摘要
理论基础在实验模型中,肺纤维化依赖于转化生长因子(TGF)-β信号传导。TGF-β与其前肽在一种潜在的复合物中分泌,TGF-α激活剂从该复合物中释放TGF-γ。由于整合素αvβ6是肺中主要的TGF-β激活物,抑制αvβ6-介导的TGF--β激活是治疗肺纤维化的合理策略。
目标:通过遗传和药理学方法确定小鼠辐射诱导的肺纤维化是否依赖于αvβ6。
方法:野生型小鼠,αvβ6缺乏(Itgb6号机组−/−)小鼠和杂合子小鼠Tgfb1型消除整合素介导激活的突变(Tgfb1型+/RGE公司)暴露于14Gy胸部辐射。用抗αvβ6单克隆抗体或可溶性TGF-β受体融合蛋白治疗一些小鼠。免疫组化检测αvβ6的表达。在照射后20–32周评估纤维化、炎症和基因表达模式。
测量和主要结果照射后18周,即纤维化开始前,肺泡上皮内β6整合素表达增加。Itgb6号机组−/−小鼠完全避免了纤维化,但不能避免晚期辐射诱导的死亡。抗αvβ6治疗(1–10 mg/kg/wk)可预防纤维化,但只有较高剂量(6–10 mg/kg wk)引起的肺部炎症与Itgb6号机组−/−老鼠。Tgfb1型-单倍体小鼠也能防止纤维化。
结论:辐射诱导的肺纤维化需要αvβ6介导的TGF-β激活。结合之前的数据,我们的结果证明了在不同的纤维化模型中对αvβ6的稳健需求。抑制αvβ6介导的TGF-β活化是一种很有前景的抗纤维化治疗新方法。
概览评论
关于该主题的科学知识
转化生长因子(TGF)-β是一种促纤维化细胞因子,其潜在形式被肺中的整合素αvβ6激活。已开发出一种单克隆抗体,能有效且特异性地抑制αvβ6。
本研究为该领域增加了什么
抑制αvβ6介导的TGF-β活化可预防小鼠辐射诱导的肺纤维化,且抗纤维化作用的剂量低于因TGF-α免疫调节作用丧失而导致肺部炎症的剂量。
肺纤维化发生于损伤后基质重塑紊乱(1). 细胞因子,转化生长因子(TGF)-β,影响参与该反应的多个过程(如基质产生、上皮细胞凋亡、上皮-间充质转化、成纤维细胞表型和蛋白酶活性),其活性是几种实验性纤维化所必需的(2,三). 因此,抑制TGF-β信号传导作为一种抗纤维化治疗正在被深入研究。
在三种转化生长因子-β亚型中,TGF-β1可能在纤维化中起着最重要的作用。TGF-β基因编码一个C端TGF-?序列和一个称为潜伏相关肽(LAP)的N端前体蛋白。TGF-β和LAP以一种复合物的形式分泌,其中TGF-α是潜伏的(即不能与TGF-γ受体结合)。LAP还与潜伏TGF-β结合蛋白家族的蛋白质相互作用,该家族将潜伏TGF-β锚定于细胞外基质(4). 从LAP(TGF-β激活)中释放TGF-(5).
假定的TGF-β激活物包括降解LAP的蛋白酶(6,7),血小板反应蛋白-1(8,9),活性氧物种(10)以及整合素αvβ6和αvβ8,它们与位于TGF-β1–LAP和TGF-α3–LAP C末端附近的氨基酸序列精氨酸-甘氨酸-天冬氨酸(RGD)相互作用(11–13). 整合素介导的转化生长因子-β1激活是转化生长因子β1在血管生成、免疫耐受和朗格汉斯细胞形成中的作用所必需的(14).
