摘要 Hutchinson-Gilford早衰综合征(HGPS)是一种早产综合征,由编码层粘连蛋白a的基因的显性突变引起,导致异常剪接和加工的蛋白质称为早衰蛋白。 先前的研究表明,前体蛋白诱导与DNA损伤信号增加相关的早期衰老,端粒酶延长HGPS细胞寿命。 我们证明,端粒酶通过减少前体蛋白诱导的DNA损伤信号和激活p53和Rb通路延长HGPS细胞寿命,否则会介导过早衰老的发生。 我们进一步表明,前体蛋白诱导的DNA损伤信号定位于端粒,并与端粒聚集和染色体畸变相关。 端粒酶对DNA损伤信号的改善相对较快,需要其催化和DNA结合功能,并与HGPS细胞获得增殖能力及时相关。 所有这些发现都证明HGPS细胞过早衰老是由前体激素诱导的端粒功能障碍所致。
关键词: 细胞衰老、DNA损伤、核层粘连蛋白、端粒酶、端粒功能障碍、肿瘤抑制剂
结果 TERT通过抑制肿瘤抑制通路激活拯救HGPS过早衰老 先前的研究提供了证据表明,异位端粒酶表达可以延长HGPS成纤维细胞的寿命( Kudlow等人,2008年 ; Ouellette等人,2000年 ; Wallis等人,2004年 ). 为了证实和扩大这些发现,我们用表达TERT的逆转录病毒构建物感染了增殖寿命接近尾声的HGPS成纤维细胞[剩余两个群体倍增(PD)]。 转染对照载体的HGPS成纤维细胞在两个额外的PDs内停止增殖( 图1A ). 然而,表达TERT的HGPS成纤维细胞持续增殖超过70个PDs,没有任何证据表明其增殖能力下降( 图1A ). 这种增殖能力与表达TERT的HGPS细胞S期增加和G0-G1期减少有关( 图1B ). 我们还分析了衰老相关β-半乳糖苷酶(SA-β-gal)的培养物,这是细胞衰老的经验标记( Dimri等人,1995年 ). 虽然基本上所有对照组HGPS成纤维细胞的SA-β-gal活性均为阳性,但在选择后2周,表达TERT的HGPS纤维细胞中超过90%的SA-?gal活性为阴性( 图1C ). 值得注意的是,外源性TERT表达并没有导致前体蛋白表达降低,即使在TERT存在的多次PDs后,前体蛋白水平保持不变( 图1D ).
图1。
TERT通过抑制肿瘤抑制途径的激活来挽救HGPS的早衰。 ( A类 )表达异位TERT或载体对照的HGPS成纤维细胞的生长曲线。 用逆转录病毒TERT或对照载体转导接近增殖寿命末期的HGPS成纤维细胞(剩余两个PD),并选择标记物2周,计算从选择时起的累积PD数。 ( B类 )表达异位TERT或载体对照的HGPS成纤维细胞的细胞周期分析。 逆转录病毒转导和用载体控制或TERT筛选后2周,用流式细胞术检测碘化丙锭。 误差条表示代表性实验的标准偏差,一式三份。 ( C类 )逆转录病毒转导和选择后2周,对表达异位TERT或载体对照的HGPS培养物进行SA-β-gal染色。 ( D类 )Western blot显示在TERT转导和选择之前、之后6个PDs和之后70个PDs中HGPS成纤维细胞的内源性前体蛋白水平。 层粘蛋白A、层粘蛋白C和β-肌动蛋白的蛋白质水平显示为对照。 ( E类 )Western blot显示p53、p21、p16和总Rb水平(顶部带表示Rb- P(P) )感染和选择后2周,表达TERT的HGPS成纤维细胞与载体对照组相比。 β-肌动蛋白水平显示为对照。 ( F类 )表达CDK4、DNp53、CDK4和DNp53或仅表达载体的HGPS成纤维细胞的生长曲线。 用编码所示蛋白的逆转录病毒载体转导接近增殖寿命末期的HGPS成纤维细胞(剩余两个PD),并计算从感染和选择时的累积PD。 在CDK4+DNp53的情况下,在64天时将DNp53添加到表达CDK4的细胞中。
p53和Rb抑癌途径与正常成纤维细胞的细胞衰老有关,并可被DNA损伤激活,包括导致 端粒功能障碍( 坎皮西,2001 ; Maslov和Vijg,2009年 ). 此外,据报道,p53通路在HGPS中被慢性激活( Kudlow等人,2008年 ; Liu等人,2005年 ; 刘等人,2006 ; Scaffidi和Misteli,2006年 ). 先前的研究表明,HPV E6(而非E7)抑制正常二倍体人成纤维细胞(NDFs)异位前体蛋白表达的生长抑制作用( Kudlow等人,2008年 ). 然而,这些研究并没有解决这种影响是否仅仅是由于HPV E6对p53的失活,或者是否涉及其他E6活性,例如TERT诱导。 此外,尚未研究通过直接干扰p53或Rb途径延长HGPS成纤维细胞的寿命,或TERT对这些途径激活的影响。 因此,我们分析了已知参与这些途径的几种细胞周期抑制剂的激活( 维达尔和科夫,2000年 ),并观察到用TERT转导的HGPS成纤维细胞表达的p53、p21和p16蛋白水平低于对照成纤维细胞,Rb(Rb)磷酸化(活性)形式的表达水平较高- P(P) ) ( 图1E ). 因此,TERT诱导的HGPS成纤维细胞寿命延长与p53和Rb抑癌途径激活减少相关。
为了直接研究p53和Rb通路在HGPS成纤维细胞表现出的细胞过早衰老中的作用,我们用逆转录病毒构建物感染了增殖寿命接近尾声的HGPS纤维细胞,这些逆转录病毒构造物通过CDK4过度表达阻断Rb或通过DNp53表达阻断p53。 通过测量这些成纤维细胞的增殖寿命,我们观察到CKD4能够将HGPS的寿命延长约26个PDs,而DNp53延长了HGPS约6个PDs( 图1F ). 此外,CDK4和DNp53的结合延长了HGPS细胞的寿命超过68个PDs( 图1F ). 尽管DNp53或CDK4延长寿命的相对能力在不同的实验中有所不同,但这种组合持续延长了许多PD的寿命,超过了单独使用这两种结构的寿命。 我们还观察到正常衰老成纤维细胞中DNp53和CDK4的寿命延长相似(数据未显示),表明这些相同的效应途径参与了HGPS过早衰老以及正常衰老。
TERT阻断进展诱导的DNA损伤信号 DNA损伤触发参与DNA损伤反应的蛋白质的磷酸化和/或活化。 其中包括H2AX,H2A组蛋白的一种变体,它分布在染色质中,并在新生的双链断裂处迅速磷酸化(γH2AX),以及通过自磷酸化(ATM)激活的ATM- P(P) )并被招募到双链断裂现场( Riches等人,2008年 ). 这些蛋白质在DNA损伤部位的细胞中形成离散的病灶,因此是此类损伤的有用标记。 据报道,HGPS成纤维细胞表现出增加的DNA损伤信号( Liu等人,2005年 ; 刘等人,2006 ). 当我们比较外源性TERT表达对γH2AX和ATM数量的影响时- P(P) 在这些细胞中观察到的病灶,我们发现TERT表达导致此类病灶数量显著减少( 图2A、B ). 同样,ATM的总水平- P(P) 在表达TERT的HGPS成纤维细胞中,通过免疫印迹分析可检测到明显减少( 图2C ). HGPS成纤维细胞中TERT表达的影响很快,在选择后7天γH2AX水平显著降低,这种降低与相同细胞的增殖能力增加有关( 图2G ).
