摘要
目标
低社会经济地位(SES)是与疾病发病率相关的最强大的社会因素之一,包括儿童期更严重的哮喘。然而,我们对解释这种联系的生物学过程的理解是有限的。这项研究测试了社会环境是否能“深入”改变哮喘儿童的基因组活动。
设计和参与者
对来自低或高社会经济地位家庭的医生诊断为哮喘的儿童进行两组设计。
结果
与SES背景较高的哮喘儿童相比,SES背景较低的哮喘儿童显示出调节炎症过程的基因过度表达,包括那些参与趋化因子活性、应激反应和创伤反应的基因。生物信息学分析表明,环AMP反应元件结合蛋白和核因子Y活性的降低以及核因子κB转录信号的增加介导了这些效应。已知这些途径调节免疫细胞中的儿茶酚胺和炎症信号。
结论
这项研究首次在一个确诊为哮喘的儿科患者样本中证明,更大的社会环境可以影响基因组水平的过程。具体而言,研究发现哮喘儿童中调节炎症和儿茶酚胺信号的基因转录控制途径因SES而异。由于这些途径是许多哮喘药物的主要靶点,这些研究结果表明,更大的社会环境可能会改变影响哮喘治疗效果的分子机制。
人们发现,更大的社会环境会影响个人一生的身体健康。在所有社会因素中,低社会经济地位(SES)与各种疾病的发病率和死亡率表现出最强烈、最一致的关联之一,1包括儿童哮喘。尤其是,与SES背景较高的儿童相比,SES背景较低的哮喘儿童更容易住院、急诊就诊和功能损害(如活动受限、卧床天数)。2–8
为了了解更大的社会环境“进入体内”对哮喘病理生理学的影响,研究人员调查了社会环境与炎症之间的关系。在健康成年人中,低SES与循环炎症标记物水平升高有关,如C反应蛋白和白细胞介素(IL)6。9 10在哮喘儿童中,低SES与嗜酸性粒细胞计数增加以及体外刺激产生的炎症细胞因子(如IL5和IL13)增加有关。11 12另一个与低SES应激相关的社会因素与哮喘患者对过敏原激发的炎症反应增强有关,13编码调节炎症的激素受体(包括糖皮质激素和β肾上腺素能受体)的基因表达减少。14
尽管SES与炎症之间的关系越来越被人们认识,但这种关系背后的分子机制仍不清楚。本研究使用基于体内基因组学的方法来确定哮喘儿童样本中与SES相关的T细胞基因表达的差异,并确定可能形成这些差异的候选信号转导途径。因为重点是了解与疾病进展相关的差异,所以我们以先前患有哮喘的儿童为样本。
基于先前将SES与炎症联系起来的研究,我们假设低SES与促炎症转录控制通路的改变有关。根据之前的数据,β肾上腺素能和糖皮质激素途径在社会因素和炎症之间的关系中,14我们假设,这些类型激素信号的转录介质将区分低SES背景和高SES背景儿童的T细胞基因表达谱。作为第二个目标,我们测试了与SES相关的社会因素,如认知解释,是否可以解释SES和全基因组转录谱之间的关联。我们的假设是,SES改变了孩子们解释社会世界的方式,而这些解释反过来又改变了全基因组转录谱显示的神经内分泌和炎症信号传递过程。
方法
有关更多详细信息,请参阅补充(仅限在线)。
参与者
参与者是从正在进行的一项针对9-18岁哮喘儿童的纵向研究中招募的。16名低社会经济地位儿童和15名高社会经济地位背景儿童为本研究提供了血样。低社会经济地位组和高社会经济地位组是根据那些在父母教育和家庭收入变量方面排名垫底和前15%的人来确定的。
措施
背景特征
家庭社会经济地位由父母的家庭年收入报告和父母教育年限来衡量。医疗信息包括哮喘药物的使用和哮喘严重程度。
压力的解释
《社交事件的认知评估和理解》(CAUSE)视频描述了与年龄相适应的模糊生活情景,例如被要求在课后与老师交谈。15法官对儿童的解释进行评分,数字越高,表示感知到的威胁越大。
基因表达阵列
通过免疫磁性阳性选择从单核细胞中分离CD2+细胞(Miltenyi Biotec;美国加利福尼亚州奥本市),并提取总RNA(RNAlater/RNeasy;美国加州巴伦西亚市齐亚根市)。如前所述,进行了全基因组转录谱分析。16–18差异表达基因是指那些在低SES和高SES背景的儿童中平均表达水平差异≥30%的基因(错误发现率为15%)。16使用GOstat生物信息学工具确定差异表达基因之间的功能共性(http://gostat.wehi.edu.au).19
转录调节分析
我们使用了转录元件监听系统(TELiS)的两个样本变体(网址:http://www.telis.ucla.edu)17如前所述,根据差异表达基因启动子内转录因子结合基序(TFBM)的流行情况分析差异基因表达数据。20
辅助分析使用了协方差分析的多元推广(ANCOVA)21以控制可能解释SES和基因表达谱之间关联的潜在混杂因素。协变量包括人口统计学变量(年龄、性别)、医学变量(哮喘严重程度、吸入性皮质类固醇药物的使用、β激动剂药物的使用)和压力解释(CAUSE视频反应)。为了证实自然杀伤细胞的流行不会影响结果,将编码CD16和CD56的mRNA的相对浓度作为协变量包括在内。
RT-PCR确认
使用TaqMan基因表达分析(Applied Biosystems,Foster City,California,USA),通过定量实时RT-PCR独立分析在微阵列分析中鉴定为差异表达的10个转录物,如在线所述补充.