在肺中,αvβ6在正常上皮中低水平表达,并在损伤后上调。β6亚基基因缺失突变导致小鼠缺乏αvβ6(Itgb6号机组−/−)在大多数方面都是健康的,但有与TGF-β1阴性小鼠相似的肺部炎症,以及由于肺泡巨噬细胞(AM)失去TGF-α介导的基质金属蛋白酶-12表达抑制而延迟发作的肺气肿(15,16). 气管内给予博莱霉素会导致急性肺损伤和快速纤维化反应;该模型中早期毛细血管渗漏和纤维化均依赖于TGF-β信号转导和αvβ6的表达(13,17). 这些观察结果增加了抑制αvβ6可能在不消除所有TGF-β信号的情况下预防肺纤维化的可能性,如肾脏模型所示(18).
我们研究了αvβ6在辐射诱导肺纤维化(RILF)模型中的作用。与博莱霉素诱导的纤维化相比,RILF发病较晚,发生在辐射损伤后20周或更长时间,并且与急性损伤或严重炎症不同步。我们选择该模型来测试αvβ6是否参与机械性不同的肺纤维化模型,并测试在纤维化反应期间而不是在初始肺损伤期间给予治疗的效果。我们的结果表明,αvβ6在不同形式的肺纤维化中发挥着强大的作用,是一个很有希望的治疗靶点。
方法
肺部照射
小鼠为C57BL/6J雌性。突变小鼠Tgfb1型如前所述,在TGF-β1–LAP的整合素结合位点产生了编码精氨酸-甘氨酸-谷氨酸(RGE)而不是精氨酸-甘氨酸-天冬氨酸的外显子5(14)在C57BL/6J背景上回交10代;Tgfb1型+/+和Tgfb1型+/RGE公司同窝婴儿被用于照射。Itgb6号机组−/−回交到C57BL/6J背景的小鼠来自Dean Sheppard(加州大学旧金山分校)。用Avertin麻醉小鼠(8-10周龄),并将其暴露于14Gy的胸部辐射下60共同来源。纽约大学医学院动物护理委员会批准了所有程序,这些程序符合美国国立卫生研究院(马里兰州贝塞斯达)的指南。
肺、血液和支气管肺泡灌洗液程序
用800μl 10%福尔马林对死亡或垂死小鼠的肺进行充气。在抗αvβ6单克隆抗体(mAb)实验中,用700μl磷酸盐缓冲盐水(PBS)冲洗肺部两次。结扎右主干支气管,右肺在液体N中冷冻2左肺用400μl 10%福尔马林充气。左肺横切成5μm切片,用Masson三色染色。用等分的支气管肺泡灌洗液(BAL)进行细胞计数和胞浆计数,其余的冰冻于N液体中2如前所述,用抗β6嵌合单克隆抗体2A1免疫组织化学检测β6蛋白(18). 如前所述计算纤维化面积百分比(%FA)(19)使用ImageJ软件(美国国立卫生研究院)。对于Tgfb1型+/+和Tgfb1型+/RGE公司小鼠,用每只小鼠的左肺和右肺测量%FA。羟脯氨酸含量的测量如前所述(20).
抗体治疗
以前已经描述过抑制性抗αvβ6单克隆抗体6.3G9、同型对照抗体1E6和重组可溶性TGF-β受体II-Fc融合蛋白(rsTGF-?RII-Fc)(18,21). 抗体每周注射一次,可以是腹膜内注射(第一次实验),也可以是皮下注射。注射体积为200μl。
右:左心室质量比测量
将照射后28至32周死亡的小鼠心脏与照射后32周死亡或7只未照射C57BL/6J小鼠的心脏进行比较。从左心室和室间隔(LV)上解剖右心室游离壁(RV),并对各个部分进行称重。
RNA分离
根据制造商的方案,使用Qiazol试剂(加利福尼亚州巴伦西亚Qiagen)从储存在−80°C下的肺部制备总RNA。在生物分析仪2100(安捷伦,圣克拉拉,加利福尼亚州)上通过毛细管电泳验证RNA质量。
Taqman引物、探针和寡核苷酸标准模板的设计
