摘要
有潜在炎症的个体表现出包括睡眠障碍和疲劳在内的非特异性并存症状的高患病率。然而,促炎细胞因子的细胞表达、睡眠深度的改变和日间疲劳之间的关系尚未同时进行研究。在健康成年人(24-61岁)中,在评估第二天的睡眠情况和疲劳程度之前,评估夜间单核细胞内促炎细胞因子的产生水平。刺激单核细胞产生白细胞介素-6(IL-6)而非肿瘤坏死因子α(TNF-α)与慢波睡眠负相关(ΔR2=17,p=.029)。相反,在第一个睡眠周期(ΔR),刺激的单核细胞产生IL-6与快速眼动(REM)睡眠时间呈正相关2=26,p<.01)。此外,傍晚刺激产生的IL-6与第二天的疲劳有关(ΔR2=17,p=0.05)。中介分析表明,慢波睡眠,而不是快速眼动睡眠时间,介导了夜间IL-6产生水平与白天疲劳之间的关系。这些结果表明,IL6刺激的单核细胞生成增加可能与睡眠结构的改变有关,慢波睡眠减少,REM睡眠持续时间增加。慢波睡眠的相对丧失可能是细胞炎症导致白天疲劳的一种途径。
关键词:促炎细胞因子、白细胞介素-6、细胞炎症、疲劳、慢波睡眠、快速眼球运动睡眠、睡眠结构
方法
参与者
参与者包括31名男性(N=16)和女性(N=15),他们通过推荐和广告招募,寻求健康的成年人参与2006年10月至2008年6月之间的睡眠研究。所有人都同意参加这项加州大学洛杉矶分校(UCLA)机构审查委员会批准的研究。参与者年龄在24岁至61岁之间(平均值=37.16,SEM=1.59),体重指数(以千克为单位的体重除以以米为单位的身高平方)小于30,不吸烟者,如2周睡眠日记所证实的那样,他们在晚上10:30至早上7:30之间有规律地睡觉。根据病史、体检和实验室测试,所有参与者的身体状况都很健康。根据诊断和统计手册第四版(SCID)结构化临床访谈确定,他们都没有精神疾病史,也没有参与者有睡眠障碍。经随机尿液物质筛查证实,没有一名参与者符合主要物质依赖性标准,也没有人在过去2周内使用过包括酒精在内的物质。此外,在睡眠方案的2周内,没有参与者服用已知会改变睡眠活动的药物(例如β受体阻滞剂、精神药物)。
程序
参与者在国立卫生研究院普通临床研究中心连续呆了3天(24小时),其中包括一个初始筛查/适应夜和两个晚上的睡眠测试;只有第一晚的睡眠测试结果才包括在内,因为那晚是疲劳评估的前夜。在适应睡眠实验室对睡眠呼吸暂停和夜间肌阵挛进行筛查后,参与者接受了多导睡眠描记仪测试,环境光线暗度超过50勒克斯。参与者遵守正常的睡眠时间表,晚上11点至早上7点熄灯。在夜间,如果参与者需要在夜间小便,则使用床边小便器。参与者在早上醒来时,打开昏暗的灯光并呼唤参与者的名字。所有参与者在与天之间一直在GCRC上,直到完成睡眠方案。
睡眠监测通过放置脑电图(C3或C4)电极、眼电图和颏下肌电图记录进行。根据Rechtschaffen和Kales的标准对EEG睡眠记录进行视觉评分(Rechtschaffen和Kales,1968年)如前所述(Irwin等人,2002年). 为了本研究的目的,计算的睡眠变量包括总睡眠时间(TST)、睡眠效率(TST/床上时间的百分比)、睡眠开始时间、第一阶段睡眠、第二阶段睡眠、SWS(包括第三阶段和第四阶段睡眠)、REM潜伏期、REM密度和第一个睡眠周期中的REM持续时间。TST被认为是从睡眠开始到最后醒来的总分钟数,包括睡眠和REM睡眠阶段1到4所花费的时间。适应/筛查之夜与测试之夜在睡眠连续性和睡眠结构的多导睡眠图测量方面没有差异。
在睡眠测试前的晚上11点(晚上)和睡眠后的早上8点(早上)分别进行血液采样,并通过静脉留置的前臂采集样本。分析单核细胞群中细胞内促炎细胞因子的表达。
