癌细胞中E-cadherin的强烈抑制和上皮特性的显示,我们试图确定这些miRNAss在细胞迁移中的作用。使用在体外跨阱迁移分析表明,miR-200簇的过度表达强烈降低了生长因子诱导的4TO7细胞定向迁移(图2G公司).
讨论
在胚胎发育过程中,EMT是各种组织和器官(如神经嵴、心脏、肌肉骨骼系统和周围神经系统)形成的关键过程。然而,在成人生活中,只有特定的细胞亚群保留了接受短暂EMT的能力,例如在伤口愈合过程中的角质形成细胞(18). 已知恶性乳腺肿瘤细胞会重新激活胚胎程序,如EMT,以获得选择性优势,如增强运动性和侵袭性(16,19,20). 由于EMT与浸润性癌密切相关,因此这一过程的分子调控可能是治疗干预的关键。
先前的研究将miR-200家族与上皮表型和ZEB家族联系起来。发现miR-200家族在上皮组织中的表达丰富(21,22)与胚胎发育期间ZEB1和ZEB2的表达呈负相关(23). 此外,ZEB家族与EMT、肿瘤发生和转移有关(24–27). 然而,在这些先前的研究中,尚未研究miR-200在EMT和肿瘤迁移中的功能性参与,以及整个miR-200家族对ZEB1和ZEB2的直接靶向性。
在我们目前的研究中,我们通过直接靶向EMT,确定miR-200家族是EMT的抑制物ZEB1号机组和ZEB2型miR-200家族由五个成员组成,排列成两个簇,miR-200a/200b/429和miR-200c/141。ZEB1号机组和ZEB2型可能是靶点,因为两者都是EMT诱导剂,并且包含miR-200家族许多成员的靶点。ZEB2型所有五种miRNAs都有靶位点,但TargetScan数据库表明ZEB1号机组miR-200b、-200c和-429只有两个靶点。相反,我们的结果表明,miR-200a和-141的过度表达也显著降低了内源性ZEB1号机组(图2天),使我们相信ZEB1号机组事实上,可能存在这些miRNAs的其他靶点。事实上,最近的一份报告发现Genbank™Refseq条目ZEB1号机组(NM_030751号)被人为截断,人工检测发现miR-200a和miR-141在泽布13′-UTR(28).
我们发现miR-200家族的异位表达单独、成簇或全部阻碍了TGFβ处理的NMuMG细胞的EMT进展ZEB1号机组和ZEB2型表达水平低,E-cadherin表达水平高(图。1电子和2天). 此外,这些过度表达miR-200家族的细胞保持着鹅卵石样上皮表型。尽管miR-200a和miR-141一直是E-cadherin最弱的阳性调节因子,但每个miR-200 miRNA的异位表达足以阻碍EMT进展。来自同一簇或两个簇的miRNAs同时过表达似乎在调节EMT进展方面没有协同作用ZEB1、ZEB2或E-cadherin,至少在在体外设置。虽然我们发现miR-200家族可以阻碍EMT,但它们的过度表达不足以完全阻断这一过程,这表明其他miRNAs也可能参与调节EMT。
miR-200家族在4TO7癌细胞中的异位表达导致ZEB1号机组和ZEB2型水平。在4TO7细胞中过度表达miR-200家族导致了从纺锤状间充质表型到更显著的上皮表型的显著形态学变化,并形成了粘附连接。此外,miR-200的过度表达显著抑制生长因子诱导的定向迁移,这是转移癌细胞的一个特征。这表明miR-200家族的异位表达可以促进间质-上皮转化,减少肿瘤细胞迁移。由于ZEB1和ZEB2与各种肿瘤类型的进展有关,miR-200家族可能为转移癌的靶向治疗提供重要途径。
致谢
我们感谢康实验室的成员提出的有益建议和讨论。我们感谢希拉里·科勒博士和阿斯特·莱格斯·米勒博士提供的技术专长和有益的讨论。
*这项工作得到了美国陆军医学研究与材料司令部(Grant W81XWH-06-1-0481)的支持,并得到了美国癌症协会(Grant RSG MGO-110765)和Susan G.Komen基金会(Grant BCTR0503765)的额外支持。这篇文章的出版费用部分由页面费支付。因此,必须在此标记“广告“根据《美国法典》第18卷第1734节,仅为了表明这一事实。
脚注
4使用的缩写是:nt,nucleotide;上皮-间充质转化;转化生长因子;miRNA,microRNA;前miR,前microRNA;UTR,非翻译区;qRT-PCR,定量逆转录-PCR。
工具书类
-
1Lee,Y.、Ahn,C.、Han,J.、Choi,H.、Kim,J.和Yim[内政部][公共医学][谷歌学者]
-
2Yi,R.、Qin,Y.、Macara,I.G.和Cullen,B.R.(2003)基因开发17 3011–3016[内政部][PMC免费文章][公共医学][谷歌学者]
-
三。Hutvagner,G.、McLachlan,J.、Pasquinelli,A.E.、Balint,E.、Tuschl,T.和Zamore,P.D.(2001)《科学》293 834–838[内政部][公共医学][谷歌学者]
-
4Wienhold,E.、Kloosterman,W.P.、Miska,E.、Alvarez-Saavedra,E.,Berezikov,E.、de Bruijn,E.、Horvitz,H.R.、Kauppinen,S.和Plasterk,R.H.(2005)《科学》309 310–311[内政部][公共医学][谷歌学者]
-
5Yi,R.、O'Carroll,D.、Pasolli,H.A.、Z.、Dietrich,F.S.、Tarakhovsky,A.和Fuchs,E.(2006)《自然遗传学》。38 356–362[内政部][公共医学][谷歌学者]
-
6Esquela-Kerscher,A.和Slack,F.J.(2006)《国家癌症评论》6 259–269[内政部][公共医学][谷歌学者]
-
7Johnson,S.M.、Grosshans,H.、Shingara,J.、Byrom,M.、Jarvis,R.、Cheng,A.、Labourier,E.、Reinert,K.L.、Brown,D.和Slack,F.J.(2005)Cell 120 635–647[内政部][公共医学][谷歌学者]
-
