方法
参与者
39名参与者完成了研究(20名女性;平均年龄21.8±3.4岁;年龄范围18-36岁;平均体重72.6±13.0 kg;年龄范围47.0-98.2 kg),其中36名参与者完成神经成像疗程(20名接受内毒素治疗,16名接受安慰剂治疗;每种情况下,男性和女性人数相等)。未接受扫描的三名参与者是试验参与者,他们接受了内毒素检查,但没有完成神经成像课程,以确保参与者能够离开加州大学洛杉矶分校临床研究中心(GCRC),在没有护士监督的情况下完成神经成像课。
参与者是从加州大学洛杉矶分校医学中心发布的传单和校园报纸上发布的广告中招募的。通过结构化电话访谈,患有以下症状的潜在参与者被排除在外:幽闭恐怖症或体内金属(与研究的神经成像部分相关)、慢性精神或身体疾病、严重的身体或心理健康问题、过敏史、自身免疫性疾病、肝脏、,或其他严重的慢性病,目前使用的处方药,夜班工作或前6周内的时区轮班(>3小时)。参与者也必须是右撇子。
电话面试后,如果仍然合格,参与者将完成额外的面对面筛选面试,以确保有资格参加研究。参与者因完成此次筛选会议而获得20美元。在这一环节中,一位经过培训的采访者与参与者一起回顾了研究程序,包括内毒素的后果和可能出现的症状类型。在此之后,参与者被问及一系列关于身体健康和药物使用的问题。参与者随后完成了DSM障碍结构化临床访谈(SCID;First等人,1996年)确保他们没有心理健康问题。最后,测量身高和体重,评估生命体征(如体温、心率、血压),收集尿样以检查药物使用情况(大麻、鸦片制剂、可卡因、安非他明、甲基苯丙胺),如果是女性,则抽血筛查怀孕情况。进行了筛查实验室检查,包括完整的血细胞计数、化学组和肝功能测试。此外,任何参与者:1)BMI大于30,2)报告身体健康问题或药物使用,3)根据SCID评估证明存在Axis I精神障碍,4)尿检呈阳性,5)妊娠试验呈阳性,如果是女性,或6)显示其筛选实验室测试中的任何异常均不符合研究条件。最后的样本是39%的欧美人、18%的亚洲人、18%的西班牙裔、7%的非洲裔美国人和18%的“其他人”。实验程序由加州大学洛杉矶分校人类受试者保护委员会批准。
程序
该研究在加州大学洛杉矶分校GCRC进行,采用随机、双盲、安慰剂对照设计。参与者于上午7:30(n=31)或8:30(n=8)到达加州大学洛杉矶分校GCRC,并被分配到单人或双人房间的床上。到达后,收集尿样并进行药物使用和妊娠测试,以确保继续有资格参与研究。没有参与者的测试结果呈阳性。完成后,参与者开始研究程序。
GCRC的一名护士对这种情况一无所知,她获得了基线生命体征(血压、脉搏、体温)以及身高和体重。然后,护士将一根带肝素锁的导管插入主要前臂(右),每小时抽血一次,将一根导管插入非主要前臂,进行连续盐水冲洗和给药。导管就位后,护士开始滴生理盐水,以保持参与者水分充足(1000cc生理盐水,150cc/hr);在整个研究过程中,除了参与者离开去完成神经成像疗程外,都在继续输注生理盐水。参与者在服用药物或安慰剂之前完成一些人口统计学调查问卷并吃一顿清淡的早餐,以此开始这项研究。
到达GCRC后90分钟,每位参与者被随机分配接受内毒素(0.8纳克/千克体重)或安慰剂(相同体积的0.9%生理盐水),由护士静脉注射。本研究中使用的内毒素来源于大肠杆菌(大肠杆菌O组:113),由美国国立卫生研究院临床中心提供,作为人体实验性炎症研究的参考内毒素(Suffredini等人,1999年). 以前的研究已经证明在许多不同的样品中安全使用这种参考内毒素(Andreasen等人,2008年;苏弗雷迪尼和奥格雷迪,1999年). 两组之间在年龄、教育年限或体重方面没有发现显著差异。
在整个研究过程中,每半小时对生命体征(血压、脉搏和体温)进行一次评估(神经成像期间除外),并在基线检查时采集血样(两次基线检查:上午8:30和8:45或上午9:30和9:45,取决于开始时间)然后在接下来的六个小时里,大约每小时一次。随后对这些抽血进行了IL-6和皮质醇水平的检测,此前已证明,这些水平随着内毒素的作用而升高(Reichenberg等人,2001年). 此外,随后对基线抽血进行雌二醇和孕酮水平分析,以控制女性月经周期的阶段。参与者还完成了每次抽血时身体症状(如肌肉疼痛、疲劳)和抑郁情绪的自我报告测量。
注射后约两小时,即先前研究显示促炎细胞因子达到峰值时(Krabbe等人,2005年;Reichenberg等人,2001年;Suffredini等人,1999年;Wright等人,2005年),参与者完成了一次神经移植课程。参与者被护送至加州大学洛杉矶分校大脑映射中心,在那里他们在扫描仪中完成了Cyberball社交排斥任务(详见fMRI范式)。神经成像课程结束后,参与者返回GCRC,吃午饭,并完成剩余的研究程序。出于安全原因,研究医生(M.I.)知道每个参与者的分组,但没有参与测试程序。研究医生在每次实验期间都会随时待命。经研究医生批准,参与者在最后一次抽血后从GCRC出院;如果自我报告的身体和心理症状恢复到基线水平,则获得批准。因此,所有参与者离开研究时的感觉与开始时一样好。在研究结束时,参与者得到了感谢、汇报,并为他们的参与支付了费用(200美元)。
行为评估
疾病症状
在基线检查时评估身体疾病症状(肌肉疼痛、颤抖、恶心、呼吸困难、疲劳),然后在注射内毒素或安慰剂6小时后每小时评估一次。参与者按照0(无症状)到4(非常严重的症状)的范围对他们的症状感觉程度进行评分。
抑郁情绪
抑郁情绪与疾病症状同时进行评估,使用情绪状态简表(McNair等人,1971年). 为了评估抑郁情绪,参与者对他们的感觉程度进行了评级:“不开心”、“忧郁”、“孤独”、“沮丧”和“毫无价值”,评分范围从0(一点也不)到4(极端)。抑郁情绪是通过平均每个时间点这些项目的得分来计算的。
fMRI范式
为了评估对社会排斥的神经认知反应,参与者在完成Cyberball社会排斥任务时进行了扫描,扫描方式与之前的工作类似(Eisenberger等人,2003年,2007;Williams等人,2000年). 在GCRC期间(内毒素/安慰剂给药后),参与者被告知,他们稍后将被护送到加州大学洛杉矶分校脑标测中心,在那里他们将完成神经成像课程。据解释,在本次会议期间,他们将与另外两名同样在fMRI扫描仪中的人玩一个虚拟的击球游戏。然而,事实上,没有其他玩家;参与者玩一个预设的电脑程序。为了支持参与者与其他两人一起玩的封面故事,参与者(仍在GCRC)被要求填写一份简短的问卷,在问卷中回答关于他们自己的问题(例如,“你将来想从事什么职业?”“你的爱好或其他兴趣是什么?”)并被告知,其他两名玩家会看到他们的答案,他们会在扫描之前看到其他两名球员的答案。到达加州大学洛杉矶分校脑图中心后,每个受试者阅读另外两个假定受试者对这些问题的答案;事实上,这些问卷都是提前写好的,每个受试者都读了相同的问卷。