疲劳是在睡眠测试后的晚上使用简式36健康调查问卷(SF-36(Ware and Sherbourne,1992年). 构成活力分量表的项目评估受访者是否充满活力、疲倦、精疲力竭或精力充沛。该量表的高分代表活力更强,而低分代表疲劳。
细胞因子的单核细胞表达
通过流式细胞术评估单核细胞内IL-6和TNF-α的表达(参见Collado-Hidalgo等人,2006年有关这些程序的完整描述)。将肝素处理过的血液与100pg/mL脂多糖(LPS;Sigma,St-Louis,Mo)和10μg/mL布雷费尔丁A(Sigma)混合,并在37°C的平台混合器中培养4小时,然后在4°C下培养过夜。我们之前在剂量-反应谱研究中发现,100 pg/ml LPS持续4小时产生阈值反应,同时检测到IL-6和TNF-α的刺激产生。这高于未刺激对照样品中的水平,但通常低于次最大反应,IL-6和TNF-α的联合水平介于50%至75%之间(Irwin等人,2010年;Irwin等人,2006b). 在许多人中,50 pg/ml的LPS剂量不能产生阈值反应,而200 pg/ml剂量的LPS产生的反应聚集在80%至90%的最大范围内。细胞在荧光活化细胞分选渗透缓冲液(加利福尼亚州圣何塞BD Biosciences)中渗透后,添加荧光结合抗体,约12000 CD14+对事件进行计数,以确定分泌细胞因子的单核细胞的净刺激百分比,并基于未刺激细胞设置象限坐标。为了确定表达TNF-α或IL-6的受刺激细胞的百分比,从受刺激百分比中减去未受刺激的细胞因子阳性事件百分比,以获得净受刺激细胞因子阳性的事件百分比。
统计分析
单核细胞因子表达与多导睡眠
数据采用SPSS 16.0软件进行分析。在进行分析之前,SWS和REM睡眠测量值进行了对数转换,以控制偏度。进行相关性分析,作为晚上和早上刺激的促炎细胞因子表达和睡眠变量之间的初步检查。基于这些结果,在控制了年龄和BMI后,进行了分层线性回归分析,以检查夜间IL-6和TNF-α的细胞表达是否与夜间SWS和REM睡眠测量值相关。在这些分析中,SWS和快速眼动睡眠测量值被输入为结果变量。根据之前的研究发现,年龄、BMI、性别和种族与炎症和睡眠有关(Hong等人,2005年;Lim等人,2005年;O'Connor等人,2009年;O'Connor等人,2007年),这些变量作为协变量输入到模型的块1中。IL-6和TNF-α的细胞表达被作为预测变量输入模型Block 2。
次级介导分析:细胞因子表达、睡眠和疲劳
对于能够显著预测REM睡眠和SWS的炎症标记物,我们进行了中介分析,以测试在单核细胞细胞因子产生量较大的个体中观察到的REM睡眠改变和SWS是否会导致第二天的疲劳。这些分析是使用Baron和Kenny概述的方法在一系列层次线性回归分析中进行的(拜伦和肯尼,1986年),REM睡眠测量和SWS作为单核细胞细胞因子产生和疲劳之间关系的介质进入。为了建立调解,我们随后测试了是否满足以下四个条件。首先,中介(SWS,REM睡眠测量)必须与结果变量(疲劳)相关。其次,预测变量(刺激的IL-6,TNF-α)必须与介质(SWS,REM睡眠测量)相关。第三,预测变量必须与结果变量(疲劳)相关。最后,当结果变量同时回归到中介变量和预测变量上时,中介变量和结果变量之间的相关性应该比预测变量和结果参数之间的相关性更强。我们使用一系列层次回归分析分三步测试了这些条件。年龄、BMI、性别和种族作为协变量输入第一组分析。
结果
样品特征
样本特征列于表1样本的种族细分为34.3%的白人美国人、31.4%的非裔美国人、14.3%的亚裔美国人和11.4%的西班牙裔。大约一半的样本年收入为30000美元或更少,平均受教育年限为15.37年(SEM=.30)。免疫和睡眠数据包括在表2.