图2。
TERT阻断进展诱导的DNA损伤信号。 ( A类 )γH2AX(绿色)和ATM的共焦免疫荧光显微镜- P(P) 逆转录病毒转导和标记物选择后2周,表达TERT或载体控制的HGPS成纤维细胞中的(红色)DNA损伤灶。 合并后的图像显示叠加在DNA的DAPI(蓝色)染色上,γH2AX和ATM共定位- P(P) 黄色。 ( B类 )染色定量如A所示。含有0、1、2-5或>5γH2AX和ATM的细胞百分比- P(P) 显示焦点。 每种情况下,至少有300个细胞被计数。 ( C类 )显示ATM水平的Western blot- P(P) 感染和选择后2周,HGPS成纤维细胞表达TERT或载体对照。 β-肌动蛋白水平显示为对照。 ( D类 )Western blots比较HGPS成纤维细胞中内源性前体蛋白和NDF中异位前体蛋白的水平。 层粘连蛋白A、层粘连蛋白C和β-肌动蛋白水平显示为对照。 ( E类 )γH2AX(绿色)和ATM的共焦免疫荧光显微镜- P(P) 慢病毒转导和选择后,在有或无异位TERT和表达前体蛋白或载体控制的NDF中出现(红色)DNA损伤灶。 合并后的图像叠加在DNA的DAPI(蓝色)染色上,γH2AX和ATM共定位- P(P) 黄色。 ( F类 )E中显示的染色定量。含有0、1、2-5或>5γH2AX和ATM的细胞百分比- P(P) 显示焦点。 每种情况下,至少有300个细胞被计数。 ( G公司 )异位TERT表达的HGPS成纤维细胞延长寿命和减少DNA损伤的时间进程。 用表达TERT的慢病毒或载体对照感染迟通过HGPS成纤维细胞并选择2周。 选择后立即通过追踪PD与时间来测量寿命。 使用G1门控细胞通过流式细胞术分析γH2AX来测量DNA损伤信号传导。 在第7、17、25和39天检测γH2AX阳性。 误差条表示代表性实验的标准偏差,一式三份。
为了进一步研究TERT对HGPS DNA损伤信号的影响,我们在人类NDF中体外表达了前体蛋白。 在这些细胞中,外源性前体蛋白的表达水平与HGPS成纤维细胞中内源性前体素的表达水平相似( 图2D ). 而NDF中异位前体蛋白的表达诱导了DNA损伤信号( 图2E、F ),progrein在先前感染TERT的NDF中未能做到这一点( 图2E、F ; 图3D ). 因此,TERT表达对前体蛋白诱导的DNA损伤信号具有保护作用。
图3。
TERT对前体蛋白诱导的生长缺陷和DNA损伤表型的补救作用取决于其在端粒的功能。 ( A类 , B类 )TERTs催化和DNA-结合功能是拯救HGPS早衰和DNA损伤表型所必需的。 (A) 比较表达野生型TERT、DNA-结合缺陷型TERT(N125A+T126A TERT)、催化活性TERT(D868A-TERT)或载体控制的晚期HGPS成纤维细胞的增殖能力。 在感染和选择后28天测定每个培养物的累积PD。 误差条表示三个独立实验的s.e.m。 (B) 在感染和选择后2周,对表达指定TERT结构的G1-门控HGPS成纤维细胞进行γH2AX流式细胞术分析。 误差条表示重复进行的代表性实验的范围。 ( C类 )Western印迹显示HGPS成纤维细胞中异位野生型和突变TERT构建体的蛋白水平相似。 β-actin作为负荷对照。 ( D类 )TERT对阿霉素(DOX)治疗的影响。 转导有或无异位TERT表达的NDFs(IMR90),并用前体蛋白或载体控制选择标记物。 载体对照成纤维细胞未经治疗或用500 nM DOX治疗1小时。 流式细胞术检测γH2AX阳性。 误差条表示重复进行的代表性实验的范围。