使用Primer Express 2.0.0版(加利福尼亚州福斯特市应用生物系统公司),根据Affymetrix(加州圣克拉拉)的共识序列设计寡核苷酸引物和Taqman小沟槽结合物(MGB)探针。Taqman MGB探针采用5′荧光报告染料、6-羧基荧光素(FAM)和3′MGB/非荧光猝灭剂(MGBNF)设计。通过在扩增子的5′端和3′端添加10 bp的基因特异性序列来设计寡核苷酸标准模板。反相高效液相色谱-纯化引物和寡核苷酸标准模板购自Biosearch Technologies Inc.(Novato,CA)。在Biogen Idec合成了小鼠甘油醛-3-磷酸脱氢酶的HPLC纯化引物和探针(序列CATGGCCTTCGTGTTCCTA、GCGGCACGTCAGATCC和6FAM-CCCCAATGTCGTC)。
塔克曼热循环
样品和标准品的四重聚合酶链反应在7900HT(Applied Biosystems,Inc.)热循环器中在以下条件下循环:50℃2分钟,95℃10分钟,以及40个95℃循环15秒和60℃循环60秒。对于每个反应孔,每7秒收集一次荧光发射。通过使用序列检测软件(Applied Biosystems,Inc.)与寡核苷酸标准曲线进行比较,确定每个样本的相对转录物数量
微阵列程序
RNA样品的质量(每个实验组至少5个)通过毛细管电泳在生物分析仪2000(安捷伦)上进行验证。从单个RNA样品中制备杂交探针,并在单独的小鼠基因组430 2.0寡核苷酸阵列(Affymetrix)上进行分析。使用制造商的协议进行杂交探针合成、杂交和微阵列扫描。阵列扫描被转换为Affymetrix。使用GC含量调整稳健微阵列平均值方法对CEL文件和结果数据集(代表整个实验的一组.CEL文件)进行标准化。使用GeneSpring(Agilent)软件进行统计和聚类分析。我们采用了两步t吨首先,在比较辐照过的PBS处理小鼠和未辐照过的老年对照组时,通过试验筛选确定信号强度发生改变的问题(P(P)<0.001),其次,当比较经辐照的PBS处理小鼠和经1 mg/kg 6.3G9处理的辐照小鼠时(P(P)< 0.05; PBS治疗的辐射组n=3,其他测试组n=5-8)。使用Ingenuity Pathways Analysis数据库(Ingenuiity,Redwood City,CA)对选定的基因列表进行功能注释。
统计分析
通过Mann-Whitney U检验评估不同治疗组的平均%FA之间差异的显著性。使用Student’s评估平均羟脯氨酸浓度或平均RV:LV比率之间的差异t吨测试。测量值的平均值用误差条显示,误差条表示SEM。使用单向方差分析对载体对照组和/或同型对照组以及不同剂量的供试品之间的BAL液中蛋白质水平进行统计比较。当确定具有统计学意义的差异时,通过Dunnet的多重比较试验(显著性定义为P(P)< 0.05).
结果
肺泡上皮αvβ6表达增加是放射损伤后的晚期事件
αvβ6整合素通常在肺上皮中低水平表达,但可通过损伤和炎症上调。在照射后2周,我们用识别β6亚单位的单克隆抗体通过免疫组织化学染色评估β6整合素的表达。β6的表达在照射后18~20周整个肺泡上皮细胞从基线水平的低水平急剧增加之前没有改变(图1A和数据未显示)。照射后20–22周,纤维化区域首先变得明显(数据未显示)。照射后24周,β6在非纤维化区域的整个肺泡上皮和位于胸膜下的纤维化区域的上皮细胞中表达显著;照射后27周及以后,纤维化区域的上皮细胞中持续存在强烈阳性的β6表达,但在非纤维化区域通常不那么显著(图1B和数据未显示)。