表1。
| 变量 |
M±SEM(扫描电镜) |
| 年龄 |
37.16± 1.59 |
| 体重指数 |
24.60 ± .74 |
| 受教育年限 |
15.55 ± .31 |
| SF-36活力/疲劳 |
72.41 ± 2.423 |
| 种族 |
N(%) |
| 非裔美国人 |
10 (32.3) |
| 亚裔美国人 |
3 (9.7) |
| 白人美国人 |
11 (35.5) |
| 西班牙裔 |
2 (6.5) |
| 收入 |
N(%) |
| <$30,0000 |
15 (48.4) |
| $30,000–49,999 |
8 (25.8) |
| ≥$50,000 |
7 (22.6) |
表2。
| 变量 |
平均值±SEM |
| 第1晚 |
第二晚 |
| 晚间IL-6产量(%) |
10.21 ± 1.17 |
7.96 ± 1.14 |
| 夜间TNF-a产量(%) |
23.99 ± 2.39 |
20.28 ± 1.62 |
| 总睡眠时间(TST;分钟) |
391.12. ± 11.07 |
414.05 ± 14.46 |
| 睡眠开始时间(分钟) |
32.13 ± 7.45 |
24.32 ± 5.14 |
| 睡眠效率(TST/床上时间×100;%) |
80.89 ± 2.22 |
82.94 ± 2.30 |
| 第1阶段(%TST) |
4.34 ± 0.43 |
5.46 ± 0.56 |
| 第2阶段(%TST) |
65.04 ± 1.51 |
62.22 ± 2.06 |
| SWS(TST百分比) |
7.05 ± 1.21 |
9.29 ± 1.43 |
| REM持续时间(分钟) |
17.48 ± 2.29 |
16.22 ± 1.91 |
| REM密度(分钟) |
1.37 ± 0.14 |
1.59 ± 0.18 |
| REM延迟(分钟) |
64.25 ± 5.68 |
62.16 ± 4.61 |
| REM(%TST) |
22.63 ± 1.43 |
23.03 ± 1.29 |
双变量相关性研究了刺激单核细胞产生IL-6和TNF-α、睡眠变量和疲劳之间的关系。与我们的假设一致,夜间刺激的单核细胞产生IL-6与较少的SWS(r=−.517,p=.01)和较长的REM持续时间(r=.553,p=.008)相关。IL-6的产生与第1或第2阶段睡眠、TST、睡眠效率、睡眠开始时间、REM睡眠、REM潜伏期或REM密度无关。夜间TNF-α的产生与睡眠阶段、总睡眠时间、睡眠效率或睡眠开始时间无关。早上产生的IL-6和TNF-α均与睡眠变量无关。
单核细胞因子的产生与睡眠结构
考虑到IL-6单核细胞夜间表达水平与SWS和REM持续时间测量值之间的双变量关联,利用该预测变量进行了分层线性回归分析。年龄、BMI、性别和种族解释了8.3%的SWS变异。在控制这些协变量后,夜间刺激的单核细胞产生IL-6越多,SWS(ΔR2=.17,p=.029)(参见图1). 如图所示图2,IL-6产生最低四分位的参与者在SWS中花费了11.3%的TST,而在IL-6产生最高四分位者中花费了2.5%的TST。
图1。
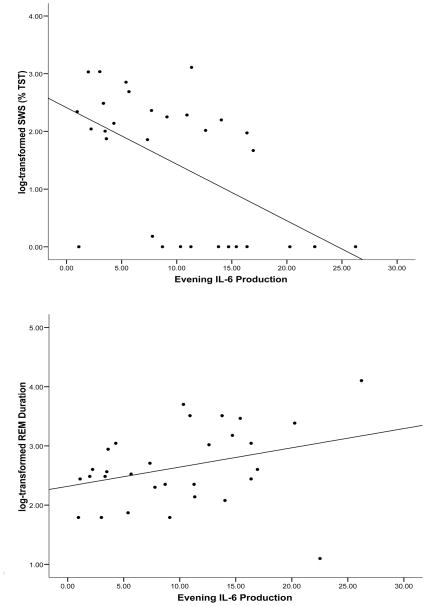
夜间刺激的单核细胞产生IL-6、对数转换SWS和对数转换REM持续时间之间的关系。IL-6分泌增加与SWS缩短和REM持续时间延长相关。
图2。
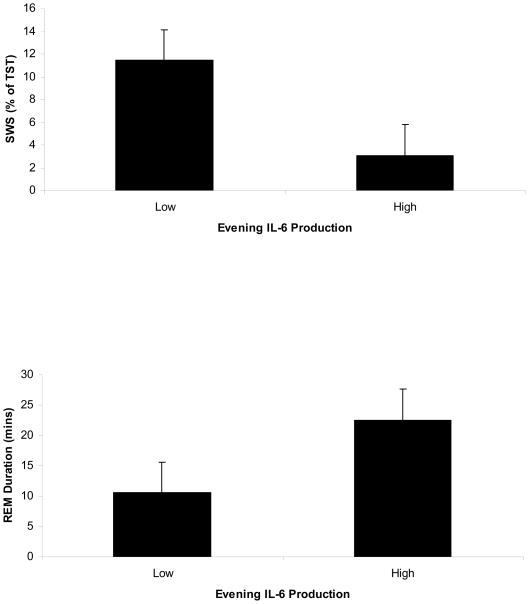
图中描绘了最高和最低四分位晚间刺激单核细胞产生IL-6的参与者的SWS和REM持续时间。
这些协变量解释了REM持续时间中12.5%的变化。如图所示图1夜间刺激的单核细胞分泌IL-6越多,REM持续时间越长(ΔR2=.18,p=.017)。图2说明了IL-6产生最低四分位参与者的REM持续时间为10.6分钟,而IL-6产生最高四分位的参与者为22.5分钟。TNF-α与SWS或REM持续时间无关。
中介分析:夜间细胞因子表达、睡眠结构和疲劳
进行中介分析以验证我们的假设,即SWS和REM持续时间会介导夜间促炎细胞因子表达和疲劳之间的关系。年龄、BMI、性别和种族占疲劳方差的10%。在控制这些协变量后,较少的SWS与第二天的疲劳相关(ΔR2=15,p=0.03;调解模式的条件1)。我们的发现满足了调解模型的条件2,即夜间刺激的单核细胞产生IL-6越多,SWS越少(见上一节)。夜间单核细胞产生IL-6与疲劳相关(ΔR2=17,p=0.05;条件3)。表3列出了中介模型条件4的最终层次线性回归分析测试。如图所示表3当同时检测IL-6和SWS的产生作为疲劳的预测因子时,只有SWS与第二天的疲劳显著相关(β=.55,p=.02)。换句话说,夜间IL-6分泌增加与疲劳有关,这种关系由较短的SWS介导(参见图3). 因此,调解模型的条件4得到了满足,并且支持我们的假设,即促炎细胞因子表达和疲劳之间的关联将由改变的SWS介导。
表3。
最终层次回归分析表明SWS介导夜间IL-6产生与疲劳之间的关系(N=31)