孕激素诱导的DNA损伤信号定位于端粒 接下来,我们试图确定前体蛋白表达诱导的DNA损伤信号是否定位于端粒。 为此,我们检测了表达进展蛋白的成纤维细胞是否存在端粒功能异常诱导的病灶(TIFs),如γH2AX和TRF1的共同定位所定义的,TRF1是端粒标记物( Takai等人,2003年 ). TRF2是一种端粒DNA结合蛋白,对正常端粒保护至关重要,也是TRF2(TRF2)的主要阴性形式 ΔBΔM )先前已被证明可诱导TIF的形成( Takai等人,2003年 ). 前体蛋白的异位表达诱导NDF中TIFs的形成,与TRF2观察到的方式类似 ΔBΔM ( 图4A、B ). 相比之下,检测到的γH2AX病灶对DOX诱导的DNA损伤反应没有显示端粒特异性的证据( 图4A、B ). 此外,进展诱导的TIFs偶尔涉及γH2AX与几个重叠的TRF1病灶(端粒聚集体)的共定位,这在任何分析的对照细胞中都没有观察到( 图4C ). 这些发现进一步表明端粒是前体诱导的DNA损伤信号的特定靶点。
图4。
孕激素诱导的DNA损伤信号定位于端粒。 ( A类 )在用孕激素TRF2感染NDFs后5天,通过γH2AX(绿色)和TRF1(端粒标记物)(红色)病灶的共聚焦免疫荧光显微镜检测端粒功能障碍诱导的病灶(TIFs) ΔBΔM 阳性对照或未经治疗或用500 nM DOX治疗1小时的病媒对照。 合并后的图像叠加在DAPI(蓝色)染色上,显示DNA与γH2AX和TRF1共定位为黄色。 白色方框表示放大10倍的区域,显示在右侧,没有DAPI。 比例尺:10μm。 ( B类 )通过γH2AX和TRF1的共同定位来确定A.细胞中TIFs的定量。 显示了含有0-1、2-5、6-10或>10 TIF的DNA损伤细胞的百分比。 对于每种情况,计算100个细胞。 ( C类 )进展表达细胞中TIF的其他合并图像,每个都显示端粒聚集。 检测到的TIF与A中相同。白色方框表示放大10倍的区域。 比例尺:10μm。
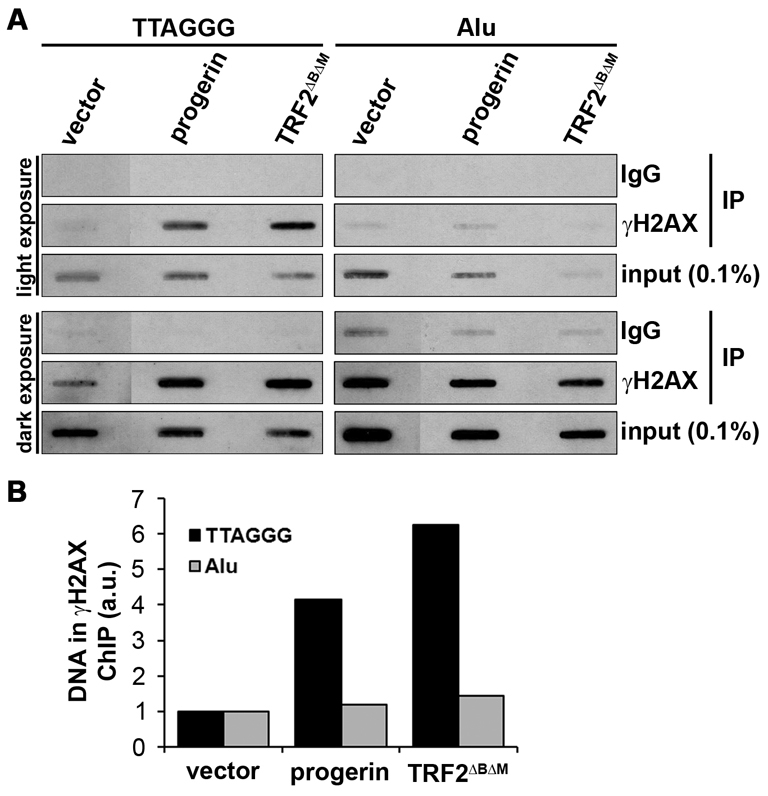
为了进一步证明在对孕激素的反应中观察到的DNA损伤信号传导定位于端粒,我们进行了端粒染色质免疫沉淀(ChIP)。 与对照组相比,progrein诱导与端粒重复序列相关的γH2AX数量增加了4.2倍,而在相同条件下,γH2AX与内部Alu序列的关联仅增加了1.2倍( 图5 ). 作为端粒特异性DNA损伤的阳性对照,TRF2 ΔBΔM 诱导与端粒重复相关的γH2AX增加6.2倍,而与Alu序列相关的γH2AX仅增加1.4倍( 图5 ). 我们对端粒DNA损伤信号的共焦成像和ChIP数据显示γH2AX增加 与端粒序列相关,提供了强有力的证据表明前体蛋白参与端粒功能障碍的诱导。
图5。
端粒染色质免疫沉淀显示,前体蛋白诱导的DNA损伤信号在端粒特异性富集。 ( A类 )感染NDF(IMR90),并用前体蛋白TRF2选择标记7天 ΔBΔM 或载体控制病毒。 免疫沉淀使用γH2AX或IgG阴性对照的特异性抗体。 使用狭缝印迹将DNA加载到尼龙膜上,并用DIG标记的端粒探针进行探测,用DIG-标记的Alu探针进行剥离和再验证。 输入DNA占总DNA的0.1%。 图中显示了明暗曝光。 ( B类 )A中所示暗暴露的量化。信号密度由ImageJ测量,直方图值表示归一化为输入信号的γH2AX端粒或Alu ChIP信号,并减去IgG对照中的背景信号。 将载体控制序列标准化为1后,每个ChIP中的DNA量以任意单位(a.u.)表示。
TRF2导致TIF形成 ΔBΔM 是TRF2从端粒DNA中分离的结果,导致3′端悬垂和DNA损伤信号的丢失( Takai等人,2003年 ). 我们使用ChIP分析比较前体蛋白和TRF2的作用 ΔBΔM TRF2与端粒DNA的关联。 如预期,TRF2 ΔBΔM 表达显著降低了TRF2与端粒的结合,而对端粒稳定性的另一个主要调节因子TRF1的结合影响最小( Smogorzewska等人,2000年 ) ( 图6A、B ). 相反,在两个独立的实验中,前体蛋白的表达与TRF1和TRF2与端粒DNA结合的增加有关( 图6A-D ). TRF1和TRF2与端粒DNA的结合是特异性的,因为在Alu序列中未检测到结合( 图6C ). 因此,与TRF2引起的端粒功能障碍不同 ΔBΔM ,进展诱导的TIF并不是由TRF2与端粒结合的显著缺失引起的。