临床变量
家长报告了过去两周内孩子的日间和夜间症状,以及他们的孩子在过去6个月内是否因哮喘而去了急诊室。
结果
样本的描述性信息如所示表1低SES组和高SES组在哮喘严重程度、吸入皮质类固醇的使用、β激动剂的使用、1s用力呼气量或特应性状态方面没有差异(所有p值均>0.15)。
表1。
|
低SES
|
高SES
|
|
% |
平均值 |
标准偏差 |
% |
平均值 |
标准偏差 |
| 年龄(年) |
|
12.8 |
3.5 |
|
14.9 |
1.9 |
| 性别(%男孩) |
63 |
|
|
85 |
|
|
| 药物(每种类型使用的百分比) |
|
|
|
|
|
|
| 吸入皮质类固醇 |
75 |
|
|
80 |
|
|
| β激动剂 |
75 |
|
|
87 |
|
|
| 特应性哮喘(%) |
75 |
|
|
93 |
|
|
| 哮喘严重程度(1-4级) |
|
2.8 |
1 |
|
2.5 |
0.5 |
| FEV公司1% |
|
94.9 |
12.6 |
|
99.8 |
17.6 |
差异基因表达
低SES组和高SES组之间有60个转录本差异表达,代表56个不同的命名人类基因。34例反映了低SES组中的过度表达,而26例反映了高SES组的过度表达(表2).
表2。
| Affymetrix探测器ID |
基因符号 |
折叠式变换 |
| 205048年_月_日 |
PSPH公司 |
2.49 |
| 202917年_月_日 |
S100A8型 |
1.80 |
| 221728_x天 |
XIST公司 |
1.65 |
| 220784秒 |
输出2 |
1.61 |
| 209480_吨 |
HLA-DQB1型 |
1.59 |
| 214146_秒_秒 |
PPBP公司 |
1.50 |
| 214218秒 |
XIST公司 |
1.49 |
| 209116_x_at |
乙型肝炎病毒 |
1.49 |
| 200782_at |
ANXA5公司 |
1.47 |
| 201743_日期 |
CD14号机组 |
1.45 |
| 211696_x_at |
乙型肝炎病毒 |
1.45 |
| 202912_at年 |
ADM公司 |
1.44 |
| 214414_x_at |
HBA2型 |
1.44 |
| 202651页 |
LPGAT1型 |
1.41 |
| 217941_s_地址 |
ERBB2IP公司 |
1.41 |
| 206390_x安 |
PF4系列 |
1.41 |
| 202503_天 |
诺基亚101 |
1.41 |
| 202602年_月_日 |
HTATSF1型 |
1.40 |
| 215223秒 |
SOD2标准 |
1.40 |
| 217022_s_地址 |
IGHA1///IGHA2 |
1.40 |
| 203535_吨 |
S100A9系列 |
1.39 |
| 202589_天 |
TYMS公司 |
1.38 |
| 221731_x _阿特 |
CSPG2公司 |
1.37 |
| 213831_at |
HLA-DQA1型 |
1.37 |
| 213376_安 |
ZBTB1公司 |
1.37 |
| 205863_安 |
S100A12型 |
1.36 |
| 209773_秒_秒 |
建议零售价2 |
1.32 |
| 204192_天 |
CD37细胞 |
1.32 |
| 217811_at |
选择 |
1.31 |
| 217414_x安 |
HBA1//HBA2型 |
1.31 |
| 212406秒 |
PCMTD2型 |
1.31 |
| 217232_x安 |
乙型肝炎病毒 |
1.31 |
| 220330_秒 |
SAMSN1公司 |
1.30 |
| 211745_x_at |