| 变量 |
B类 |
SEB公司 |
β |
R(右)2
|
ΔR2
|
| 区块1 |
|
|
|
0.10 |
0.10 |
| 体重指数 |
−0.32 |
0.57 |
−0.10 |
|
|
| 年龄 |
0.25 |
0.24 |
0.182 |
|
|
| 性别 |
−6.48 |
4.67 |
−0.25 |
|
|
| 种族 |
0.24 |
1.39 |
0.03 |
|
|
| 区块2 |
|
|
|
0.32 |
0.22* |
| IL-6产生 |
−.59 |
0.45 |
−0.22 |
|
|
| 开关量
|
6.38 |
2.44 |
0.55 |
|
|
图3。
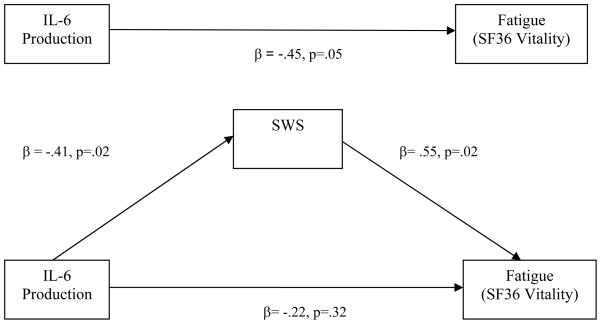
通径分析表明,对数转换SWS介导夜间刺激的单核细胞产生IL-6与疲劳之间的关系。当IL6和SWS都进入回归模型时,SWS(而非IL-6)仍然是疲劳的重要预测因子。
还进行了介导分析,以确定REM持续时间的改变是否介导了IL-6和疲劳之间的关系。在控制了年龄和BMI后,REM持续时间与疲劳无关。因此,没有证据表明REM持续时间的增加介导了IL-6的产生和疲劳之间的关系。
讨论
这项研究检查了促炎细胞因子表达、睡眠结构和疲劳之间的关系。有人支持我们的假设,即促炎细胞因子表达的增加与睡眠结构的改变有关。具体而言,夜间刺激的单核细胞产生IL-6越多,SWS越少。这支持了一项研究结果,该研究表明,循环IL-6水平高的个体SWS水平较低(Burgos等人,2006年;Hong等人,2005年),并提示免疫细胞(即单核细胞)可能是与SWS相关的IL-6循环水平变化的来源。
夜间IL-6分泌增加也与第一个睡眠周期的REM持续时间延长有关。我们之前发现,IL-6的循环和刺激产生都与快速眼动睡眠量的增加有关(Irwin等人,2004年;Motivala等人,2005年;Redwine等人,2003年). 然而,在本研究中,刺激的IL-6和REM睡眠之间的相关性最强的是REM持续时间,即第一个睡眠周期中REM睡眠的时间。考虑到正常睡眠者的第一个睡眠周期通常以SWS为主,我们的研究结果可能反映出在促炎细胞因子高表达的参与者中,从SWS向REM睡眠的转变。因此,如图2睡眠阶段的这种改变显著降低了个体在总睡眠时间内获得SWS的总百分比。
有趣的是,在目前的研究中,TNF-α与睡眠结构无关。根据研究发现,TNF-α增加与酗酒者REM睡眠时间延长有关(Irwin和Rinetti,2004年)我们预计TNF-α的产生增加会对睡眠结构产生有害影响。然而,这些因素在当前研究中是无关的。与TNF-α相反,IL-6可能在调节健康人的SWS和REM睡眠中发挥独特作用。然而,我们发现TNF拮抗剂可以部分恢复REM睡眠(Irwin等人,2009年),这种药物对体内TNF-α和IL-6水平。
心理压力可能是预测IL-6产生增加和睡眠结构改变的关键因素。有充分证据表明,慢性应激与SNS活性增加有关(Thomas等人,2004年)以及产生更多的促炎细胞因子(Kiecolt-Glaser等人,2002年;Kiecolt-Glaser等人,2003年). 此外,研究发现心理压力与SWS减少和REM睡眠增加有关(Cheeta等人,1997年;Kim和Dimsdale,2007年). 鉴于心理压力会触发社交网络活动(Grippo and Johnson,2009年)有趣的是,我们观察到了一个类似的模式,即IL-6与睡眠阶段的关系,正如SNS与睡眠阶段之间所显示的那样。具体来说,夜间SNS活动增加与SWS减少和REM睡眠增加有关(普兰特,2006年;Rasch等人,2007年). 综上所述,这些发现表明,夜间SNS活动增强,可能是由于压力所致,可能会导致促炎细胞因子的产生增加,从而改变睡眠阶段。然而,在目前的研究中,我们没有检查心理压力和SNS输出是否会导致促炎性细胞因子的产生增加。未来的研究应该在经历应激性生活事件的个体中检验促炎细胞因子、睡眠结构和活力之间的关系。
我们的发现将对促炎细胞因子和睡眠结构的研究扩展到循环细胞因子水平之外,以检查睡眠结构和细胞因子表达之间的关系。鉴于循环细胞因子水平可能来自免疫细胞以外的其他来源,可能并不一定表明免疫失调,我们的研究结果进一步证明,免疫失调导致的促炎细胞因子活性增加在调节睡眠阶段中起着重要作用。
在目前的研究中,IL-6的产生增加与疲劳有关。值得注意的是,SWS(而非REM睡眠)介导了这种关系。具体而言,促炎细胞因子活性增加的个体的SWS较短解释了疲劳的报告。我们以前的研究表明,结扎Toll-4受体后测得的单核细胞IL-6的生成在睡眠剥夺后增加(Irwin等人,2010年;Irwin等人,2006b)与乳腺癌幸存者的疲劳有着独特的关系(Collado-Hidalgo等人,2006年;Collado-Hidalgo等人,2008年a). 此外,导致IL-6过度表达的细胞因子基因多态性(Collado-Hidalgo等人,2008b)与IL-6单核细胞生成增加相关,IL-6与乳腺癌幸存者的疲劳相关(Collado-Hidalgo等人,2006年).