图6。
前体蛋白对端粒结合蛋白与端粒DNA关联的影响。 ( A类 )NDF(IMR90)感染了表达前体蛋白TRF2的病毒 ΔBΔM 或选择7天的病媒控制和标记。 对于ChiP分析,在交联和DNA剪切后,使用针对TRF1、TRF2或IgG阴性对照物的抗体进行免疫沉淀。使用狭缝印迹将DNA加载到尼龙膜上,并与DIG标记的端粒探针杂交。 输入DNA占总DNA的1%。 ( B类 )A中显示的印迹定量。通过ImageJ测量信号密度,直方图值表示TRF1和TRF2端粒ChIP信号归一化为输入信号。 在载体对照序列标准化为1后,每个ChIP中端粒DNA的量以任意单位(a.u.)表达。 ( C类 )NDF(IMR90)感染表达前体蛋白或载体控制和标记的慢病毒5天。 对于ChiP分析,在交联和DNA剪切后,使用针对TRF1、TRF2或IgG阴性对照的抗体进行免疫沉淀。使用槽印迹将DNA加载到尼龙膜上,并与DIG标记的端粒探针杂交,剥离并与DIG-标记的Alu探针重杂交。 输入DNA占总DNA的1%。 ( D类 )按B所述对C中的斑点进行定量。
孕激素诱导的染色体畸变 TIFs对前体蛋白表达的反应促使我们研究是否可以检测到端粒功能障碍条件下发生的任何染色体畸变( Bolzan和Bianchi,2006年 ; 达沃利等人,2010年 ; Hande等人,2001年 ; Michishita等人,2008年 ). 我们对表达孕激素的NDFs(IMR90)的中期扩散以及载体对照进行了端粒荧光原位杂交(FISH)。 在表达进展蛋白的NDF中很难识别中期细胞,这与它们的早衰表型一致。 然而,在观察到的完整或部分前体中期中,我们发现1344条染色体中有3.5%存在异常。 这些包括染色体和姐妹染色单体融合,姐妹端粒 丢失、单个染色单体上的双端粒信号(端粒加倍)、染色体断裂、胞外端粒信号,以及两个包含双染色体的中期和一个包含核膜重组后持续存在的端粒信号的染色质桥( 图7A-H ). 相比之下,我们仅在分析的1506条控制染色体中观察到0.8%的异常。 这些结果表明,在端粒功能障碍的情况下,前体蛋白的表达促进染色体畸变。 progrein诱导过早衰老的事实可能保护这些细胞免受更严重的染色体不稳定。
图7。
孕激素诱导的染色体畸变。 来自早期传代NDF(IMR90)的中期端粒FISH外源性表达前体蛋白5、8或13天。 端粒PNA杂交信号显示为绿色,DAPI反染染色体显示为蓝色。 ( A类 )表达前体蛋白的NDF(IMR90)的代表性完全中期扩散。箭头表示端粒处两条染色体的融合,放大后显示在右下角。 ( B类 - H(H) )进展诱导染色体畸变的其他示例。 箭头表示畸变。 (B) 姐妹染色融合。 (C) 失去姐妹情侣。 (D) 端粒加倍。 (E) 染色体断裂。 (F) 染色体外端粒信号。 (G) 双染色体。 (H) 两个间期细胞核之间含有端粒信号的染色质桥。 比例尺:2μm。
材料和方法 细胞系 获得来自HGPS患者的成纤维细胞(AG01972,Coriell细胞库),并进行测序以确认G608G LMNA公司 突变。 培养物保存在补充了0.2 mM非必需氨基酸(NEAA;Invitrogen)、15%热灭活胎牛血清(FBS;Invit罗gen)和50 U/ml青霉素和链霉素(Pen/Strep;Invitorgen)的最低必需培养基(MEM;Invitro gen)中。 使用的NDF为501T(来源于成人皮肤),除非如图所示使用IMR90(胎肺,ATCC)成纤维细胞。 NDF生长在添加了10%FBS和50 U/ml Pen/Strep的Dulbecco改良Eagle's培养基(DMEM;Invitrogen)中。 所有细胞在37°C和5%CO中培养 2 细胞增殖寿命是通过以1:4或1:2的比例继代培养成纤维细胞来测量的,这取决于90%融合时的生长速度,并记录总人口倍增(PD)和培养时间。 PD通过公式PD=log计算 2 (1/分流比)( 哈雷和舍伍德,1997年 ).