在阅读了其他假定受试者的问卷后,参与者被安置在扫描仪中,并进行了网络游戏。每个游戏开始时,屏幕上角有两个虚拟玩家的静态图片,屏幕中下角有一只代表参与者的手。9秒后,左上角的卡通玩家开始游戏,将球扔给其他卡通玩家或参与者。参与者可以按下右手按钮盒上的两个键中的一个,将球交还给其中一名球员。Cyberball程序设置为每场比赛60次投球,电脑玩家在投球前等待0.5-3.0秒(随机确定),以增强参与者实际上是在和其他人一起玩的感觉。
在任务期间,参与者完成了两次扫描。在第一次扫描(包括)中,参与者在整个扫描期间与其他两名球员一起玩,每个虚拟球员向参与者投掷约50%的球。在第二次扫描(排除)中,参与者只接受了总共十次投掷的球,然后在剩下的扫描中,当两名球员停止向参与者投掷球时(50-70秒),参与者被排除在外。尽管在参与者之间平衡纳入和排除扫描的顺序是理想的,但这很可能改变了被试拒绝心理体验的含义。因此,为了确保有意义的排除体验,所有受试者首先完成了包括扫描。
fMRI数据采集和数据分析
数据通过西门子Allegra 3T正面扫描仪采集。用泡沫垫和外科胶带在每位参与者的额头上固定头部运动。每个参与者的高分辨率结构T2加权回波平面成像体积(自旋回波;TR=5000ms;TE=33ms;矩阵大小128×128;36个轴向切片;视野=20-cm;3-mm厚,跳跃1-mm)与功能扫描共面。采集两次功能扫描(回波平面T2*加权梯度回波,TR=2000ms,TE=25ms,翻转角度=90°,矩阵尺寸64×64,36个轴向切片,视野=20-cm;3-mm厚,跳跃1-mm),每次持续2分48秒。
使用SPM’5(英国伦敦神经病学研究所威康认知神经病学系)对影像数据进行分析。对每个受试者的图像进行重新校准,以纠正头部运动,将其归一化为标准立体定向空间,并使用8mm高斯核(最大宽度为一半)进行平滑处理,以提高信噪比。该设计采用与典型血流动力学响应函数卷积的箱形函数进行建模。对于每个参与者,根据参与者的包含和排除事件的长度,将包含和排除的时间建模为时间段(由于虚拟球员在投球时的随机延迟以及受试者自己返回投球的反应时间,参与者之间的时间略有不同)。在对每个参与者的任务进行建模后,将计划的比较计算为线性对比,以调查排除事件与包含事件之间的神经活动。使用为每个参与者生成的对比度图像计算该组的随机效应分析。
血浆IL-6、皮质醇、雌二醇和孕酮水平
血浆样本收集在含有乙二胺四乙酸钠和抑肽酶的预冷试管中,立即离心、校准并放置在-70°F的冰箱中。采用高灵敏度酶联免疫吸附测定法(明尼苏达州明尼阿波利斯R&D Systems)定量血浆IL-6水平。所有样品均重复运行,并在同一时间进行分析,单个操作员使用单个批次的试剂和消耗品,所有变量的组内变异系数均小于5%。使用DPC IMMULITE 1000免疫分析化学分析仪(美国加利福尼亚州诊断产品公司)测量血浆皮质醇水平。IMMULITE 1000皮质醇分析是一种固相竞争性化学发光酶免疫分析,根据制造商的说明进行。该分析的分析灵敏度为0.2μg/dL(5.5 nmol/L)。对基线时采集的血浆样本进行雌二醇和孕酮的激素评估。使用涂层管放射免疫分析试剂盒。使用的特异性试剂盒为DSL-4300 ACTIVE®雌二醇包被管放射免疫试剂盒和DSL-5000 ACTIVE®17αOH孕酮包被管放射免疫试剂盒。
统计分析
行为结果
为了评估内毒素与安慰剂对IL-6水平、皮质醇水平、身体疾病症状(如疲劳)、生命体征(如心率、血压、体温的变化)和自我报告的抑郁情绪影响的组间差异,我们使用标准统计软件程序(SPSS)进行重复测量方差分析。由于IL-6水平在任何时间点都不是正态分布,因此每个值都是对数转换的。
为了评估内毒素状态下白细胞介素-6(IL-6)的增加与相关因变量(从基线检查到注射后两小时,即神经成像之前)的增加之间的相关性,计算了皮尔逊相关系数。同样,由于IL-6水平不是正态分布的,我们通过注射后两小时的对数转换值减去对数转换基线值来计算差异分数。
神经影像学结果
为了评估内毒素与安慰剂对赛博游戏神经反应性影响的组间差异,在SPM’5中计算了一个单向方差分析,以对比内毒素组与对照组在排除期间相对于纳入期间的神经活动(p<0.001,10-体素范围阈值;Forman等人1995年). 为了探讨内毒素组IL-6升高与神经反应之间的关系,我们进行了全脑回归分析,以了解哪些神经区域与IL-6升高相关。IL-6升高的测量值(从基线到注射后两小时)作为回归因子输入到随机效应全脑组分析(仅针对内毒素条件下的患者)中,比较排除期间的激活与纳入期间的激活(p<0.001,10体素范围阈值)。由于我们对理解在细胞因子相关抑郁情绪中起作用的神经回路感兴趣,我们进一步研究了与IL-6升高显著相关的神经区域,以了解它们是否也与抑郁情绪的变化相关。此外,由于众所周知的性别差异,抑郁症和炎症性疾病的发病率也存在差异(Nolen-Hoeksema,2001年;Whitacre等人,1999年)这些分析也分别针对男性和女性进行。所有坐标均以蒙特利尔神经研究所(MNI)的格式报告。
结果
内毒素的生理和行为反应
与安慰剂相比,内毒素导致除最后一个时间点外的所有时间点的IL-6水平显著增加(注射后2-3小时达到峰值),与基线(T1(注射后1小时)到T5(注射后5小时)相比,每个时间点的显著时间X条件交互作用证明了这一点:F(1,37)=23.19、144.14、177.11、63.67、11.36,所有p<.005;图1a). 与安慰剂相比,内毒素还导致除基线检查后第一个时间点外的所有测量时间点的皮质醇水平显著增加(注射后3小时达到峰值;T2-T6时间X条件相互作用:F(1,37)=21.83、52.78、20.59、10.29、7.96,所有p均<0.01)。此外,与安慰剂相比,内毒素在所有测量时间点都显著增加了自我报告的身体疾病症状(注射后2小时达到峰值;T1-T6的时间X条件相互作用:F(1,37)=9.08,32.37,13.60,8.54,11.65,8.01,注射后3小时开始,体温显著升高(时间X条件相互作用:T3-T6:F(1,37)=14.18,17.14,17.93,5.70,所有p<.05)和脉搏显著升高(温度X条件相互影响:T3-T7:F(1,33)=48.43,24.01,23.74,14.59,所有p<.001)。最后,我们发现内毒素组与安慰剂组相比,注射后两小时自我报告的抑郁情绪显著增加(时间x条件交互作用:F(1,35)=8.13,p<0.01;图1b). 在这些影响中没有任何性别差异。此外,这些影响似乎不是由于月经周期的阶段,因为在控制了作为月经周期阶段标志物的雌二醇和孕酮的基线水平(无论是在完整样本中还是在单独检查女性时)后,这些影响都没有改变。
图1。
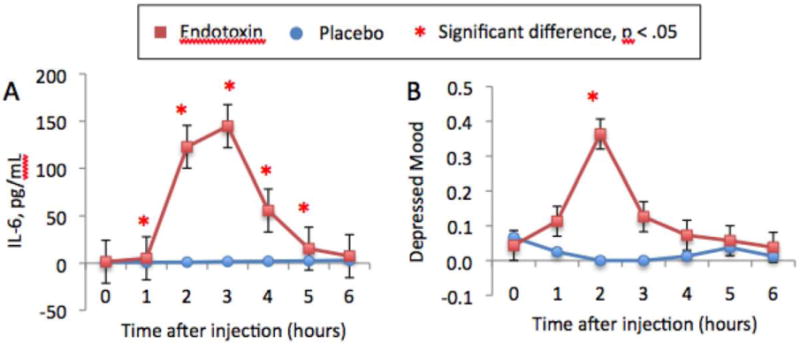