HBA1型 |
1.30 |
| 201464_x天 |
6月 |
0.78 |
| 2012年36月_日 |
基站2 |
0.77 |
| 221833_天 |
伦敦 |
0.77 |
| 205436秒_秒 |
H2AFX公司 |
0.77 |
| 214469_安 |
HIST1H2AE公司 |
0.77 |
| 205376_吨 |
INPP4B公司 |
0.77 |
| 214131_安 |
15亿日元 |
0.77 |
| 200664年_月_日 |
DNAJB1号机组 |
0.77 |
| 214805_吨 |
EIF4A1型 |
0.77 |
| 206385_秒_秒 |
银行3 |
0.76 |
| 2014年65月_日 |
6月 |
0.75 |
| 206700秒 |
SMCY公司 |
0.72 |
| 200799年_月 |
热休克蛋白A1 |
0.72 |
| 204141_吨 |
TUB2A公司 |
0.69 |
| 2016年93月_日 |
表皮生长因子受体1 |
0.69 |
| 2016年94月_日 |
表皮生长因子受体1 |
0.68 |
| 200800_秒_秒 |
热休克蛋白A1A///HSPA1B |
0.65 |
| 209189_安 |
FOS公司 |
0.65 |
| 219629_阿特 |
FAM118A型 |
0.64 |
| 213418_安 |
热休克蛋白A6 |
0.63 |
| 202768_天 |
FOSB公司 |
0.60 |
| 212671_s_at(电话) |
HLA-DQA1///HLA-DQA2//LOC650946 |
0.60 |
| 205000_吨 |
DDX3Y系列 |
0.55 |
| 202581页 |
HSPA1B型 |
0.50 |
| 201909_日期 |
RPS4Y1型 |
0.48 |
| 209728_安 |
HLA-DRB4型 |
0.46 |
在来自低SES背景的哮喘儿童中显示过度表达的基因包含以下主题:免疫反应和炎症(例如,S100A8型,S100A9系列,S100A12型,基因本体术语:GO:0006955,GO:000695,GO:0002376),趋化因子活性(例如,PPBP公司,PF4系列,GO:0008009,GO:0042379),压力响应(例如,SOD2标准,ANXA5公司,TYMS公司,GO:0006950)、伤口反应(例如,ADM公司,ANXA5公司,GO:0009611)和抗原处理和呈现(例如,HLA-DQA1型,HLA-DQB1型,GO:0042613,GO:042611,GO:0002504,GO:0019882)(所有p值均<0.05)。一些过度表达的基因与一个以上的GO注释相关,因此目前的结果不应被视为统计上独立的。然而,结果的聚合模式表明低SES背景哮喘儿童循环T淋巴细胞中炎症反应的相对激活。
相反,在SES背景较高的哮喘儿童中过度表达的基因具有与甲基化相关的功能主题(例如,FOS公司,基站2,GO:0043414,GO:0032259),蛋白质折叠(例如,热休克蛋白A1,热休克蛋白A6,GO:0006986,GO:0051789),细胞分化(例如,6月,GO:0035026),压力响应(例如,H2AFX公司,DNAJB1号机组,GO:0006950)和抗原处理和呈递(例如,HLA-DQA1型,HLA-DQA2型,GO:0042613,GO:0002504,GO:042611,GO:0019882)(所有p值均<0.05)。这些模式表明调节炎症过程和炎症反调节反应的基因相对激活。
验证性RT-PCR分析分析了在微阵列分析中鉴定为差异表达的10个转录物的表达。与全基因组差异表达分析中估计的<15%的错误发现率一致,所有测试的转录物在表达上都显示出统计上显著的变化(p<0.05)(表3).