表达结构、病毒产生和感染 通过RT-PCR扩增,从AG01972 HGPS细胞株的总RNA中获得全长cDNA编码前体 LMNA公司 (有义,包括GCCACC-Kozak序列,5′-GCCCACCATGGAGCCCCGTCCCAGC-3′;和反义,5′-GGCCCAGATTACATGATGCTGC-3′)。 孕激素在含有嘌呤霉素选择标记的NSPI衍生慢病毒载体中表达( Akiri等人,2009年 ). 野生型TERT、显性阴性p53(R248W)和野生型CDK4分别在含有嘌呤霉素、潮霉素和新霉素选择标记的pBabe衍生逆转录病毒载体中表达( Mahale等人,2008年 ). pBABE逆转录病毒主干中的TERT突变体N125A+T126A和D868A( Kim等人,2003年 )与野生型TERT一起亚克隆并在NSPI衍生的慢病毒载体主干中表达,该慢病毒载体具有杀鼠素选择标记。 2号机房 ΔBΔM 在pLPC逆转录病毒载体中表达,带有嘌呤霉素选择标记(Addgene质粒18008)( Karlseder等人,2002年 ). 为了建立逆转录病毒载体,HEK293T细胞与适当的逆转录病毒表达载体和pCL-ampho包装质粒共同转染。 为了建立慢病毒载体,HEK293T细胞与适当的慢病毒表达载体pCMVΔR8.74包装载体和pMD2-VSVG包膜载体共同转染。 在同一个检测细胞HT1080中进行标记选择后,通过菌落形成来确定每个病毒株的滴度,从而可以在不同的实验中使用相同数量的病毒来比较结果。 逆转录病毒和慢病毒感染是在大量纤维母细胞中使用8μg/ml聚brene(Sigma)进行的。 随后选择细胞进行抗生素耐药性检测(2μg/ml嘌呤霉素、100μg/ml潮霉素、750μg/ml新霉素和5μg/ml急变霉素),并将其扩大为群体。 在所有情况下,都使用了类似的MOI。
抗体 使用了针对以下蛋白质的抗体:Lamin A/C(Millipore,MAB3211)、p53(1801,西奈山医学院杂交瘤中心)、p21(BD Biosciences,556431)、p16(Santa Cruz,sc-468)、Rb(Cell Signaling,9309)、Ser139γH2AX(Milliporte,05-636)、Ser133γH2AX(Millipose,07-164)、Ser-1981磷酸化-ATM(Rockland,600-401-400)、, TERT(abcam,ab32020)、TRF1(#370)(纽约洛克菲勒大学Titia de Lange赠送的实物)、TRF1(Santa Cruz,sc-6165)、TRF2(Imgenex,IMG-124A)、β-肌动蛋白(Sigma,A5441)和IgG(Millipore,12-371)。
流式细胞术 根据制造商的说明,使用CycleTEST Plus DNA试剂盒(Beckton Dickinson)进行细胞周期分析。 根据制造商的说明,使用γH2AX流式细胞术磷酸化试剂盒(Millipore,17-344)检测细胞γH2AX。 为了结合细胞周期分析和γH2AX染色,将已染色的γH2AX细胞与10μg/ml碘化丙啶(Trevigen,4830-250-3)和10μg/mlRNase A(Invitrogen,12091-021)在37℃孵育10分钟。 通过FACS(FACSCalibur,Beckton Dickinson)对至少10000个染色细胞进行分类,并使用Cell Quest 3.2软件(Beckton狄金森)进行分析。
衰老相关β-半乳糖苷酶(SA-β-gal)染色 细胞在PBS中清洗,并在室温下用2%甲醛和0.2%戊二醛在PBS内固定5分钟,然后按照前面所述进行染色( Dimri等人,1995年 ).
免疫印迹法 通过在裂解缓冲液中溶解细胞获得全细胞提取物(50 mM Tris-HCl,pH 7.5,150 mM NaCl,1%Triton X-100,补充以下蛋白酶和磷酸酶抑制剂:5 mM EDTA,50 mM氟化钠,25 mMβ-甘油磷酸,1 mM原钒酸钠,0.5 mM苯甲基磺酰氟和10μg/ml抑肽酶) 或SDS裂解缓冲液[50mM Tris-HCl,pH 8.1,1%SDS,10mM EDTA,补充有完整的迷你蛋白酶抑制剂鸡尾酒(Roche,11836153001)]。 将蛋白质样品(50μg)进行SDS-PAGE,转移到Immobilon-P或Immobilon-FL过滤器(Millipore)上,并用指示的抗体进行探测。 使用ECL系统(GE Healthcare)和辣根过氧化物酶结合二级抗体(GE Hearthcare,GE Healthcare)进行检测,或使用奥德赛红外成像系统(LI-COR Biosciences,LI-COR bioscience)和IR-dye标记二级抗体进行检测。
共焦显微镜 将在玻璃盖玻片上电镀并生长至少48小时的细胞用PBS清洗两次,然后用甲醇在−20°C下固定20分钟。 然后用PBS-T(含0.1%吐温20的PBS)清洗细胞两次,在PBS-BSA-T(含1%BSA和0.1%吐温20的PBS-)中封闭1小时,并在4°C下用一级抗体孵育过夜。 然后用PBS-BSA-T清洗样品三次,并用二级抗体孵育1小时。 使用抗Abbit Cy3(Jackson ImmunoResearch,711-165-152)或抗鼠Alexa Fluor 488(Molecular Probes,A11029)二级抗体。 在一些实验中,在固定之前,添加500 nM的阿霉素(DOX)(Sigma,D1515)1小时。 使用带有DAPI(Vector Laboratories,H-1200)的Vectashield安装介质安装盖玻片。 使用蔡司LSM 510 META共焦显微镜(卡尔蔡司显微成像),使用63×油物镜进行共焦成像。 在Adobe Photoshop中对图像进行裁剪和组合。 DNA损伤量通过对含有0、1、2-5或>5γH2AX和ATM的细胞百分比进行评分来量化- P(P) 共定位病灶。 每个变量至少有300个细胞被评分。 对于TIF分析,细胞被评分为具有0-1、2-5、5-10或>10个TRF1病灶,与γH2AX染色共定位。 每个变量至少有100个细胞被评分。
端粒染色质免疫沉淀(ChIP) 根据制造商的说明,使用EZ-ChIP分析试剂盒(Millipore,17-371)进行ChIP分析。 使用Sonicator 3000(Misonix)在以下条件下进行超声检测:Amp=5.5,6个周期为20秒开启和20秒关闭。使用Ser139γH2AX(Millipore,07-164)或IgG(Millibore,12-371)抗体进行免疫沉淀,并将免疫沉淀DNA转移到Hybond-N(GE Healthcare,RPN2020N) 使用slot-blot设备的膜。 然后将膜与DIG标记的端粒(TTAGGG)杂交 4 探针并使用TeloTAGGG端粒长度检测试剂盒(Roche 12209136001)进行检测。 然后用0.2 M NaOH和0.1%SDS在52°C下洗涤两次,将膜剥离30分钟,并用DIG-标记的Alu(GGAGGCTGAGGCAGGAGAATTGCT)探针重新混合。 使用DIG寡核苷酸3′末端标记试剂盒(Roche,03353575910)进行DIG标记。 用ImageJ软件(NIH)对信号进行量化。 ChIP后端粒和Alu DNA的数量归一化为每种情况下的总输入信号。
端粒FISH在中期扩散中的作用 用0.1μg/ml去甲秋水仙素(Sigma,D7385)处理细胞16-22小时,通过胰蛋白酶消化法获取,并在37°C的0.075 M KCl中溶胀20分钟。 将细胞在−20°C的3:1甲醇:乙酸中固定过夜,滴在湿润的载玻片上,并风干过夜。 然后将细胞在PBS中再水化,并在室温下用4%甲醛在PBS内固定4分钟。 冲洗后,在70%、90%和100%乙醇系列中对细胞进行脱水,并进行空气干燥。 添加杂交溶液【70%甲酰胺,0.5%封闭试剂(Roche,11096176001),20 mM Tris-HCl,pH 7.5,和500 nM FAM-OO-(CCCTAAA)3 PNA探针(Panagene)】,在80°C下将载玻片加热3分钟,然后在室温下在黑暗中培养2小时。 将载玻片分别在洗涤液1(70%甲酰胺、10 mM Tris-HCl、pH 7.5和0.1%封闭液)中洗涤两次,每次15分钟,在洗涤液2(100 mM Tris-HCl、PH7.5、150 mM NaCl和0.08%吐温20)中洗涤三次,每次5分钟。 使用带有DAPI(Vector Laboratories,H-1200)的Vectashield安装介质安装载玻片,并使用蔡司Axioplan II显微镜(卡尔蔡司显微成像)使用63倍油物镜进行成像。 图像在Adobe Photoshop中进行裁剪和合并。