这些发现也与Cho及其同事一致(Cho等人,2009年)在参与基于CARDIA人群的研究的健康个体中,循环炎症标记物(即C反应蛋白)与更大的疲劳有关(Cho等人,2009年). 此外,目前的研究结果表明,SWS减少可能是炎症导致健康人总体健康和疲劳降低的一种途径。有趣的是,在目前的研究以及Cho及其同事进行的研究中(Cho等人,2009年),参与者一般年轻,没有重大疾病。鉴于在这两项研究中炎症都与疲劳有关,这些发现强调了免疫失调对一般健康和其他健康个体健康的有害影响。目前尚不清楚在患有疾病的个体中,促炎细胞因子、睡眠结构和疲劳之间的关系是否相同。未来的研究应该检查这些发现对这些人的影响程度。
在解释这些发现时,应考虑当前研究中的几个局限性。首先,这项研究是在一个小型便利的社区居住成年人样本上进行的,应该在更大的样本中进行复制。还应注意的是,睡眠呼吸暂停和肌阵挛的筛查是在适应之夜进行的。这可能会影响参与者适应睡眠实验室的能力,导致测试第一晚出现“睡眠反弹”,尽管筛选/适应夜和测试夜的睡眠参数没有差异。理想情况下,参与者的睡眠将在其家庭环境中进行评估,以获得更可靠的促炎细胞因子与睡眠参数之间的关系。然而,这在当前研究中是不可行的。
这是首次发表的研究,旨在研究刺激的IL-6与睡眠结构之间的关系。此外,鉴于本研究的相关性,我们无法推断促炎细胞因子、睡眠结构和疲劳之间存在因果关系。因此,应该注意的是,这些发现是初步的,需要在未来的研究中重复。尽管如此,目前的研究结果对免疫活动、睡眠和一般健康之间的关系具有重要意义。因为人们认为SWS具有恢复功能(Tasali等人,2008年)睡眠阶段的中断可能导致白天疲劳。
致谢
这项工作得到了T32-MH19925、HL 079955、AG 026364、CA 10014152、CA116778、RR00827、P30-AG028748、普通临床研究中心计划、塞梅尔神经科学研究所加州大学洛杉矶分校表亲中心和加州大学洛杉矶大学老年美国人独立中心炎症生物学核心基金的部分支持。
脚注
出版商免责声明:这是一份未经编辑的手稿的PDF文件,已被接受出版。作为对客户的服务,我们正在提供这份早期版本的手稿。手稿在以最终可引用的形式出版之前,将经过编辑、排版和校对结果证明。请注意,在制作过程中可能会发现可能影响内容的错误,适用于该期刊的所有法律免责声明均适用。
工具书类
-
Andreakos E.类风湿性关节炎和慢性阻塞性肺病的常见和不常见特征:未来治疗的线索。当前药物靶向免疫内分泌代谢紊乱。2004;4:85–92. doi:10.2174/1568008043339910。[内政部] [公共医学] [谷歌学者]
-
Baron RM,Kenny DA。社会心理学研究中的调节-中介变量区别:概念、战略和统计考虑。心理学心理学杂志。1986;51:1173–1182. doi:10.1037//0022-3514.51.6.1173。[内政部] [公共医学] [谷歌学者]
-
Bolge SC、Doan JF、Kannan H、Baran RW。失眠与生活质量、工作效率和活动障碍的关系。Qual Life Res.2009;18:415–422. doi:10.1007/s11136-009-9462-6。[内政部] [公共医学] [谷歌学者]
-
Burgos I、Richter L、Klein T、Fiebich B、Feige B、Lieb K、Voderholzer U、Riemann D。原发性失眠症患者夜间白细胞介素-6分泌增加:一项初步研究。大脑行为免疫。2006;20:246–253. doi:10.1016/j.bbi.2005.06.007。[内政部] [公共医学] [谷歌学者]
-
Byles JE、Mishra GD、Harris MA、Nair K。老年女性的睡眠问题:健康结果的变化。年龄老化。2003;32:154–163. doi:10.1093/ageing/32.2154。[内政部] [公共医学] [谷歌学者]
-
Cheeta S、Ruigt G、van Proosdij J、Willner P。慢性轻度应激后睡眠结构的变化。生物精神病学。1997;41:419–427. doi:10.1016/S0006-3223(96)00058-3。[内政部] [公共医学] [谷歌学者]
-
Cho HJ,Seeman TE,Bower JE,Kiefe CI,Irwin MR。年轻人冠状动脉风险发展中C反应蛋白与疲劳之间的前瞻性关联研究。生物精神病学。2009年doi:10.1016/j.biopsych.2009.06.008。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Collado-Hidalgo A、Bower JE、Ganz PA、Cole SW、Irwin MR。乳腺癌幸存者持续疲劳的炎症生物标记物。2006年临床癌症研究;12:2759–2766. doi:10.1158/1078-0432.CCR-05-2398。[内政部] [公共医学] [谷歌学者]