表3。
| 基因 |
微阵列比率*
|
RT-PCR比率*
|
RT-PCR p值†
|
| 表皮生长因子受体1 |
0.68 |
0.52 |
0.0001 |
| FOS公司 |
0.65 |
0.59 |
0.0001 |
| FOSB公司 |
0.60 |
0.79 |
0.0487 |
| 6月 |
0.75 |
0.51 |
0.0001 |
| ADM公司 |
1.44 |
1.52 |
0.0114 |
| SOD2标准 |
1.40 |
1.25 |
0.0006 |
| S100A8型 |
1.80 |
1.65 |
0.0001 |
| S100A9系列 |
1.39 |
1.74 |
0.0001 |
| S100A12型 |
1.36 |
2.30 |
0.0001 |
| XIST公司 |
1.65 |
5.58 |
0.0001 |
转录控制途径
生物信息学分析用于识别差异表达基因启动子中的TFBMs。四种不同的激素或细胞因子信号通路始终与低SES相关(图1). 低SES组哮喘儿童中过度表达的基因启动子中,循环AMP反应元件结合蛋白(CREB)TFBM的发病率显著降低(平均下降55%,p=0.031)(表4). CREB蛋白是免疫细胞对儿茶酚胺转录反应的关键介质。22
图1。
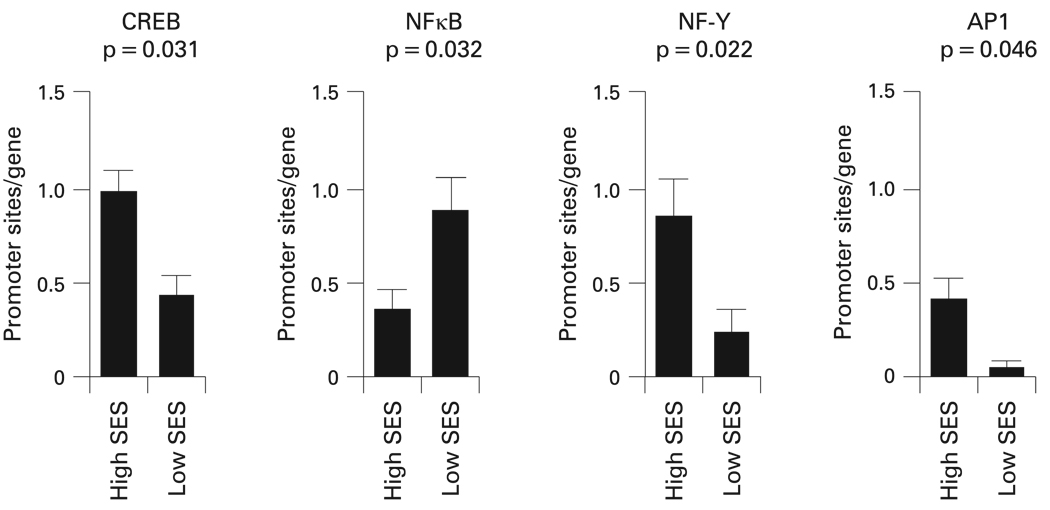
转录因子结合基序在低社会经济地位(SES)和高社会经济地位背景的哮喘儿童中过度表达的基因启动子中表现出差异。数据表示各组启动子中反应元件的平均(SEM)流行率。活化蛋白1;CREB,环AMP反应元件结合蛋白;核因子κB;NF-Y,核因子Y。
表4。
|
|
p值
|
| TFBM公司 |
比率*
|
单变量†
|
随机‡
|
敏感§
|
| V$CP2_01 |
2.33 |
0.0156 |
0.0114 |
0.0943 |
| V$TAL1BETAITF2_01 |
7.62 |
0.0166 |
0.0084 |
0.0922 |
| V$NFY_C |
0.28 |
0.0223 |
0.0212 |
0.0485 |
| V$CREB_Q4(沃尔沃) |
0.45 |
0.0306 |
0.0334 |
0.0056 |
| V$NFKB_Q6 |
2.54 |
0.0316 |
0.0232 |
0.0200 |
| V$SREBP1_01 |
4.50 |
0.0391 |
0.0316 |
0.0759 |
| V$COUP_01版 |
4.85 |
0.0434 |
0.0360 |
0.0926 |
| 伏$HSF2_01 |
0.71 |
0.0435 |
0.0460 |
0.0513 |
| V$AP1FJ_Q2 |
0.65 |
0.0464 |
0.0484 |
0.0005 |
| V$VJUN_01版 |
0.20 |
0.0477 |
0.0512 |
0.0342 |
第二项发现涉及低SES背景儿童中过度表达基因启动子中NF-Y TFBM的发病率下降(启动子TFBM发病率平均下降72%,p=0.022)。NF-Y被与CREB相同的cAMP/PKA信号通路激活,提供了通路激活的集中指示。
还发现来自低SES背景儿童T细胞的AP1(Fos/Jun)转录因子活性降低(AP1 p=0.046;VJUN p=0.048)。该途径活性的降低与观察到的FOS公司,FOSB公司和6月转录本,编码AP1转录因子家族的关键元素。
此外,低SES组中过度表达的基因中NFκB TFBM的患病率升高(启动子TFBM患病率平均增加2.54倍,p=0.032)。NFκB激活许多与炎症、伤口愈合和细胞应激反应有关的基因。
所有上述发现在参数变化的启动子长度和TFBM检测严格性的敏感性分析中仍有显著意义(表4). 在初步分析中,其他TFBM被确定为观察到的SES相关基因表达差异的潜在贡献者。然而,这些TFBM在参数敏感性分析中没有显示一致的统计显著性(p值>0.05)(表4). 为了评估观察到的差异可能偶然出现的可能性,我们从Affymetrix U133A高密度寡核苷酸阵列检测的基因群体中随机抽取了60个转录物的10000个样本,并计算了初始分析中确定的10个TFBM的差异流行程度。结果证实,在随机基因样本中观察到的CREB、NF-Y、AP1和NFκB的差异分布不太可能发生(p<0.05)(表4).