随时间变化:A)血浆IL-6水平(这些是未经对数转换的原始值)和B)自我报告的抑郁情绪。带星号的时间点表示重要的时间X条件交互。这些数字单独公布,并根据Eisenberger等人(2009年)改编。
IL-6变化与抑郁情绪变化的相关性
为了检测IL-6的增加是否与自我报告的抑郁情绪的增加相关,我们检测了IL-6从基线水平到注射后两小时(就在神经成像治疗之前)的增加之间的相关性以及同一时间段内抑郁情绪的自我报告变化(这两项指标均以差异分数计算)。对于接触内毒素的受试者,IL-6的增加和抑郁情绪的增加之间没有显著相关性。然而,当样本按性别分层时,IL-6水平的增加与女性自我报告的抑郁情绪的增加显著相关(r=.70,p<0.05),而男性则没有相关性(r=-.20;p=.59);这些相关性之间存在显著差异(Z=2.12,p<0.05)。此外,对于女性,当自我报告的身体症状(肌肉疼痛、颤抖、恶心、呼吸困难、疲劳)的增加得到控制时,IL-6的增加和抑郁情绪的增加之间的关系没有改变(r=.59,p=.05)。类似地,控制温度或脉冲的增加也没有显著改变这种关系(分别为r=.71,p<.05;r=.73,p<.05)。在控制了基线水平的雌二醇和孕酮后,IL-6增加与抑郁情绪之间的相关性也保持不变(r=.74,p<0.001)。
社会排斥的神经反应
群体间神经活动差异与社会排斥
为了研究促炎细胞因子激活对社会排斥的神经反应的影响,我们首先检查了内毒素组与安慰剂组在社会排斥反应(与纳入反应相比)方面不同的神经区域。总的来说,与内毒素组和安慰剂组的纳入相比,排除期间社会疼痛相关神经回路没有显著差异。在枕皮质的一个区域发现了各组间排除与包含的神经活动的唯一差异(安慰剂>内毒素:52,-78,6,t=3.70,k=117;p<0.001)。当分别检查每种性别时,观察到类似的效果;与安慰剂组相比,无论是男性还是女性,内毒素组都没有表现出更大的神经活性。与内毒素组相比,安慰剂组的女性在一些通常不参与社会疼痛过程的神经区域表现出更强的活动性(左腹外侧前额叶皮层:-34、32、-8、t=5.24、k=14、p<0.001;枕叶皮层:20、-72、30、t=4.81、k=16、p<0.01和48、-78、0、t=4.31、k=21、p<0.00)。
考虑到IL-6对内毒素的反应存在大量的变异性,这种组间差异的缺乏并不令人惊讶(参见图1a; 范围:17.52–356.03 pg/ml),这一发现已在之前得到证实(Stephens等人,2005年). 为了利用这种可变性,我们接下来研究了IL-6对内毒素的反应(内毒素组受试者之间)的个体差异与社会排斥与社会包容相比的神经反应之间的关系。此外,鉴于抑郁症和免疫状况的患病率存在众所周知的性别差异,以及在当前研究中观察到的IL-6和抑郁情绪增加之间的关系存在性别差异,还分别对每种性别的IL-6增加和神经活动之间的关系进行了研究。
内毒素组IL-6升高与神经活动的相关性
我们首先检查了内毒素组所有受试者在社会排斥期间与社会包容期间IL-6增加与神经活动之间的相关性。关于疼痛相关的神经回路,我们发现IL-6的增加(从基线到扫描前)与双侧前岛叶活动增加有关(图2a)以及更大的双侧后岛叶活动(参见表1). 此外,左后岛叶的激活与自我报告的抑郁情绪增加显著相关(r=.51,p<.05),右前岛叶的活化与自我报告抑郁情绪增加略微相关(r=0.40,p<.10)。2
图2。
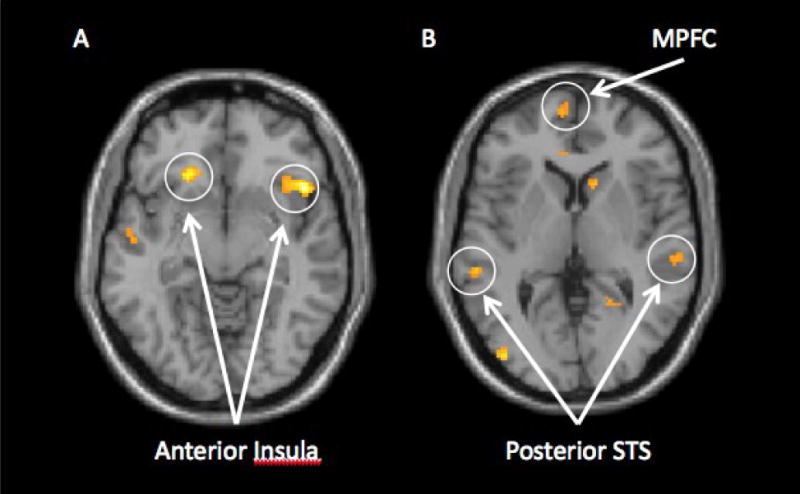
内毒素受试者在排除与纳入期间与IL-6水平增加呈正相关的神经活动(从基线到神经成像治疗前):A)双侧岛叶的活动和B)内侧前额叶皮质(MPFC)和后上颞沟(pSTS)的活动。
表1。
内毒素组所有参与者在社会排斥与包容期间的神经激活与IL-6水平从基线到注射后两小时的增加呈正相关(p<0.001,10-体素范围阈值)。没有与IL-6增加负相关的区域。R(ΔDep)是指所列神经激活的参数估计值与抑郁情绪自报增加之间的相关性。*p<0.05;S公司p<.10
| 地区 |
文学士 |
MNI坐标 |
T型 |
k个 |
R(ΔDep) |
| 边缘旁的 |
|
|
|
|
|
| 左前胰岛素 |
|
-20 28 -4 |
7.83 |
158 |
.09 |
| 右前胰岛素 |
|
48 20 -8 |
7.69 |
306 |
.40A类
|
| 左后胰岛 |
|
-46 -18 12 |
4.55 |
81 |
第51条* |
| 右后胰岛素 |
|
46 -14 12 |
4.36 |
31 |
第二十六条 |
| 膝周前扣带肌 |
32 |
-8 40 8 |
4.41 |
21 |
.33 |
| 后扣带回 |
31 |
-16 -22 36 |
4.89 |
14 |
.20 |
| 后扣带回 |
31 |
14 -40 14 |
4.14 |
12 |
.08 |
| Caudate公司 |
|
12 16 2 |
4.77 |
58 |
.41A类
|
| 前叶 |
|
|
|
|
|
| 背侧前额叶皮层(PFC) |
9 |
-8 60 40 |
6 |
52 |
.10 |
| 多索美迪尔PFC |
9 |
-10 46 54 |
5.42 |
23 |
.36 |
| 多索美迪尔PFC |
9 |
18 56 42 |
5.33 |
33 |
.09 |
| 多索美迪尔PFC |
8 |
18 24 62 |
5.22 |
23 |
.07 |
| 多索美迪尔PFC |
8 |
-2 28 54 |
4.11 |
17 |
.41A类
|
| 左背侧PFC |
9 |
-34 46 38 |
5.01 |
16 |
.17 |
| 右Rostrolateral PFC |
10 |
30 68 12 |
4.15 |
24 |
.43A类
|
| 内侧PFC |
10 |
-6 62 4 |
4.02 |
18 |
.42A类
|
| 右背前运动皮层 |
6 |
60 6 40 |
5.08 |
30 |
.25 |
| 辅助电机区 |
4/6 |
0 -26 68 |
5.58 |
73 |
2007年 |
| 辅助电机区 |
6 |
-6 8 74 |
4.62 |
12 |
.49* |
| 辅助电机区 |
6 |
12 -20 72 |
4.39 |
31 |
.20 |
| 左前运动皮层 |
4 |
-20 -22 54 |
4.29 |
13 |
第10条 |
| 顶叶 |
|
|
|
|
|
| 左侧体感皮层 |
1/2/3 |
-24 -28 64 |
4.32 |
17 |
第二十六条 |
| 早熟的 |
7 |
16 -42 66 |
4.55 |
50 |
.14 |
| 颞叶 |
|
|
|