-
Collado-Hidalgo A、Bower JE、Ganz PA、Irwin MR、Cole SW。乳腺癌幸存者的细胞因子基因多态性和疲劳:早期发现。大脑行为免疫。2008a文件编号:10.1016/j.bbi.2008.05.009。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Collado-Hidalgo A、Bower JE、Ganz PA、Irwin MR、Cole SW。乳腺癌幸存者的细胞因子基因多态性和疲劳:早期发现。大脑行为免疫。2008年b;22:1197–1200. doi:10.1016/j.bbi.2008.05.009。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Cook DN,Pisetsky DS,Schwartz DA。人类疾病发病机制中的Toll样受体。自然免疫学。2004;5:975–979. doi:10.1038/ni1116。[内政部] [公共医学] [谷歌学者]
-
Dantzer R.细胞因子诱导的疾病行为:机制和意义。美国科学院。2001;933:222–234. doi:10.1111/j.1749-6632.2001.tb05827.x。[内政部] [公共医学] [谷歌学者]
-
Dantzer R.细胞因子、疾病行为和抑郁。北美免疫过敏临床杂志2009;29:247–264. doi:10.1016/j.iac.2009.02.002。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Grippo AJ,Johnson AK。压力、抑郁和心血管失调:神经生物学机制综述和临床前疾病模型研究的整合。强调。2009;12:1–21. doi:10.1080/10253890802046281。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Hong S、Mills PJ、Loredo JS、Adler KA、Dimsdale JE。白细胞介素-6、睡眠和人口统计学特征之间的关系。大脑行为免疫。2005;19:165–172. doi:10.1016/j.bbi.2004.07.008。[内政部] [公共医学] [谷歌学者]
-
Irwin M、Gillin JC、Dang J、Weissman J、Phillips E、Ehlers CL。睡眠剥夺作为初级酗酒者稳态睡眠调节的探针。生物精神病学。2002;51:632–641. doi:10.1016/s0006-3223(01)01304-x。[内政部] [公共医学] [谷歌学者]
-
Irwin M,Rinetti G,Redwine L,Motivala S,Dang J,Ehlers C.非裔美国人戒酒后夜间促炎性细胞因子相关睡眠障碍。大脑行为免疫。2004;18:349–360. doi:10.1016/j.bi.2004.02.001。[内政部] [公共医学] [谷歌学者]
-
Irwin MR、Carrillo C、Olmstead R.睡眠不足会激活炎症的细胞标记物:性别差异。大脑行为免疫。2010;24:54–57. doi:10.1016/j.bbi.2009.06.001。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Irwin MR、Olmstead R、Valladares EM、Breen EC、Ehlers CL。肿瘤坏死因子拮抗使酒精依赖患者快速眼球运动睡眠正常化。生物精神病学。2009;66:191–195. doi:10.1016/j.biopych.2008.12.004。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Irwin MR、Rinetti G.睡眠障碍、夜间细胞因子和免疫:酒精依赖与非洲裔美国人之间的相互作用。酒精。2004;32:53–61. doi:10.1016/j.alcocol.2003.10002。[内政部] [公共医学] [谷歌学者]
-
Irwin MR、Valladares EM、Motivala S、Thayer JF、Ehlers CL。夜间迷走神经张力与酒精依赖患者的睡眠深度、睡眠质量和疲劳之间的关系。《精神病医学》,2006年a;68:159–166. doi:10.1097/01.psy.0000195743.60952.00。[内政部] [公共医学] [谷歌学者]
-
Irwin MR、Wang M、Campomayor CO、Collado-Hidalgo A、Cole S.睡眠剥夺和早晨炎症细胞和基因组标记物水平的激活。内科实习医师档案2006b;166:1756–1762. doi:10.1001/archinte.66.16.1756。[内政部] [公共医学] [谷歌学者]