困惑者
在控制年龄和性别后,与CREB、NF-Y和NF-κB的关联模式仍具有统计学意义(所有p值均<0.045),只有一个例外,即SES和CREB控制年龄的关系(p=0.068)。在控制哮喘严重程度、吸入皮质类固醇和β受体激动剂使用后,相关性仍然显著(所有p值均≤.05)。与其他转录控制途径不同,SES和AP1活性之间的关联迹象通过控制几个混杂因素而变得不显著,包括年龄(p=0.52)、性别(p=0.15)和β激动剂的使用(p=0.11)。
控制认知解释将SES和CREB活动之间的关系降低到不显著(p=0.25)。SES与NFκB之间的关系也是如此(p=0.12),SES和NF-Y之间的关系也一样(p=0.077)。这些分析表明,低SES组的儿童更有可能在模糊的情况下感知到威胁,这种趋势可能会激活神经内分泌过程(通过cAMP/PKA通路影响CREB和NF-Y),最终影响炎症信号通路(如NFκB)。
最后,控制编码CD16和CD56的mRNA的流行率并没有显著改变CREB、NF-Y或NF-κB的结果(所有p值均<0.05),这表明结果不仅仅是各组中T细胞与NK细胞亚型的不同相对比例的函数。
临床结果
低SES组和高SES组的临床表现也有所不同,因此低SES组的儿童白天症状更严重(低SES平均值5.68(SD 5.24);高SES平均值1.50(SD 2.10);t=2.77,p=0.010),夜间症状加重(低SES平均值2.78(SD 4.19);高SES平均值0.36(SD 1.08);t=2.10,p=0.046),由于哮喘,过去6个月急诊科的使用率呈上升趋势(低SES 20%;高SES 0%;χ2=3.33,p=0.068)。
讨论
本研究提供了第一个在临床炎症性疾病背景下将较大社会环境(SES)与T细胞基因表达联系起来的实证证据。在哮喘儿童中,低SES背景的儿童表现出调节多种炎症过程的基因过度表达,包括趋化因子活性、应激反应和创伤反应。基于启动子的生物信息学分析确定了可能构建差异基因表达观察模式的转录控制途径,包括CREB、AP1和NF-Y减少,NFκB信号增加。低SES组的儿童也报告了较差的临床结果,如哮喘症状加重。虽然这项研究的横截面性质排除了关于因果方向的明确结论,但这些发现与更大的社会环境可以在基因组转录控制途径的水平上“深入”的假设是一致的。
低SES组儿童中过度表达的基因包括:趋化因子配体,如CXCL4和CXCL7,其招募和激活白细胞;23参与抗原处理和呈递(MHC II类蛋白复合物)的那些,这是哮喘炎症生物学的一个关键特征;那些与氧化应激有关的(SOD2标准)激活免疫细胞,延长炎症;以及与钙结合蛋白相关的(S100A8型,S100A9系列,S100A12型)对白细胞具有趋化作用,对内皮细胞具有促炎作用。24 25这些模式表明,低SES背景哮喘儿童的分子调控机制可能会加剧炎症并恶化临床结果。
高SES组儿童中过度表达的基因包括:编码热休克蛋白的基因(热休克蛋白A1,HSPA1B型,热休克蛋白A6,DNAJB1号机组)保护细胞免受活性氧等炎症分子的影响,并防止肺部炎症26 27; 那些参与细胞分化和增殖的细胞,如c-fos和c-jun,它们随着活性氧的暴露而增加28; 那些与细胞周期控制有关的(基站2)有助于抑制DNA损伤的影响29; 以及那些参与基因甲基化或沉默的人。请注意FOS公司,FOSB公司和6月基因产物还编码转录因子AP1家族的关键成分,在基于启动子的生物信息学分析中,高SES组的活性增加。总之,这些模式表明,在高SES背景的哮喘儿童中,炎症加剧可能会被旨在控制炎症损伤的细胞调节机制所抵消。
这项研究还确定了作为SES功能的基因表达差异模式的候选转录控制途径。生物信息学分析表明,低SES背景哮喘儿童CREB、AP1和NF-Y活性下调,NFκB介导的转录上调。CREB通过腺苷酸环化酶/cAMP/蛋白激酶A途径介导对β肾上腺素能受体信号的转录反应。22 30哮喘患者过敏原激发后腺苷酸环化酶活性受损。31cAMP通路的调节信号减弱可能导致T细胞活化增加,随后Th-2细胞因子表达增加。31CREB信号传导减少也可能降低用作哮喘治疗剂的支气管扩张剂的疗效。这些模式与患有慢性和急性生活应激的哮喘儿童淋巴细胞中发现的β肾上腺素能受体mRNA下降相一致。14
生物信息学分析还表明,低SES背景的哮喘儿童NF-Y介导的转录降低。与CREB一样,NF-Y被PKA磷酸化,因此可以作为β肾上腺素能信号通路的信号指示物。32事实上,这两种转录因子在低SES组中均表现出下调,这表明儿茶酚胺信号通路中存在多重平行缺陷。
生物信息学分析也表明低SES组哮喘儿童的NFκB信号上调。NFκB可激活多种炎症介质。一些数据表明,cAMP活性升高可以抑制NFκB活性33; 因此,NFκB的增加和CREB信号的减少可能代表了一种常见的调节性改变,这种改变使低SES背景的儿童的基因表达谱向更具炎症性的表型转变。