致谢 这项工作得到了NCI授予号PO1CA80058和纽约州干细胞合同#C024313的支持。 E.B.得到了NCI癌症生物学培训计划(T32 CA078207)和NIGMS细胞和分子生物学培训计划的支持(T32 GM008553)。 共聚焦激光扫描显微镜在MSSM显微镜共享资源设施进行,由NIH-NCI共享资源拨款(5R24 CA095823-04)、NSF主要研究仪器拨款(DBI-9724504)和NIH共享仪器拨款(1 S10 RR0 9145-01)资助。 我们感谢Elizabeth Blackburn(UCSF)为我们提供pBABE-puro主干中的TERT突变结构体N125A+T126A和D868A。 我们感谢Titia de Lange(洛克菲勒大学)提供的TRF1(#370)抗体和TRF2 ΔBΔM 通过Addgene获得的构造。 我们还感谢赵波和塞萨尔·穆诺兹·丰特拉的有益讨论。 存放在PMC中,12个月后发布。
工具书类
Akiri G.、Cherian M.、Vijayakumar S.、Liu G.、Bafico A.、Aaronson S.A.(2009年)。 Wnt通路异常包括Wnt自分泌激活在人非小细胞肺癌中发生率很高。 癌基因 28, 2163-2172 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Allsopp R.C.、Vaziri H.、Patterson C.、Goldstein S.、Younglai E.V.、Futcher A.B.、Greider C.W.、Harley C.B.(1992)。 端粒长度预测人成纤维细胞的复制能力。 程序。 国家。 阿卡德。 科学。 美国 89, 10114-10118 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Amiel A.、Fejgin M.D.、Goldberg Bittman L.、Sharoni R.、Hadary R.、Kitay Cohen Y.(2009年)。 丙型肝炎患者的端粒聚集。 癌症投资。 27, 650-654 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Bolzan A.D.、Bianchi M.S.(2006年)。 端粒、间质端粒重复序列和染色体畸变。 穆塔特。 物件。 612, 189-214 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Callen E.,Surralles J.(2004)。 基因组不稳定综合征中的端粒功能障碍。 穆塔特。 物件。 567, 85-104 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Campisi J.(2001)。 细胞衰老是一种肿瘤抑制机制。 趋势细胞生物学。 11,S27-S31 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Candelario J.、Sudhakar S.、Navarro S.、Reddy S.、Comai L.(2008年)。 野生型层粘连蛋白A代谢的扰动导致进展型表型。 老化细胞 7, 355-367 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Cao K.、Capell B.C.、Erdos M.R.、Djabali K.、Collins F.S.(2007年)。 Hutchinson-Gilford早衰综合征中一种层粘连蛋白A亚型过度表达,干扰早衰和正常细胞的有丝分裂。 程序。 国家。 阿卡德。 科学。 美国 104, 4949-4954 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Capell B.C.、Erdos M.R.、Madigan J.P.、Fiordalisi J.J.、Varga R.、Conneely K.N.、Gordon L.B.、Der C.J.、Cox A.D.、Collins F.S.(2005)。 抑制前体蛋白的法尼化可预防Hutchinson-Gilford早衰综合征的特征性核泡。 程序。 国家。 阿卡德。 科学。 美国 102, 12879-12884 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Choi J.、Southworth L.K.、Sarin K.Y.、Venteicher A.S.、Ma W.、Chang W.、Cheung P.、Jun S.、Artandi M.K.和Shah N.等人(2008年)。 TERT通过Myc和Wnt相关发育程序的转录控制促进上皮细胞增殖。 公共科学图书馆-遗传学。 4,e10 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Columbaro M.、Capanni C.、Mattioli E.、Novelli G.、Parnaik V.K.、Squarzoni S.、Maraldi N.M.、Lattanzi G.(2005)。 通过药物治疗挽救Hutchinson-Gilford早衰症患者的异染色质组织。 单元格。 分子生命科学。 62, 2669-2678 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Cong Y.,Shay J.W.(2008)。 人类端粒酶在端粒以外的作用。 细胞研究。 18, 725-732 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Crabbe L.、Jauch A.、Naeger C.M.、Holtgref-Grez H.、Karlseder J.(2007年)。 端粒功能障碍是沃纳综合征基因组不稳定的一个原因。 程序。 国家。 阿卡德。 科学。 美国 104, 2205-2210 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Davoli T.、Denchi E.L.、de Lange T.(2010年)。 持续的端粒损伤导致有丝分裂旁路和四倍体。 单元格 141, 81-93 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
De Sandre Giovannoli A.、Bernard R.、Cau P.、Navarro C.、Amiel J.、Boccaccio I.、Lyonnet S.、Stewart C.L.、Munnich A.、Le Merrer M.等人(2003年)。 拉明在哈钦森-吉尔福德早熟症中被截断。 科学类 300, 2055 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
De Semir D.、Nosrati M.、Li S.、Kashan-Sabet M.(2007年)。 端粒酶:超越末端。 细胞周期 6, 546-549 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Dechat T.、Pfleghar K.、Sengupta K.、Shimi T.、Shumaker D.K.、Solimando L.、Goldman R.D.(2008)。 核层粘连蛋白:细胞核和染色质结构组织和功能的主要因素。 基因发育。 22, 832-853 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Decker M.L.、Chavez E.、Vulto I.、Lansdorp P.M.(2009年)。 Hutchinson-Gilford早衰综合征的端粒长度。 机械。 老龄化发展。 130, 377-383 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Dimri G.P.、Lee X.、Basile G.、Acosta M.、Scott G.、Roskelley C.、Medrano E.E.、Linskens M.、Rubelj I.、Pereira-Smith O.等人(1995年)。 一种生物标记物,用于识别培养中和体内老化皮肤中的衰老人类细胞。 程序。 国家。 阿卡德。 科学。 美国 92, 9363-9367 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Eriksson M.、Brown W.T.、Gordon L.B.、Glynn M.W.、Singer J.、Scott L.、Erdos M.R.、Robbins C.M.、Moses T.Y.、Berglund P.等人(2003年)。 层粘连蛋白A的复发性新发点突变导致Hutchinson-Gilford早衰综合征。 