-
Kiecolt-Glaser JK、McGuire L、Robles TF、Glaser R。心理神经免疫学:对免疫功能和健康的心理影响。咨询临床心理学。2002;70:537–547. doi:10.1037//0022-006x.70.3.537。[内政部] [公共医学] [谷歌学者]
-
Kiecolt-Glaser JK、Preacher KJ、MacCallum RC、Atkinson C、Malarkey WB、Glaser R.促炎细胞因子IL-6的慢性应激和年龄相关性增加。美国国家科学院院刊,2003年;100:9090–9095. doi:10.1073/pnas.1531903100。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Kim EJ,Dimsdale JE。心理社会应激对睡眠的影响:多导睡眠图证据综述。Behav睡眠医学2007;5:256–278. doi:10.1080/15402000701557383。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
LeBlanc M、Beaulieu-Bonneau S、Merette C、Savard J、Ivers H、Morin CM。基于人群的样本中与失眠相关的心理和健康相关生活质量因素。2007年精神病学研究杂志;63:157–166. doi:10.1016/j.jpsychores.2007.03.004。[内政部] [公共医学] [谷歌学者]
-
Lee M、Choh AC、Demerath EW、Knutson KL、Duren DL、Sherwood RJ、Sun SS、Chumlea WM、Towne B、Siervogel RM、Czerwinski SA。睡眠障碍与成人健康相关生活质量的关系:Fels纵向研究。《坚果健康老化杂志》。2009;13:576–583. doi:10.1007/s12603-009-010-1。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Lim W,Hong S,Nelesen R,Dimsdale JE。肥胖、细胞因子水平和抑郁症状与健康受试者疲劳程度的关系。2005年《内科实习医师档案》;165:910–915. doi:10.1001/archinte.65.8.910。[内政部] [公共医学] [谷歌学者]
-
Lorton D、Lubahn CL、Zautra AJ、Bellinger DL。风湿性疾病中的促炎细胞因子和疾病行为。当前药物设计。2008;14:1242–1260. doi:10.2174/138161208799316375。[内政部] [公共医学] [谷歌学者]
-
Motivala SJ、Sarfatti A、Olmos L、Irwin MR。抑郁症患者的炎症标记物和睡眠障碍。《精神病医学》,2005年;67:187–194. doi:10.1097/01.psy.0000149259.72488.09。[内政部] [公共医学] [谷歌学者]
-
迈尔斯JS。促炎细胞因子与疾病行为:抑郁症和癌症相关症状的影响。Oncol Nurs论坛。2008;35:802–807. doi:10.1188/08.ONF.802-807。[内政部] [公共医学] [谷歌学者]
-
O'Connor MF、Bower JE、Cho HJ、Creswell JD、Dimitrov S、Hamby ME、Hoyt MA、Martin JL、Robles TF、Sloan EK等。评估、控制和排除:生物行为因素对循环炎症标记物的影响。大脑行为免疫。2009;23:887–897. doi:10.1016/j.bi.2009.04.005。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
O'Connor MF、Motivala SJ、Valladares EM、Olmstead R、Irwin MR。IL-6单核细胞表达的性别差异:自主机制的作用。美国生理学杂志Regul Integr Comp Physiol。2007;293:R145–151。doi:10.1152/ajpregu.00752.2006。[内政部] [公共医学] [谷歌学者]
-
Ohayon MM。非恢复性睡眠投诉的患病率和相关因素。2005年《内科实习医师档案》;165:35–41. doi:10.1001/archinte.65.1.35。[内政部] [公共医学] [谷歌学者]
-
Ohayon MM。美国普通人群中的夜间觉醒和共病障碍。2008年精神病学研究杂志;43:48–54. doi:10.1016/j.jpsychires.2008.02.001。[内政部] [公共医学] [谷歌学者]