|
|
| 右下颞回旋 |
20 |
46 -12 -30 |
4.95 |
47 |
.51* |
| 时间极点 |
38 |
-46 10 -40 |
4.65 |
22 |
.41A类
|
| 左中颞回旋 |
21 |
-54 -8 -12 |
4.40 |
58 |
.49* |
| 右中颞部陀螺仪 |
21 |
62 -10 -12 |
4.19 |
15 |
.34 |
| 右中颞部陀螺仪 |
21 |
58 -2 -16 |
4.06 |
20 |
.53* |
| 后颞上沟 |
22 |
62 -26 0 |
4.58 |
72 |
第十八条 |
| 后颞上沟 |
22 |
-58 -32 4 |
4.48 |
21 |
.17 |
| 后颞上沟 |
22 |
-52 -38 10 |
4.16 |
13 |
.57* |
| 右梭形陀螺 |
37 |
46 -42 -12 |
4.34 |
19 |
.35 |
| 枕叶 |
|
|
|
|
|
| 枕皮质 |
18 |
0-74 14 |
4.95 |
48 |
.39A类
|
| 枕皮质 |
17 |
-10 -66 14 |
4.80 |
18 |
.38 |
| 枕皮质 |
19 |
28 -54 2 |
3.88 |
15 |
.40A类
|
| 小脑 |
|
|
|
|
|
| 小脑 |
|
20 -60 -38 |
6.03 |
69 |
.23 |
| 小脑 |
|
-40 -68 -34 |
5.22 |
35 |
.15 |
| 小脑 |
|
-18 -46 -12 |
5.10 |
57 |
.42A类
|
| 小脑 |
|
-24 -56 -42 |
4.88 |
17 |
.35 |
| 小脑 |
|
-10 -60 -12 |
4.71 |
19 |
.48* |
| 小脑 |
|
8 -40 -30 |
4.67 |
42 |
.32 |
| 小脑 |
|
-4 -84 -38 |
3.91 |
10 |
.16 |
有趣的是,IL-6水平的增加也与一些神经区域的更大活动有关,这些区域与“心理化”(思考他人思想内容的过程)有关(弗里斯和弗里斯,1999年,2003,2006). 从基线检查到扫描前一刻IL-6水平增加较多的个体,其内侧和背内侧前额叶皮层(MPFC;DMPFC)的活动性更强(对社会排斥与包容的反应),这些区域与理解他人的心理状态有关(弗里斯和弗里斯,1999年,2003,2006;Mitchell等人,2005年)以及颞后上沟(pSTS)、颞极、后扣带回皮质和楔前叶的区域,所有这些区域都被证明与理解他人的思想或行为有关(图2b;弗里斯和弗里斯,1999年,2003,2006). 此外,这些区域中的一些也与自我报告的抑郁情绪增加呈正相关(MPFC:r=.42,p<.10;DMPFC:r=.41,p<.10;颞极:r=0.41,p<0.10;pSTS:r=.57,p<.05;参见表1).
内毒素组女性和男性IL-6增加与神经活动的相关性
为了进一步研究白细胞介素-6水平的增加与男性和女性对社会排斥的神经反应之间的关系,我们分别对男性和女性进行了全脑回归分析。因此,我们研究了女性内毒素受试者和男性内毒素受试人员中IL-6水平的增加与神经活动、社会排斥和包容之间的关系。
在检查内毒素组女性IL-6增加与神经活动之间的相关性时,我们发现整个大脑中只有四个显著的激活,其中三个激活位于社会疼痛相关的神经回路中(表2a). 具体而言,IL-6的增加与dACC的活性增加有关(图3a)以及左右前脑岛(图3b)与包容相比,应对社会排斥。此外,本分析中确定的dACC和右前岛叶簇也与自报告抑郁情绪从基线到注射后2小时的增加显著相关(dACC:r=.72,p<0.05;图3c; 右前岛叶:(r=.69,p<0.05;图3d). 此外,左前岛叶活动与自我报告的抑郁情绪增加略微相关(r=.58,p=.08)。只有另外一个神经区域,即躯体感觉皮层,与IL-6增加呈负相关(参见表2a). 在控制了雌二醇或孕酮的基线水平后,这些结果都没有改变。
表2。
从基线检查到注射后两小时,社会排斥与纳入期间的神经激活与IL-6水平增加相关:A)内毒素组的所有女性和B)内毒素组所有男性(p<0.001,10-体素范围阈值)。R(ΔDep)是指所列神经激活的参数估计值与抑郁情绪自报增加之间的相关性。*p<0.05;S公司p<.10。
| 地区 |
文学士 |
MNI坐标 |
T型 |
k个 |
R(ΔDep) |
| A) 女性内毒素参与者 |
|
|
|
|
|
| 与IL-6增加呈正相关 |
|
|
|
|
|
| 边缘旁的 |
|
|
|
|
|
| 左前胰岛素 |
|
-22 32 4 |
6.06 |
17 |
.58A类
|
| 右前胰岛素 |
|
42 22 -2 |
5.70 |
19 |
.69* |
| 背前扣带回(ACC) |
32 |
-8 22 46 |
5.82 |
17 |
.72* |
| 颞叶 |
|
|
|
|
|
| 右梭状回 |
37 |
44 -40 -12 |
8.57 |
30 |
.55 |
| 与IL-6增加呈负相关 |
|
|
|
|
|
| 右侧体感皮层 |
1/2/3 |
64 -16 44 |
6.07 |
11 |
-.41 |
| B) 男性内毒素参与者 |
|
|
|
|
|
| 与IL-6升高呈正相关 |
|
|
|
|
|
| 边缘边缘的 |
|
|
|
|
|
| 右前胰岛素 |
|
36 18 -12 |
7.75 |
60 |
.03 |
| 左前胰岛素 |
|
-20 28 -6 |
7.64 |
21 |
-.21 |
| 右后胰岛素 |
|
46 -16 12 |
12.02 |
62 |
-.25 |
| 左后胰岛 |
|
-36 -12 12 |
6.39 |
34 |
-.12 |
| Dorsal/Rostral ACC公司 |
32 |
12 34 24 |
6.79 |
23 |
-.09 |
| 前叶 |
|
|
|
|
|
| 左腹外侧前额叶皮层 |
44 |
-60 4 34 |
9.87 |
12 |
-.09 |
| 右腹外侧前额叶皮层 |
47 |
46 20 -12 |
7.20 |
14 |
.00 |
| 背侧前额叶皮层 |
8 |
18 56 40 |
6.26 |
19 |
-.24 |
| 右背前运动皮层 |
6 |
54 4 42 |
7.09 |
19 |
-.09 |
| 辅助电机区 |
6 |
10 -18 72 |
5.14 |
10 |
-.07 |
| 顶叶 |
|
|
|
|
|
| 左侧体感皮层 |
1/2/3 |
-40-16 32 |
8.39 |
27 |
-.02 |
| 颞叶 |
|
|
|
|
|