我们的发现与之前研究功能性免疫措施的研究一致。例如,低SES背景的哮喘儿童与高SES背景儿童相比,Th-2细胞因子的产生和嗜酸性粒细胞计数更多。12哮喘患者对过敏原激发的体内炎症反应和体外细胞因子的产生与更大的压力有关13 34并且易患过敏性疾病。35此外,压力与调节炎症的激素受体编码基因表达减少有关。14
统计分析表明,压力解释解释了CREB和NFκB活性指标中与SES相关的一些差异。这表明,为了使社会环境产生生物效应,可能必须首先以威胁的方式感知它。反过来,这些认知感知可能伴随着转录调控和炎症生物学的生物代价。我们注意到,SES与基因组模式之间可能存在许多其他的联系途径,这应该在未来的研究中进行测试。
目前尚不清楚在健康儿童中是否会发现类似的影响。然而,即使相似的作用很明显,这些生物学机制对那些已经存在炎症疾病的患者仍可能有不同的影响。尽管如此,测试SES对健康个体基因表达谱的影响对未来的研究也很重要。
局限性包括观察设计,排除了关于因果关系和方向性的结论。这是对涉及难以随机分配的社会因素(如社会经济地位)的人类工作的必要限制。其次,按流行病学标准来看,样本量较小,尽管在全基因组微阵列研究中并不罕见。小样本的一个担忧是功率不足;然而,几个转录控制途径的显著差异表明,这种效应的大小足以在这种大小的样本中检测到。第二个问题是研究结果是否稳健可靠。然而,请注意,本研究中使用的生物信息学方法利用了数千个基因的数据,形成了转录因子活性的聚合和更可靠的指标。第三,微阵列技术有时被批评为过于探索性。然而,在这项研究中,微阵列数据被用于测试先前研究中记录的特定激素信号通路导致SES炎症增加的先验假设。12 14此外,使用错误发现率分析是基因组文献中调整统计数据以解释多重比较的标准方法。然而,在其他样本中复制这些结果在未来的研究中很重要。最后,选择CD2抗原来分离T淋巴细胞可能已经允许被自然杀伤(NK)细胞污染。(没有使用CD3标记物,因为CD3的免疫磁性分离过程会激活T细胞。)然而,在控制NK细胞标记物mRNA的分析中,目前的结果没有显著改变。
目前的数据确定了哮喘儿童T淋巴细胞中低SES环境的一个独特的转录指纹,以及构成这些差异的候选转录控制通路。低SES背景的哮喘儿童显示与炎症、趋化因子活性、应激和创伤反应相关的基因过度表达,生物信息学显示CREB、AP1和NF-Y降低,NFκB信号增加。这项研究为哮喘提供了第一个临床证据,即更广泛的社会环境影响基因组水平的过程,特别是调节炎症和儿茶酚胺信号的转录控制途径。由于这些途径是许多哮喘药物的主要靶点,这些研究结果表明,更大的社会环境也可能影响哮喘治疗的疗效。最后,对压力的感知在解释SES如何在基因组水平上转化为改变方面发挥着重要作用。总的来说,这些发现为社会因素影响炎症疾病过程的机制提供了新的见解,并强调了未来研究哮喘的病理生理学如何由社会环境形成的必要性。
致谢
基金:这项研究得到了NIH拨款HL073975和加拿大卫生研究院的支持。
参考文献
-
1Starfield B,Robertson J,Riley AW.社会阶层梯度与儿童健康。Ambul儿科。2002;2:238–246. doi:10.1367/1539-4409(2002)002<0238:scgahi>2.0.co;2[内政部] [公共医学] [谷歌学者]
-
2Chen E,Matthews KA,Boyce WT。儿童健康的社会经济差异:这些关系如何以及为什么会随着年龄的增长而变化?精神斗牛。2002;128:295–329. doi:10.1037/0033-2909.128.2.295。[内政部] [公共医学] [谷歌学者]
-
三。Maziak W、von Mutius E、Keil U等。社区哮喘儿童卫生保健利用的预测因素。儿童过敏免疫学。2004;15:166–171. doi:10.1046/j.1399-3038.2003.00105.x。[内政部] [公共医学] [谷歌学者]
-
4Amre DK、Infante-Rivard C、Gautrin D等。哮喘儿童的社会经济状况和医疗服务利用。哮喘杂志。2002;39:625–631. doi:10.1081/jas-120014927。[内政部] [公共医学] [谷歌学者]
-
5Dales RE,Choi B,Chen Y,等。家庭收入对加拿大学龄儿童哮喘就诊的影响。胸部。2002;57:513–517. doi:10.1136/thorax.57.6.513。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