自然 423, 293-298 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Goldkorn A.,Blackburn E.H.(2006年)。 将突变模板端粒酶RNA组装成可作用于端粒的催化活性端粒酶核糖核蛋白是人类癌症细胞凋亡和细胞周期阻滞所必需的。 癌症研究。 66, 5763-5771 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Goldman R.D.、Gruenbaum Y.、Moir R.D.和Shumaker D.K.、Span T.P.(2002)。 核层压板:核结构的构件。 基因发育。 16, 533-547 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Goldman R.D.、Shumaker D.K.、Erdos M.R.、Eriksson M.、Goldman A.E.、Gordon L.B.、Gruenbaum Y.、Khuon S.、Mendez M.、Varga R.等人(2004年)。 突变层粘连蛋白A的积累导致Hutchinson-Gilford早衰综合征核结构的进行性改变。 程序。 国家。 阿卡德。 科学。 美国 101, 8963-8968 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Gonzalez-Suarez I.、Redwood A.B.、Perkins S.M.、Vermolen B.、Lichtensztejin D.、Grotsky D.A.、Morgado-Palacin L.、Gapud E.J.、Sleckman B.P.、Sullivan T.等人(2009年)。 A型层粘连蛋白在端粒生物学和DNA损伤反应途径中的新作用。 EMBO J。 28, 2414-2427 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Gruenbaum Y.、Margalit A.、Goldman R.D.、Shumaker D.K.和Wilson K.L.(2005年)。 核膜成熟。 自然修订版分子细胞生物学。 6, 21-31 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Hande M.P.、Balajee A.S.、Tchirkov A.、Wynshaw-Boris A.、Lansdorp P.M.(2001)。 Atm(−/−)小鼠和共济失调-扩张症患者细胞中的染色体外端粒DNA。 嗯,分子遗传学。 10, 519-528 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Harley C.B.、Sherwood S.W.(1997年)。 培养的人类皮肤成纤维细胞的老化。 方法分子生物学。 75, 23-30 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Hennekam R.C.(2006)。 Hutchinson-Gilford早衰综合征:表型回顾。 美国医学遗传学杂志。 A类 140, 2603-2624 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Huang S.、Risques R.A.、Martin G.M.、Rabinovitch P.S.和Oshima J.(2008)。 突变型和野生型层粘连蛋白A过度表达的人成纤维细胞的端粒缩短和复制性衰老加速。 实验细胞研究。 314, 82-91 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Karlseder J.、Smogorzewska A.、de Lange T.(2002)。 端粒状态改变引起的衰老,而不是端粒丢失。 科学类 295, 2446-2449 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Kim M.、Xu L.、Blackburn E.H.(2003)。 具有等位基因特异生物学特性的催化活性人类端粒酶突变体。 实验细胞研究。 288, 277-287 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Kudlow B.A.、Stanfel M.N.、Burtner C.R.、Johnston E.D.、Kennedy B.K.(2008)。 通过hTERT或p53失活抑制与加工缺陷型层粘连蛋白A突变体相关的增殖缺陷。 分子生物学。 单元格 19, 5238-5248 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Lee J.、Sung Y.H.、Cheong C.、Choi Y.S.、Jeon H.K.、Sun W.、Hahn W.C.、Ishikawa F.、Lee H.W.(2008)。 TERT独立于端粒酶活性促进细胞和生物体的存活。 癌基因 27, 3754-3760 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Limon A.、Reyes-Ruiz J.M.、Eusebi F.、Miledi R.(2007年)。 氨基或羧基末端GFP标记的GluR3受体的特性。 程序。 国家。 阿卡德。 科学。 美国 104, 15526-15530 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
刘斌、王杰、陈国美、丁家伟、邓伟、关旭、黄建德、李国美、周培云、陈建杰等(2005)。 基于椎板病的早衰的基因组不稳定性。 自然医学。 11, 780-785 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Liu Y.,Rusinol A.,Sinensky M.,Wang Y.,Zou Y.(2006)。 进行性综合征中的DNA损伤反应源于前层蛋白A成熟缺陷。 细胞科学杂志。 119, 4644-4649 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Louis S.F.、Vermolen B.J.、Garini Y.、Young I.T.、Guffei A.、Lichtensztejn Z.、Kuttler F.、Chuang T.C.、Moshir S.、Mougey V.等人(2005年)。 c-Myc通过端粒和间期核的染色体重塑诱导染色体重排。 程序。 国家。 阿卡德。 科学。 美国 102, 9613-9618 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Ly D.H.、Lockhart D.J.、Lerner R.A.、Schultz P.G.(2000年)。 有丝分裂失调与人类衰老。 科学类 287, 2486-2492 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Mahale A.M.、Khan Z.A.、Igarashi M.、Nanjangud G.J.、Qiao R.F.、Yao S.、Lee S.W.、Aaronson S.A.(2008)。 用确定的细胞癌基因转导的人成纤维细胞恶性转化中的克隆选择。 癌症研究。 68, 1417-1426 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Maslov A.Y.、Vijg J.(2009年)。 基因组不稳定、癌症和衰老。 生物化学。 生物物理学。 学报 1790, 963-969 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