| 后颞上沟 |
22 |
-58 -42 18 |
11.85 |
46 |
-.31 |
| 左中颞回旋 |
21 |
-56 -4 -22 |
7.84 |
38 |
-.08 |
| 右下颞回旋 |
20 |
56 -20 -24 |
5.97 |
19 |
-.15 |
| 枕叶 |
|
|
|
|
|
| 枕皮质 |
18 |
-40 -80 0 |
8.51 |
50 |
-.35 |
| 小脑 |
|
|
|
|
|
| 小脑 |
|
-10 -48 -20 |
10.91 |
100 |
-.23 |
| 小脑 |
|
14 -42 -22 |
8.16 |
128 |
-.03 |
| 小脑 |
|
22 -72 -44 |
6.15 |
11 |
第十八条 |
| 小脑 |
|
-8 -54 -38 |
6.07 |
20 |
.02 |
| 小脑 |
|
22 -58 -40 |
5.94 |
61 |
-.22 |
图3。
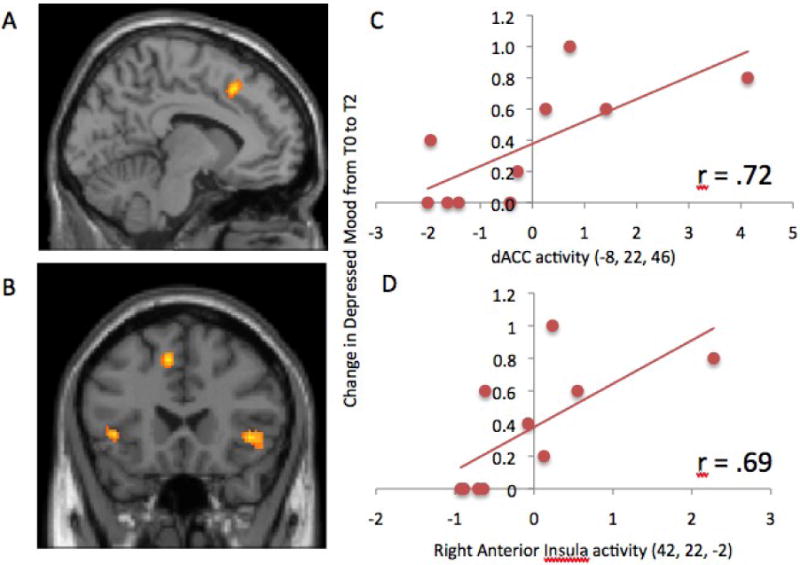
A)背前扣带回皮层(dACC;-8,22,46)和B)双侧前岛叶(左:-22,32,4;右:42,22,-2)的活性与内毒素组雌性大鼠从基线到注射后2小时IL-6水平的增加相关。散点图显示内毒素组女性中C)dACC活性和D)右前岛叶活性与抑郁情绪从基线到注射后2小时增加之间的关系。
因为IL-6增加与自我报告的抑郁情绪增加之间以及dACC和右前岛叶活动与这两个变量之间存在显著相关性,我们进行了统计中介分析,以了解这些区域的活动是否介导了女性IL-6增加和自我报告抑郁情绪增加之间的关系。发现dACC和右前岛叶均显著或略微显著地介导了IL-6升高和抑郁情绪之间的关系(dACC:Sobel试验=1.71,p<0.05,单尾;右前岛:Sobel测试=1.34,p=0.09,单尾);MacKinnon等人,2002年);这表明,对于女性来说,IL-6可能与抑郁情绪有关,部分是通过社会疼痛相关的神经活动。
当研究内毒素组男性IL-6升高与神经活动之间的相关性时,我们再次发现IL-6升高与几个不同的社会疼痛处理区域相关,包括前岛叶和后岛叶以及ACC的一个区域(尽管这种激活比在女性身上观察到的更为明显;参见表2b). 然而,这些激活与抑郁情绪的增加没有显著相关(所有p>.16)。此外,对于男性,而不是女性,IL-6水平的增加与几个前额叶调节区域(右侧和左侧腹外侧前额叶皮层(rVLPFC,lVLPFC)以及几个涉及精神化的区域(DMPFC,pSTS)的更大激活相关。然而,这些区域中没有一个与自我报告的抑郁情绪增加显著相关。有趣的是,rVLPFC中的活动与情绪和疼痛调节有关(Eisenberger等人,2003年;利伯曼等人,2004年;Ochsner&Gross,2005年;Wager等人,2004年)与右侧前岛叶的激活呈负相关(34,24,14,r=-.89,p<0.01),该区域与右侧前岛屿的激活相似,略微介导了女性IL-6增加与抑郁情绪增加之间的关系(42,22,-2)。
讨论
目前的研究旨在通过检测这些影响的一些可能的神经认知底物,进一步了解促炎细胞因子活性与抑郁情绪之间的新出现的关系。具体来说,我们对促炎细胞因子活性是否有助于提高社会疼痛敏感性感兴趣,这可能会增加抑郁症状的易感性。为了做到这一点,我们通过暴露于内毒素或安慰剂来实验性操纵促炎细胞因子活性,然后测量自我报告的抑郁症状以及对社会排斥事件的神经反应。虽然我们重复了以前的研究(Reichenberg等人,2001年)与安慰剂相比,内毒素导致抑郁情绪增加,我们没有发现两组对排斥与包容的神经反应存在显著差异。相反,出现了性别差异,例如,对于女性来说,社会疼痛相关的神经活动介导了IL-6增加和抑郁情绪增加之间的关系;而对于男性,IL-6升高与抑郁情绪之间没有显著关系。
基于对社会排斥的神经反应缺乏群体间差异,人们可能得出结论,实验性炎症不会改变社会疼痛敏感性。然而,我们的研究结果表明,炎症反应的生物变异性,如这里通过循环IL-6浓度测量的,在与神经反应差异相关方面可能比实验挑战本身更重要。事实上,IL-6对内毒素反应的变异性与神经对社会排斥的反应有关,因此IL-6增加越大,前岛叶和后岛叶活动越大,这两个区域都参与生理和社会疼痛处理(Apkarian等人,2005年;Aziz等人,2000年;Cechetto和Saper,1987年;Eisenberger等人,2003年). 此外,这两个区域的活动都与抑郁情绪的增加有关。然而,当根据本分析中的神经激活进行参数估计并分别检查其与每种性别抑郁情绪的相关性时,在全内毒素样本中,岛叶活动与抑郁情绪之间的相关性似乎主要由女性驱动(左后岛叶:女性r=.70,p<0.05;男性r=.33,纳秒; 右侧前岛叶:女性r=.77,p<0.05;男性r=0.03,纳秒). 因此,尽管对于内毒素暴露的受试者来说,较高的IL-6对内毒素的反应与较高的社会疼痛相关神经活动有关,但这些神经激活与抑郁情绪之间的关系似乎仅限于内毒素暴露的女性。这与下面讨论的针对每种性别分别进行的神经分析一致。
出乎意料的是,暴露于内毒素的人群中IL-6水平的增加也与一些已知在“心理化”或理解他人的心理状态和行为中起作用的神经区域的活动增强有关(弗里斯和弗里斯,1999年,2003,2006;Mitchell等人,2005年). 因此,炎症水平的升高可能与在具有社会威胁的情况下更努力地处理他人的目标和意图有关,可能是为了更好地预测他人的行为,从而在生理功能受损时保护自己。这与疾病行为的动物模型一致,在该模型中,动机资源被重新定向,以保存能量,促进康复,并确保患病动物的生存(哈特,1988). 生病时,可能需要对他人的目标和意图更加敏感,以便更好地预测他们的行为,以最小的能量消耗确保自己的安全。促炎性细胞因子活性对心理化能力的影响以前没有研究过,值得进一步研究。未来的研究将受益于纳入特定的心理化范式,以更仔细地研究炎症状态对心理化能力和趋势的影响。
细胞因子相关抑郁情绪和排斥反应的性别差异
有趣的是,促炎细胞因子活性、抑郁情绪和对社会排斥的神经反应之间的关系出现了性别差异。首先,对于暴露于内毒素的女性,IL-6水平的增加与抑郁情绪的增加显著相关,而对于暴露于毒素的男性,没有显著关系。此外,对于接触内毒素的女性,IL-6的增加几乎完全与社会疼痛相关神经活动的更大活动有关,而这种活动介导了IL-6增加和抑郁情绪增加之间的关系。对于男性,IL-6的增加与一些与社会疼痛相关的神经区域以及参与心理化和情绪调节的区域的增加相关;然而,这些区域中没有一个与自我报告的抑郁情绪增加显著相关。换句话说,虽然IL-6水平的增加与男性和女性更大的社会疼痛相关神经活动有关,但这些神经激活与抑郁情绪之间的关系是女性特有的。