6Simon PA、Zeng ZW、Wold CM等。洛杉矶县儿童哮喘的患病率和相关发病率:种族/民族和收入的影响。哮喘杂志。2003;40:535–543. doi:10.1081/jas-120018788。[内政部] [公共医学] [谷歌学者]
-
7Miller JE。种族/民族和收入对儿童早期哮喘发病率和卫生保健使用的影响。美国公共卫生杂志。2000;90:428–430. doi:10.2105/ajph.90.3.428。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
8Halfon N,Newacheck PW。儿童哮喘与贫困:不同影响和卫生服务利用。儿科学。1993;91:56–61.[公共医学] [谷歌学者]
-
9.Panagiotakos DB、Pitsavos C、Manios Y等。ATTICA研究中健康个体与心血管疾病相关风险因素的社会经济状况。《欧洲心血管病防治杂志》,2005年;12:68–74.[公共医学] [谷歌学者]
-
10海明威H、希普利M、马伦MJ等。公务员炎症标记物和血管功能的社会和心理社会影响(白厅II研究)美国心脏病杂志。2003;92:984–987. doi:10.1016/s0002-9149(03)00985-8。[内政部] [公共医学] [谷歌学者]
-
11Chen E,Fisher EB,Jr,Bacharier LB,等。青少年哮喘患者的社会经济状况、压力和免疫标记物。《精神病医学》,2003年;65:984–992. doi:10.1097/01.psy.0000097340.54195.3c。[内政部] [公共医学] [谷歌学者]
-
12Chen E、Hanson MD、Paterson LQ等。儿童哮喘的社会经济状况和炎症过程:心理压力的作用。过敏临床免疫学杂志。2006;117:1014–1020. doi:10.1016/j.jaci.2006.01.036。[内政部] [公共医学] [谷歌学者]
-
13Liu LY,Coe CL,Swenson CA,等。学校考试增强了呼吸道炎症对抗原的挑战。Am J Respir Crit Care Med.2002;165:1062–1067. doi:10.1164/ajrccm.165.8.2109065。[内政部] [公共医学] [谷歌学者]
-
14Miller GE,Chen E.哮喘儿童的生活压力和编码糖皮质激素受体和β(2)-肾上腺素受体的基因表达减少。国家科学院院刊。2006;103:5496–5501. doi:10.1073/pnas.050631203。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
15Chen E,Matthews KA。社会事件认知评估和理解(CAUSE)视频的开发。健康心理学。2003;22:106–110. doi:10.1037//0278-6133.22.1106。[内政部] [公共医学] [谷歌学者]
-
16Cole SW、Galic Z、Zack JA。控制微阵列差异表达分析中的假阴性错误:PRIM方法。生物信息学。2003;19:1808–1816. doi:10.1093/bioinformatics/btg242。[内政部] [公共医学] [谷歌学者]
-
17Cole SW、Yan W、Galic Z等。转录因子活性的基于表达的监测:TELiS数据库。生物信息学。2005;21:803–810. doi:10.1093/bioinformatics/bti038。[内政部] [公共医学] [谷歌学者]
-
18Bolstad BM、Irizarry RA、Astrand M等。基于偏差和方差的高密度寡核苷酸阵列数据归一化方法的比较。生物信息学。2003;19:185–193. doi:10.1093/bioinformatics/19.2.185。[内政部] [公共医学] [谷歌学者]
-
19Beissbarth T,速度TP。GOstat:在一组基因中发现统计上表现过度的基因本体。生物信息学。2004;6:1464–1465. doi:10.1093/bioinformatics/bth088。[内政部] [公共医学] [谷歌学者]
-
20Wingender E、Dietze P、Karas H等。TRANSFAC:转录因子及其DNA结合位点数据库。1996年《核酸研究》;24:238–241. doi:10.1093/nar/24.238。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