8Huang,Q.、Gumireddy,K.、Schrier,M.、le Sage,C.、Nagel,R.、Nair,S.、Egan,D.A.、Li,A.、Huang,G.、Klein Szanto,A.J.、Gimotty,P.A.、Katsaros,D.、Coukos,G.、Zhang,L.、Pure,E.和Agami,R.(2008)《自然细胞生物学》。10 202–210[内政部][公共医学][谷歌学者]
-
9Ma,L.、Teruya-Feldstein,J.和Weinberg,R.A.(2007)《自然》449 682–688[内政部][公共医学][谷歌学者]
-
10Tavazoie,S.F.、Alarcon,C.、Oskarsson,T.、Padua,D.、Wang,Q.、Bos,P.D.、Gerald,W.L.和Massague,J.(2008)《自然》451 147–152[内政部][PMC免费文章][公共医学][谷歌学者]
-
11Zhu,S.,Wu,H.,Wu,F.,Nie,D.,Sheng,S.和Mo,Y.Y.(2008)细胞研究18 350–359[内政部][公共医学][谷歌学者]
-
12Asangani,I.A.、Rasheed,S.A.、Nikolova,D.A.、Leupold,J.H.、Colburn,N.H.,Post,S.和Allgayer,H.(2008)癌基因27 2128–2136[内政部][公共医学][谷歌学者]
-
13Christoffersen,N.R.、Silahtaroglu,A.、Orom,U.A.、Kauppinen,S.和Lund,A.H.(2007)《RNA(冷泉港)》13 1172-1178[内政部][PMC免费文章][公共医学][谷歌学者]
-
14Hurteau,G.J.、Carlson,J.A.、Spivack,S.D.和Brock,G.J(2007)《癌症研究》67 7972–7976[内政部][公共医学][谷歌学者]
-
15Aslakson,C.J.和Miller,F.R.(1992)《癌症研究》52 1399–1405[公共医学][谷歌学者]
-
16Yang,J.、Mani,S.A.、Donaher,J.L.、Ramaswamy,S.、Itzykson,R.A.、Come,C.、Savagner,P.、Gitelman,I.、Richardson,A.和Weinberg,R.A.(2004)Cell 117 927–939[内政部][公共医学][谷歌学者]
-
17Lewis,B.P.、Burge,C.B.和Bartel,D.P.(2005)Cell 120 15–20[内政部][公共医学][谷歌学者]
-
18Thiery,J.P.(2003)当前版本。操作。细胞生物学。15 740–746[内政部][公共医学][谷歌学者]
-
19Huber,M.A.、Azoitei,N.、Baumann,B.、Grunert,S.、Sommer,A.、Pehamberger,H.、Kraut,N.,Beug,H.和Wirth,T.(2004)《临床杂志》。投资。114 569–581[内政部][PMC免费文章][公共医学][谷歌学者]
-
20Muraoka,R.S.、Dumont,N.、Ritter,C.A.、Dugger,T.C.、Brantley,D.M.、Chen,J.、Easterly,E.、Roebuck,L.R.、Ryan,S.、Gotwals,P.J.、Koteliansky,V.和Arteaga,C.L.(2002)J.Clin。投资。109 1551–1559[内政部][PMC免费文章][公共医学][谷歌学者]
-
21Baskerville,S.和Bartel,D.P.(2005)RNA(冷泉港)11 241–247[谷歌学者]
-
22Lu,J.,Getz,G.,Miska,E.A.,Alvarez-Saavedra,E.,Lamb,J..,Peck,D.,Sweet-Cordero,A.,Ebert,B.L.,Mak,R.H.,Ferrando,A.A.,Downing,J.R.,Jacks,T.,Horvitz,H.R.和Golub,T.R.(2005)《自然》435 834–838[内政部][公共医学][谷歌学者]
-
23Miyoshi,T.、Maruhashi,M.、Van De Putte,T.、Kondoh,H.、Huylebroeck,D.和Higashi,Y.(2006)开发动力学235 1941-1952[内政部][公共医学][谷歌学者]
-
24Spoelstra,N.S.、Manning,N.G.、Higashi,Y.、Darling,D.、Singh,M.、Shroyer,K.R.、Broaddus,R.R.、Horwitz,K.B.和Richer,J.K.(2006)《癌症研究》66 3893–3902[内政部][公共医学][谷歌学者]
-
25Spaderna,S.、Schmalhofer,O.、Hlubek,F.、Berx,G.、Eger,A.、Merkel,S.,Jung,A.、Kirchner,T.和Brabletz,T.(2006)《胃肠病学》131 830–840[内政部][公共医学][谷歌学者]
-
26Maeda,G.、Chiba,T.、Okazaki,M.、Satoh,T.,Taya,Y.、Aoba,T..、Kato,K.、Kawashiri,S.和Imai,K.(2005)《国际期刊》。27 1535–1541[公共医学][谷歌学者]
-
27Imamichi,Y.、Konig,A.、Gress,T.和Menke,A.(2007)癌基因26 2381–2385[内政部][公共医学][谷歌学者]
-
28Gregory,P.A.、Bert,A.G.、Paterson,E.L.、Barry,S.C.、Tsykin,A.、Farshid,G.、Vadas,M.A.、Khew-Goodall,Y.和Goodall,G.J.(2008)《自然细胞生物学》。10.1038/ncb1722[内政部][公共医学]