目前尚不清楚是什么导致了这些性别差异;有几种不同的可能性。在文化或社会层面上,本研究中的男性不太可能报告感到抑郁,因为这与性别刻板印象背道而驰,性别刻板观念规定男性表达悲伤或抑郁的情绪在社会上不太被接受(Fabes和Martin,1991年). 尽管内毒素与安慰剂对自我报告的抑郁情绪的影响没有性别差异,但低估抑郁情绪细微变化的倾向可能使我们无法发现抑郁情绪与男性IL-6变化或对排斥的神经反应之间的显著关系。除自我报告外,未来的研究还应收集抑郁情绪的行为指数,以减少男性和女性报告特定负面情绪状态时的偏见。
在神经水平上,细胞因子相关抑郁情绪的性别差异可能部分归因于男性与女性典型参与的不同神经过程。在当前的研究中,女性主要表现出IL-6的增加与社会疼痛相关的神经活动之间的相关性,而男性表现出IL-60的增加与几种不同类型的神经活动(包括社会疼痛相关神经活动以及心理化和情绪调节神经活动)之间的相关性。事实上,对于男性来说,rVLPFC(一个参与情绪调节的区域)的活动与前岛叶的活动呈负相关,前岛叶与介导IL-6增加和女性抑郁情绪增加之间关系的岛叶激活类似。因此,男性在社会排斥期间可能参与更多的调节性神经活动,这与情感神经反应性降低有关;我们推测,这种增强的神经调节活动可能在一定程度上缓冲了男性IL-6反应、社会疼痛相关神经活动和女性抑郁情绪之间的强烈联系。然而,为什么炎症反应更强的男性会比女性招募更多的神经调节活动,目前尚不清楚。
在神经化学水平上,性别特异性激素底物可能导致细胞因子相关抑郁情绪的性别差异。基于女性抑郁症和自身免疫性疾病的较高患病率(Nolen-Hoeksema,2001年;Whitacre等人,1999年)性激素似乎是导致细胞因子相关抑郁情绪性别差异的因素。这是否属实尚不清楚。然而,基于对抑郁症性别差异的研究(Nolen-Hoeksema和Girgus,1994年)虽然我们的分析没有揭示雌二醇或孕酮水平的变化对我们的研究结果有影响,但性激素与其他生物系统结合可能导致细胞因子相关抑郁症的性别差异。
最后,炎症信号过程中的性别差异(除外周炎症外)也可能与观察到的神经差异有关。周围的炎症反应被传递到大脑,以改变神经活动和抑郁情绪。因此,除了“信号生成”(外围释放的细胞因子)外,神经和行为反应还可能反映“听到的信号”,这在一定程度上取决于炎症细胞因子(如IL-6)对大脑中炎症信号的影响。核因子κ-B(NF-κ-B)是一种协调炎症信号级联的关键转录因子,被炎症和其他挑战激活。我们推测,炎症信号传导神经反应的性别差异可能部分是由于NF-κ-B激活的差异。事实上,我们之前发现,女性而非男性在急性应激源(睡眠不足;Irwin等人,2008年)已知其可增加促炎细胞因子(Mullington等人,2009年). 需要进一步研究,以确定女性与男性相比,是否在NF-κ-B激活或其他核信号通路对内毒素的反应中表现出差异性增加。
局限性和结论
出于几个原因,应对此处报告的调查结果进行谨慎解释。首先,观察到的性别差异是基于非常小的样本量,因此未来需要进行包括更多男性和女性的研究,以确定这些性别差异是否重复。第二,尽管本研究的目的是检查促炎性细胞因子对社会排斥的神经敏感性的影响,但所有的神经发现都是基于仅在内毒素条件下受试者之间进行的相关分析。因此,尚不清楚促炎细胞因子是否在此处观察到的神经反应中起到因果作用,或者是否有第三个变量促成了细胞因子(IL-6)增加与排除神经反应之间的相关性。然而,观察到的神经活动不太可能导致IL-6增加,因为我们对IL-6增加的测量发生在神经成像治疗之前。此外,应该强调的是,内毒素已被证明能增加几种不同促炎细胞因子的产生和分泌,包括TNF-α和IL-1受体拮抗剂(Reichenberg等人,2001年). 因此,尽管我们在当前研究中仅检测了IL-6增加与神经活动之间的相关性,但其他促炎细胞因子可能显示出类似或不同的关系模式。对未来的研究来说,检查与其他促炎细胞因子的关系也很重要。
此外,与观察到的神经活动有关,尽管我们已经将脑岛或dACC等区域描述为参与社会疼痛处理,但这些区域也已被证明在许多其他过程中发挥作用(例如,厌恶、行为监测;菲利普斯等人,2003年)因此,这里观察到的活动可能不是特定于社会痛苦。然而,鉴于这些神经反应是针对被社会排斥而进行评估的,因此这些类型的激活更有可能与社会痛苦经历相关,而不是与其他类型的过程相关。最后,本研究和其他实验诱导的促炎细胞因子活性研究的固有局限性是,我们只能检测细胞因子对抑郁情绪的影响,而不能检测全面抑郁。尽管如此,了解细胞因子激活和抑郁情绪之间的因果关系可能会为我们提供有关炎症过程和抑郁之间关系的宝贵信息。
总之,实验诱导的细胞因子激活并没有全面提高社会疼痛敏感性;然而,对于女性,内毒素注射后促炎细胞因子(IL-6)增加与社会排斥导致的社会疼痛相关神经活动增加以及抑郁情绪增加有关。对于男性,内毒素注射后IL-6增加与与社会疼痛敏感性、心理化和情绪调节相关的神经活动增加有关;然而,这些激活都与抑郁情绪的变化无关。虽然还需要进一步研究来确定这些发现是否重复,但这项研究提高了我们对与细胞因子激活变化相关的社会和情感变化的理解,这些变化可能对理解促炎细胞因子活动对抑郁情绪的贡献至关重要。
致谢
作者没有与本研究结果相关的财务收益,也没有潜在的利益冲突。我们要感谢加州大学洛杉矶分校综合临床研究中心和加州大学洛杉矶分校脑映射中心的工作人员和支持。我们还要感谢美国国立卫生研究院沃伦·格兰特·马格努森临床中心的医学博士安东尼·苏弗雷迪尼(Anthony Suffredini)和R.P.乔治·格里姆斯(George Grimes)提供标准参考内毒素,感谢Thanh Luu进行细胞因子分析,感谢理查德·奥姆斯特德(Richard Olmstead)博士提供统计建议,感谢马修·利伯曼(Matthew Lieberman)博士。,征求对手稿的反馈意见。本研究由NARSAD青年研究员奖(N.I.E.)、达纳基金会拨款(N.I.E)和博士后研究奖学金(N.I.E.T32-MH19925)资助。此外,作者感谢拨款HL-079955、AG-026364、CA-10014152、CA-116778、P30-AG028748、M01-RR00865、加州大学洛杉矶分校塞梅尔神经科学研究所表亲中心、加州大学洛德·D·佩珀老年美国人独立中心炎症生物学核心、,和普通临床研究中心计划(M01-RR00865)。
脚注
1值得注意的是,炎症挑战(如内毒素)已被证明导致多种不同促炎细胞因子(如IL-6、TNF-α、IL-1受体拮抗剂)的协同增加,因此我们并不是假设内毒素的作用主要或唯一与IL-6水平相关。然而,在当前的研究中,我们检测了IL-6水平,因为之前已经证明,这种炎症标记物与抑郁的测量值相关(Howren,Lamkin&Suls,2008),并且与其他炎症标记物(例如TNF-α)一起映射以应对炎症挑战(Reichenberg等人,2001年).