21米勒RG。超越方差分析:应用统计学基础。纽约州纽约市:Wiley;1986[谷歌学者]
-
22Kammer GM。腺苷酸环化酶–cAMP-蛋白激酶A途径和免疫反应的调节。今日免疫学。1988;9:222–229. doi:10.1016/0167-5699(88)91220-0。[内政部] [公共医学] [谷歌学者]
-
23Lukacs NW,Miller AL,Hogaboam CM。哮喘中的趋化因子受体:寻找正确的免疫靶点。免疫学杂志。2003;171:11–15. doi:10.4049/jimmunol.171.1.11。[内政部] [公共医学] [谷歌学者]
-
24Lackmann M、Rajasekariah P、Lismaa SE等。前炎症S100蛋白CP-10趋化域的鉴定。免疫学杂志。1993;150:2981–2991.[公共医学] [谷歌学者]
-
25.Hofmann MA、Drury S、Fu CF等。RAGE介导一种新的促炎轴:S100/钙粒蛋白多肽的中央细胞表面受体。单元格。1999;97:889–901. doi:10.1016/s0092-8674(00)80801-6。[内政部] [公共医学] [谷歌学者]
-
26Jacquiersarlin MR,Fuller K,Dinhxuan AT,等。HSP70在炎症中的保护作用。体验。1994;50:1031–1038. doi:10.1007/BF01923458。[内政部] [公共医学] [谷歌学者]
-
27Villar J、Edelson JD、Post M等。热应激蛋白的诱导与急性肺损伤动物模型死亡率的降低相关。美国Rev Respir Dis。1993;147:177–181. doi:10.1164/ajrccm/147.1.177。[内政部] [公共医学] [谷歌学者]
-
28Janssen YMW、Matalon S、Mossman BT。暴露于ROS或RNS的肺上皮细胞中c-fos、c-jun和凋亡的差异诱导。美国生理学杂志肺细胞分子生理学。1997;17:L789–L796。doi:10.1152/ajplung.1997.273.L789。[内政部] [公共医学] [谷歌学者]
-
29Rouault JP、Falette N、Guehenneux F等。BTG2的鉴定,BTG2是DNA损伤细胞反应途径的一种抗增殖p53依赖性成分。自然遗传学。1996;14:482–486. doi:10.1038/ng1296-482。[内政部] [公共医学] [谷歌学者]
-
30.Sanders VM、Straub RH、去甲肾上腺素、β-肾上腺素能受体和免疫。大脑行为免疫。2002;16:290–332. doi:10.1006/brbi.2001.0639。[内政部] [公共医学] [谷歌学者]
-
31Borger P,Postma DS,Vellenga E等。通过环腺苷酸依赖性信号通路调节哮喘相关T细胞因子。《临床实验过敏》2000;30:920–926. doi:10.1046/j.1365-2222.2000.00794.x。[内政部] [公共医学] [谷歌学者]
-
32Kapatos G,Vunnava P,Wu Y.大鼠GTP环水解酶I近端启动子RNA聚合酶II、C/EBPbeta和NF-Y依赖于蛋白激酶A的募集,组蛋白乙酰化无变化。神经化学杂志。2007;101:1119–1133. doi:10.1111/j.1471-4159.2007.04486.x。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
33Elenkov IJ、Wilder RL、Chrousos GP等。交感神经——两个超级系统之间的整合接口:大脑和免疫系统。药理学修订版2000;52:595–638.[公共医学] [谷歌学者]
-
34Kang D,Coe C,McCarthy DO,等。整个学年哮喘和健康青少年受刺激血淋巴细胞的细胞因子谱。干扰素细胞因子研究杂志1997;17:481–487. doi:10.1089/jir.1997.17.481。[内政部] [公共医学] [谷歌学者]
-
35.Wright RJ、Finn P、Contreras JP等。慢性照顾者压力和IgE表达、过敏原诱导的增殖和易患特应性反应的出生队列中的细胞因子特征。过敏临床免疫学杂志。2004;113:1051–1057. doi:10.1016/j.jaci.2004.03.032。[内政部] [公共医学] [谷歌学者]
关联数据
本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。