McClintock D.、Ratner D.、Lokuge M.、Owens D.M.、Gordon L.B.、Collins F.S.、Djabali K.(2007)。 导致Hutchinson-Gilford早衰的层粘连蛋白A突变形式是人类皮肤细胞老化的生物标志物。 公共科学图书馆 2,e1269 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Merideth M.A.、Gordon L.B.、Clauss S.、Sachdev V.、Smith A.C.、Perry M.B.、Brewer C.、Zalewski C.、Kim H.J.、Solomon B.等人(2008年)。 Hutchinson-Gilford早衰综合征的表型和病程。 北英格兰。 医学杂志。 358, 592-604 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Michishita E.、McCord R.A.、Berber E.、Kioi M.、Padilla-Nash H.、Damian M.、Cheung P.、Kusumoto R.、Kawahara T.L.、Barrett J.C.等人(2008年)。 SIRT6是一种组蛋白H3赖氨酸9脱乙酰酶,调节端粒染色质。 自然 452, 492-496 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Mukherjee A.B.,Costello C.(1998年)。 体外细胞老化过程中FISH对人类早衰综合征成纤维细胞的非整倍体分析。 机械。 老龄化发展。 103, 209-222 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Ouellette M.、McDaniel L.D.、Wright W.E.、Shay J.W.、Schultz R.A.(2000年)。 代表人类染色体不稳定综合征的端粒酶永久化细胞系的建立。 嗯,分子遗传学。 9, 403-411 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Park J.I.、Venteicher A.S.、Hong J.Y.、Choi J.、Jun S.、Shkreli M.、Chang W.、Meng Z.、Cheung P.、Ji H.等人(2009年)。 端粒酶通过与靶基因染色质结合调节Wnt信号。 自然 460, 66-72 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Pegoraro G.、Kubben N.、Wickert U.、Gohler H.、Hoffmann K.、Misteli T.(2009年)。 NURD复合物丢失导致的与年龄相关的染色质缺陷。 自然细胞生物学。 11, 1261-1267 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Puzianowska Kuznicka M.,Kuznicki J.(2005年)。 加速老化综合征的遗传改变。 它们在自然衰老中起作用吗? 国际生物化学杂志。 细胞生物学。 37, 947-960 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Raz V.、Vermolen B.J.、Garini Y.、Onderwater J.、Mommaas-Kienhuis M.A.、Koster A.J.、Young I.T.、Tanke H.、Dirks R.W.(2008)。 核膜在人类间充质干细胞衰老过程中促进端粒聚集和着丝粒外周定位。 细胞科学杂志。 121, 4018-4028 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Riches L.C.、Lynch A.M.、Goodeham N.J.(2008年)。 哺乳动物对DNA双链断裂反应的早期事件。 突变 23, 331-339 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Scaffidi P.、Misteli T.(2006年)。 人类衰老过程中的层粘连蛋白A依赖性核缺陷。 科学类 312, 1059-1063 16645051 [ 谷歌学者 ]
Scaffidi P.、Misteli T.(2008)。 与加速衰老相关的成人干细胞的层粘连蛋白A依赖性失调。 自然细胞生物学。 10, 452-459 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Shumaker D.K.、Dechat T.、Kohlmaier A.、Adam S.A.、Bozovsky M.R.、Erdos M.R、Eriksson M.、Goldman A.E.、Khuon S.、Collins F.S.等人(2006年)。 封面:突变的核层粘连蛋白A导致早衰表观遗传控制的渐进性改变。 程序。 国家。 阿卡德。 科学。 美国 103, 8703-8708 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Skube S.B.、Chaverri J.M.、Goodson H.V.(2009年)。 GFP标签对EB1和EB1片段在体内定位的影响。 细胞模型。 细胞骨架 67, 1-12 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Smith L.L.、Coller H.A.和Roberts J.M.(2003)。 端粒酶调节生长控制基因的表达并促进细胞增殖。 自然细胞生物学。 5, 474-479 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Smogorzewska A.、van Steensel B.、Bianchi A.、Oelmann S.、Schaefer M.R.、Schnapp G.、de Lange T.(2000)。 通过TRF1和TRF2控制人类端粒长度。 分子细胞。 生物。 20, 1659-1668 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Stuurman N.、Heins S.、Aebi U.(1998年)。 核层粘连蛋白:它们的结构、组装和相互作用。 J.结构。 生物。 122, 42-66 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Taddei A.、Hediger F.、Neumann F.R.、Gasser S.M.(2004)。 核结构的功能:遗传学方法。 每年。 修订版Genet。 38, 305-345 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Takai H.、Smogorzewska A.、de Lange T.(2003)。 DNA损伤集中在功能失调的端粒。 货币。 生物。 13, 1549-1556 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Tewey K.M.、Rowe T.C.、Yang L.、Halligan B.D.、Liu L.F.(1984年)。 阿霉素诱导哺乳动物DNA拓扑异构酶II介导的DNA损伤。 科学类 226, 466-468 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Toth J.I.、Yang S.H.、Qiao X.、Beigneux A.P.、Gelb M.H.和Moulson C.L.、Miner J.H.,Young S.G.和Fong L.G.(2005)。 阻断蛋白法尼基转移酶改善进展综合征患者成纤维细胞的核形状。 程序。 国家。 阿卡德。 科学。 美国 102, 12873-12878 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Vidal A.,Koff A.(2000年)。 细胞周期抑制剂:三个家族因一个共同的原因团结在一起。 基因 247, 1-15 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Wallis C.V.、Sheerin A.N.、Green M.H.、Jones C.J.、Kipling D.、Faragher R.G.(2004)。 尽管存在端粒酶,Hutchinson-Gilford早衰患者的成纤维细胞克隆仍会衰老。 实验Gerontol。 39, 461-467 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
王海斌、关建生、鲍立、张欣(2008)。 delta-opioid受体在PC12细胞中的独特亚细胞分布与各种标签融合。 神经化学。 物件。 33, 2028-2034 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
周磊、郑德、王明、聪颖S.(2009)。 端粒酶逆转录酶独立于端粒酶活性激活血管内皮生长因子的表达。 生物化学。 生物物理学。 Res.Commun公司。 386, 739-743 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]