2应该注意的是,尽管焦虑不是本次调查的重点,但它是通过(使用情绪状态简介的缩写版本)进行测量的(McNair等人,1971年)). 自我报告的焦虑增加与本文报告的神经激活没有显著相关性。此外,控制自我报告的焦虑增加并没有显著改变这些神经激活与自我报告的抑郁情绪增加之间的关系。
出版商免责声明:这是一份未经编辑的手稿的PDF文件,已被接受出版。作为对客户的服务,我们正在提供这份早期版本的手稿。手稿在以最终可引用的形式出版之前,将经过编辑、排版和校对结果证明。请注意,在制作过程中可能会发现可能影响内容的错误,适用于该期刊的所有法律免责声明均适用。
工具书类
-
Andreasen AS、Krabbe KS、Krogh-Madsen R、Taudorf S、Pedersen BK、Möller K。人体内毒素血症作为全身炎症模型。现代药物化学。2008;15:1697–1705. doi:10.2174/092986708784872393。[内政部] [公共医学] [谷歌学者]
-
Apkarian AV,Bushnell MC,Treede RD,Zubieta JK。健康和疾病中疼痛感知和调节的人脑机制。欧洲疼痛杂志。2005;9:463–484. doi:10.1016/j.ejpain.2004.11.001。[内政部] [公共医学] [谷歌学者]
-
Aziz Q,Schnitzler A,Enck P.内脏感觉的功能神经成像。临床神经生理杂志。2000;17:604–612. doi:10.1097/00004691-200011000-00006。[内政部] [公共医学] [谷歌学者]
-
Bonaccorso S、Metzer H、Maes M.干扰素的心理和行为影响。精神病学的当前观点。2000;13:673–677.[谷歌学者]
-
《鲍尔比J·依恋与失落》第三卷。失落、悲伤和沮丧。纽约:基础图书;1980[谷歌学者]
-
Boyce P、Parker G、Barnett B、Cooney M和Smith F。人格是抑郁的脆弱因素。英国精神病学杂志。1991;159:106–114. doi:10.1192/bjp.159.106。[内政部] [公共医学] [谷歌学者]
-
Brydon L、Harrison NA、Walker C、Steptoe A、Critchley HD。外周炎症与人类黑质活动的改变和精神运动减慢有关。生物精神病学。2008;63:1022–1029. doi:10.1016/j.biopych.2007.12.007。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Capuron L,Miller AH。细胞因子与精神病理学:干扰素-α的教训。生物精神病学。2004;56:819–824. doi:10.1016/j.biopych.2004.02.009。[内政部] [公共医学] [谷歌学者]
-
Capuron L、Pagnoni G、Demetrashvili M、Woolwine BJ、Nemeroff CB、Berns GS等。干扰素-α治疗期间的前扣带激活和错误处理。生物精神病学。2005;58:190–196. doi:10.1016/j.biopych.2005.03.033。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Cechetto DF、Saper CB。大鼠大脑皮层和丘脑内视感觉表现的证据。《计算机神经学杂志》。1987;262:27–45. doi:10.1002/cne.902620104。[内政部] [公共医学] [谷歌学者]
-
Dantzer R.细胞因子诱导的疾病行为:机制和意义。Ann N Y科学院。2001;933:222–234. doi:10.1111/j.1749-6632.2001.tb05827.x。[内政部] [公共医学] [谷歌学者]
-
Dantzer R、O'Connor JC、Freund GG、Johnson RW、Kelley KW。从炎症到疾病和抑郁:当免疫系统征服大脑时。《自然·神经科学评论》。2008;9:46–57. doi:10.1038/nrn2297。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Dickerson SS,Kemeny ME。急性应激源和皮质醇反应:实验室研究的理论整合和综合。精神斗牛。2004;130:355–391. doi:10.1037/0033-2909.130.3.355。[内政部] [公共医学] [谷歌学者]
-
艾森伯格NI,利伯曼医学博士。为什么拒绝伤害:生理和社会痛苦之间的神经认知重叠。趋势认知科学。2004;8:294–300. doi:10.1016/j.tics.2004.05.010。[内政部] [公共医学] [谷歌学者]
-
艾森伯格NI,利伯曼医学博士,威廉姆斯KD。拒绝是否有害:一项社会排斥的功能磁共振成像研究。科学。2003;302:290–292. doi:10.1126/science.1089134。[内政部] [公共医学] [谷歌学者]
-
艾森伯格NI、泰勒SE、盖博SL、希尔默特CJ、利伯曼MD。神经通路将社会支持与减弱的神经内分泌应激反应联系起来。神经影像学。2007;35:1601–1612. doi:10.1016/j.neuroimage.2007.01.038。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Fabes RA,Martin CL。情绪化的性别和年龄刻板印象。Pers-Soc心理公牛。1991;17:532–540.[谷歌学者]
-
第一个MB,Spitzer RL,Gibbon M,Williams JB。DSM-IV Axis I疾病结构化临床访谈,患者版,2.0版。纽约州精神病研究所;1996[谷歌学者]
-
Forman SD、Cohen JD、Fitzgerald M、Eddy WF、Mintun MA、Noll DC。改进功能性磁共振成像(fMRI)中显著激活的评估:使用簇大小阈值。麦格纳森医学,1995年;33:636–647. doi:10.1002/mrm.1910330508。[内政部] [公共医学] [谷歌学者]
-
弗里斯CD,弗里斯U。互动思维:生物学基础。科学。1999年;286:1692–1695. doi:10.1126/science.286.5445.1692。[内政部] [公共医学] [谷歌学者]
-
Frith U,Frith CD。心理化的发展和神经生理学。伦敦皇家学会哲学学报。2003;358:459–473. doi:10.1098/rstb.2002.1218。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Frith CD,Frith U。我们如何预测其他人会做什么。大脑研究。2006;1079:36–46. doi:10.1016/j.braines.2005.12.126。[内政部] [公共医学] [谷歌学者]
-
Frommberger UH、Bauer J、Haselbauer P、Fräulin A、Riemann D、Berger M.抑郁症和精神分裂症患者的白细胞介素-6(IL-6)血浆水平:急性状态和缓解后的比较。欧洲精神病学协会临床神经病学。1997;247:228–233. doi:10.1007/BF02900219。[内政部] [公共医学] [谷歌学者]
-
Gillespie CF、Nemeroff CB。皮质醇血症和抑郁症。《精神病医学》,2005年;67:S26–S28。doi:10.1097/01.psy.0000163456.22154.d2。[内政部] [公共医学] [谷歌学者]
-
Gold PW,Chrousos GP。抑郁症和非典型抑郁症中应激系统的组织及其失调:CRH/NE高与低状态。分子精神病学。2002;7:254–275. doi:10.1038/sj.mp.4001032。[内政部] [公共医学] [谷歌学者]
-
哈特BL。患病动物行为的生物学基础。《神经科学与生物行为评论》1988;12:123–137. doi:10.1016/s0149-7634(88)80004-6。[内政部] [公共医学] [谷歌学者]
-
Irwin MR、Wang M、Ribeiro D、Cho HJ、Olmstead R、Breen EC、Martinez-Maza O、Cole S.睡眠不足会激活细胞炎症信号。生物精神病学。2008;64:538–540. doi:10.1016/j.biopych.2008.05.004。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Kent S、BluthéRM、Kelley KW、Dantzer R.疾病行为作为药物开发的新目标。药理学趋势。1992;13:24–28. doi:10.1016/0165-6147(92)90012-u。[内政部] [公共医学] [谷歌学者]
-
Krabbe KS、Reichenberg A、Yirmiya R、Smed A、Pedersen BK、Bruunsgaard H.内毒素血症和人类神经心理功能低。大脑、行为和免疫。2005;19:453–460. doi:10.1016/j.bbi.2005.04.010。[内政部] [公共医学] [谷歌学者]
-
Kronfol Z.抑郁症患者的免疫失调:现有证据的批判性回顾。国际神经病药理学杂志。2002;5:333–343. doi:10.1017/S1461145702003024。[内政部] [公共医学] [谷歌学者]
-
Lane RD、Reiman EM、Ahern GL、Schwartz GE、Davidson RJ。快乐、悲伤和厌恶的神经解剖学关联。美国精神病学杂志。1997;154:926–933. doi:10.1176/ajp.154.7.926。[内政部] [公共医学] [谷歌学者]
-