-
Plante GE。睡眠和血管疾病。新陈代谢。2006;55:S45-49。doi:10.1016/j.metabol.2006.07.013。[内政部] [公共医学] [谷歌学者]
-
Raison CL、Lin JM、Reeves WC。以人群为基础的样本中外周炎症标记物与慢性疲劳的相关性。大脑行为免疫。2009;23:327–337. doi:10.1016/j.bbi.2008.11.005。[内政部] [公共医学] [谷歌学者]
-
Rasch B,Dodt C,Molle M,Born J.血浆儿茶酚胺浓度的睡眠-年龄特异性调节。心理神经内分泌学。2007;32:884–891. doi:10.1016/j.psyneuen.2007.06.007。[内政部] [公共医学] [谷歌学者]
-
Rechtschaffen A,Kales A.人类受试者睡眠阶段标准化术语、技术和评分系统手册。贝塞斯达:1968年。[内政部] [公共医学] [谷歌学者]
-
Redwine L,Dang J,Hall M,Irwin M。酗酒者睡眠障碍、夜间细胞因子和免疫。《精神病医学》,2003年;65:75–85. doi:10.1097/01.psy.0000038943.33335.d2。[内政部] [公共医学] [谷歌学者]
-
Satoh M、Shimoda Y、Maesawa C、Akatsu T、Ishikawa Y、Minami Y、Hiramori K、Nakamura M。单核细胞中活化的toll样受体4与急性心肌梗死后心力衰竭相关。国际心脏病杂志。2006;109:226–234. doi:10.1016/j.ijcard.2005.06.023。[内政部] [公共医学] [谷歌学者]
-
Spiegel K.睡眠不足是肥胖和糖尿病的危险因素。国际儿科肥胖杂志。2008;3(补充2):27–28。doi:10.1080/17477160802404681。[内政部] [公共医学] [谷歌学者]
-
Stein MB、Belik SL、Jacobi F、Sareen J.社区睡眠问题相关损害:与身心健康共病的关系。《精神病医学》2008;70:913–919. doi:10.1097/PSY.0b013e3181871405。[内政部] [公共医学] [谷歌学者]
-
Tasali E、Leproult R、Ehrmann DA、Van Cauter E.人类慢波睡眠与2型糖尿病风险。美国国家科学院院刊2008;105:1044–1049. doi:10.1073/pnas.0706446105。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Thomas KS、Nelsen RA、Ziegler MG、Bardwell WA、Dimsdale JE。工作压力、种族和交感神经系统活动。高血压。2004;44:891–896. doi:10.1161/01.HHY.0000148499.54730.0d。[内政部] [公共医学] [谷歌学者]
-
Van Cauter E、Spiegel K、Tasali E、Leproult R。睡眠和睡眠不足的代谢后果。《睡眠医学》2008;9(补充1):S23–28。doi:10.1016/S1389-9457(08)70013-3。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Vgontzas AN、Zoumakis M、Bixler EO、Lin HM、Prolo P、Vela-Bueno A、Kales A、Chrousos GP。健康老年人与年轻人夜间睡眠受损与血浆白细胞介素-6和皮质醇水平升高相关:生理学和治疗学意义。临床内分泌代谢杂志。2003;88:2087–2095. doi:10.1210/jc.2002-021176。[内政部] [公共医学] [谷歌学者]
-
Ware JE,Jr,Sherbourne CD。MOS 36项短期健康调查(SF-36)。一、概念框架和项目选择。医疗保健。1992;30:473–483.[公共医学] [谷歌学者]
-
Winkelman JW、Redline S、Baldwin CM、Resnick HE、Newman AB、Gottlieb DJ。睡眠心脏健康研究中不宁腿综合征的多导睡眠图和健康相关生活质量相关性。睡觉。2009;32:772–778. doi:10.1093/sleep/32.6.772。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Wood LJ、Nail LM、Gilster A、Winters KA、Elsa CR。癌症化疗相关症状:提示促炎细胞因子作用的证据。Oncol Nurs论坛。2006;33:535–542. doi:10.1188/06.ONF.535-542。[内政部] [公共医学] [谷歌学者]