Levine J,Barak Y,Chengappa KRN,Rapport A,Antelman SM,Barak V.急性抑郁症患者脑脊液可溶性白细胞介素2受体水平低。神经传输杂志。1999年;106:1011–1015. doi:10.1007/s007020050219。[内政部] [公共医学] [谷歌学者]
-
Lieberman MD、Jarcho JM、Berman S、Naliboff BD、Suyenobu BY、Mandelkern M、Mayer EA。安慰剂效应的神经相关性:干扰解释。神经影像学。2004;22:447–455. doi:10.1016/j.neuroimage.2004.01.037。[内政部] [公共医学] [谷歌学者]
-
MacKinnon DP、Lockwood CM、Hoffman JM、West SG、Sheets V.测试调解和其他干预变量效应的方法比较。心理方法。2002;7:83–104. doi:10.1037/1082-989x.7.1.83。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Maier SF,Watkins LR。心理学家用细胞因子:双向免疫-大脑沟通对理解行为、情绪和认知的影响。《心理学评论》1998;105:83–107. doi:10.1037/0033-295x.105.1.83。[内政部] [公共医学] [谷歌学者]
-
McNair DM、Lorr M、Droppleman LF。教育和工业测试服务。加利福尼亚州圣地亚哥:1971年。情绪状态简介手册。[谷歌学者]
-
Mitchell JP、Banaji MR、Macrae CN。内侧前额叶皮层中社会认知和自我参照思维之间的联系。《认知神经科学杂志》。2005;18:1306–1315. doi:10.1162/0898929055002418。[内政部] [公共医学] [谷歌学者]
-
Mullington JM、Haack M、Toth M、Serrador JM、Meier-Ewert HK。睡眠剥夺的心血管、炎症和代谢后果。心血管疾病进展。2009;51:294–302. doi:10.1016/j.pcad.2008.10.003。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Musselman DL、Miller AH、Porter MR、Manatunga A、Gao F、Penna S等。癌症抑郁症患者血浆白细胞介素-6浓度高于正常值:初步研究结果。美国精神病学杂志。2001;158:1252–1257. doi:10.1176/appi.ajp.158.8.1252。[内政部] [公共医学] [谷歌学者]
-
Nolen Hoeksema S.抑郁症的性别差异。心理科学的当前方向。2001;10:173–176.[谷歌学者]
-
Nolen-Hoeksema S,Girgus JS。青春期抑郁症中出现的性别差异。精神斗牛。1994;115:424–443. doi:10.1037/0033-2909.115.3.424。[内政部] [公共医学] [谷歌学者]
-
Ochsner KN,Gross JJ。情绪的认知控制。趋势认知科学。2005;9:242–249. doi:10.1016/j.tics.2005.03.010。[内政部] [公共医学] [谷歌学者]
-
Peyron R,Laurent B,Garcia Larrea L.大脑对疼痛反应的功能成像。综述和荟萃分析。神经生理学临床。2000;30:263–288. doi:10.1016/s0987-7053(00)00227-6。[内政部] [公共医学] [谷歌学者]
-
Phan KL、Wager TD、Taylor SF、Liberzon I.人类情绪的功能神经成像研究。CNS规范。2004;9:258–266. doi:10.1017/s1092852900009196。[内政部] [公共医学] [谷歌学者]
-
Phillips ML、Drevets WC、Rauch SL、Lane R.情绪感知的神经生物学I:正常情绪感知的神经学基础。生物精神病学。2003;54:504–514. doi:10.1016/s0006-3223(03)00168-9。[内政部] [公共医学] [谷歌学者]
-
Phillips ML、Young AW、Senior C、Brammer M、Andrew C、Calder AJ等。感知厌恶面部表情的特定神经基质。自然。1997;389:495–498. doi:10.1038/39051。[内政部] [公共医学] [谷歌学者]
-
Ploghaus A、Tracey I、Gati JS、Clare S、Menon RS、Matthews PM、Rawlins JNP。将疼痛与人类大脑中的预期分离。科学。1999年;284:1979–1981. doi:10.1126/science.284.5422.1979。[内政部] [公共医学] [谷歌学者]
-
Rainville P、Duncan GH、Price DD、Carrier B、Bushnell MD。疼痛影响编码于人类前扣带回,但不编码于体感皮层。科学。1997;277:968–971. doi:10.1126/science.277.5328.968。[内政部] [公共医学] [谷歌学者]
-
Raison CL、Capuron L、Miller AH。细胞因子唱布鲁斯:炎症和抑郁症的发病机制。Tr免疫学。2006;27:24–31. doi:10.1016/j.it.2005.11.006。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Raison CL、Demetrashvili M、Capuron L、Miller AH。干扰素α的神经精神不良反应:识别和管理。实践中的治疗。中枢神经系统药物。2005;19:105–123. doi:10.2165/00023210-200519020-00002。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Reichenberg A、Yirmiya R、Schuld A、Kraus T、Haack M、Morag A、Pollmächer T。人类细胞因子相关情绪和认知障碍。Arch Gen精神病学。2001;58:445–452. doi:10.1001/archpsyc.58.5445。[内政部] [公共医学] [谷歌学者]
-
Rivier C.白细胞介素-1β对大鼠下丘脑-垂体-肾上腺轴的刺激作用:年龄、性别和循环性类固醇的影响。内分泌杂志。1994;140:365–372. doi:10.1677/joe.01400365。[内政部] [公共医学] [谷歌学者]
-
Schiepers OJG、Wichers MC、Maes M.细胞因子与抑郁症。神经心理药理学和生物精神病学进展。2005;29:201–217。doi:10.1016/j.pnbp.2004.11.003。[内政部] [公共医学] [谷歌学者]
-
Stephens RCM、O'Malley CMN、Frumento RF、Mythen MG、Bennett-Guerrero E.低剂量内毒素引起健康志愿者炎症反应的变化。《内毒素研究杂志》。2005;11:204–209. doi:10.1179/096805105X58661。[内政部] [公共医学] [谷歌学者]
-
Suffredini AF,Hochstein HD,McMahon FG。人类静脉注射内毒素的剂量相关炎症效应:一批新的临床大肠杆菌O:113内毒素的评估。传染病杂志。1999年;179:1278–1282. doi:10.1086/314717。[内政部] [公共医学] [谷歌学者]
-
Suffredini AF,O'Grady NP.人类对内毒素的病理生理反应。收录人:Braude H、Opal SM、Vogel SN、Morrison DC,编辑。健康与疾病中的内毒素。第一。纽约:马塞尔·德克尔;1999年,第817-830页。[谷歌学者]
-
Tonelli LH,Holmes A,Postolache TT.鼻内免疫激发诱导大脑中的性依赖性抑郁行为和细胞因子表达。神经精神药理学。2008;33:1039–1048. doi:10.1038/sj.npp.1301488。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
Wager TD、Rilling JK、Smith EE、Sokolik A、Casey KL、Davidson RJ、Kosslyn SM、Rose RM、Cohen JD。安慰剂诱导的fMRI在疼痛预期和体验中的变化。科学。2004;303:1162–1167. doi:10.1126/science.1093065。[内政部] [公共医学] [谷歌学者]
-
Whitacre CC、Reingold SC、O'Looney PA。自体免疫的性别差异。科学。1999年;283:1277–1278. doi:10.1126/science.283.5406.1277。[内政部] [公共医学] [谷歌学者]
-
Williams KD、Cheung CKT、Choi W.网络排斥:在互联网上被忽视的影响。《社会心理学杂志》。2000;79:748–762. doi:10.1037//0022-3514.79.5.748。[内政部] [公共医学] [谷歌学者]
-
Wright CE,Strike PC,Brydon L,Steptoe A.急性炎症和负面情绪:细胞因子激活的介导。大脑、行为和免疫。2005;19:345–350. doi:10.1016/j.bbi.2004.10.003。[内政部] [公共医学] [谷歌学者]
-
Zautra AJ、Hamilton NA、Potter P、Smith B。类风湿关节炎应激与疾病活动之间关系的现场研究。Ann N Y科学院。1999年;876:397–412. doi:10.1111/j.1749-6632.1999.tb07664.x。[内政部] [公共医学] [谷歌学者]