图1。
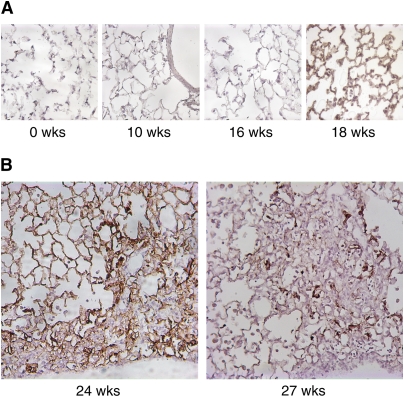
αvβ6在照射后肺部的表达。(A类)用β6整合素亚基特异性抗体(ch2A1)对辐照野生型小鼠的肺部进行免疫染色。照射后18周,αvβ6表达增加。(B类)αvβ6在纤维化区域高表达。照射后24周,非纤维化区域的免疫染色也很强烈。在照射后27周,非纤维化区域的免疫染色不太明显,但在纤维化病变内的细胞中仍很强烈。
缺乏αvβ6的小鼠不会发展成RILF
αvβ6表达与RILF病变之间的密切时空相关性支持了αvβ6-介导的TGF-β活化参与纤维化过程的观点。为了确定αvβ6是否对RILF的发展是必要的,我们比较了辐照后的纤维化反应Itgb6号机组+/+和Itgb6号机组−/−老鼠。在Itgb6号机组+/+肺(图2A),纤维化区域界限清楚,位于胸膜下。我们在23例患者中的21例切片中观察到纤维化区域Itgb6号机组+/+小鼠在照射后24-28周(平均26.0周)死亡,但在17个肺切片中均未发现纤维化区域Itgb6号机组−/−小鼠放疗后26-28周(平均放疗后27.1周),这一差异具有统计学意义(P(P)=0.001,Fisher精确检验)。来自的截面的%FAItgb6号机组+/+小鼠(照射后27周)为17±3%。其他章节分析Itgb6号机组−/−小鼠没有发现纤维化区域,证实了Haston及其同事的报告,即分析每只小鼠的单个肺切片足以评估纤维化(19). 我们通过测量来自Itgb6号机组+/+和Itgb6号机组−/−小鼠照射后27周(图2B).
图2。
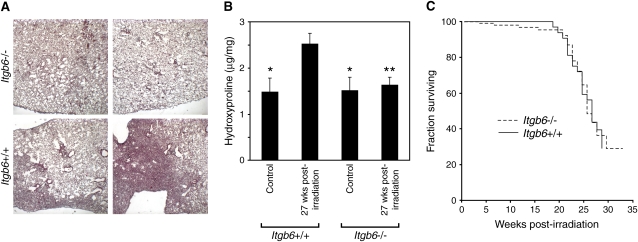
缺乏αvβ6整合素的小鼠不会发生辐射诱导的肺纤维化。(A类)代表性肺部Itgb6号机组+/+和Itgb6号机组−/−小鼠(照射后27周)用Masson三色染色。(B类)肺羟脯氨酸含量Itgb6号机组+/+和Itgb6号机组−/−小鼠(照射后27周)。辐照后羟脯氨酸含量Itgb6号机组+/+肺明显大于未受辐射的肺Itgb6号机组+/+肺部以及照射和未照射的Itgb6号机组−/−肺部(*P(P)<0.03 vs.辐照Itgb6号机组+/+; **P(P)<0.02 vs.辐照Itgb6号机组+/+; 每组n=5-6)。误差线表示1个SEM(C类)Kaplan-Meier曲线Itgb6号机组+/+和Itgb6号机组−/−接受14Gy胸部放射治疗的小鼠。
胸部照射14Gy后,直到照射后18-20周死亡率可忽略不计,照射后26周死亡率达到50%(图2C). 存活率无显著差异Itgb6号机组+/+和Itgb6号机组−/−老鼠。这些生存曲线与先前对雌性C57BL/6小鼠的结果相似(19).
抑制性抗αvβ6单克隆抗体和TGF-β拮抗剂预防RILF
在辐射诱导纤维化晚期阻断TGF-β信号传导的作用尚未明确。为了解决这个问题,我们在照射后16周,即αvβ6上调之前,开始使用抗αvβ6mAb 6.3G9进行治疗,抗αvα6mAb是一种能够阻止αvβ6-介导的TGF-β活化的特异性有效抑制剂(21). 此外,我们用rsTGF-βRII-Fc治疗小鼠,其抑制活性TGF-(18). 对照射后20周以上死亡的小鼠和照射后26周在研究终点死亡的小鼠进行了纤维化评估。
用PBS、对照IgG或0.3mg/kg/周治疗的小鼠6.3G9出现类似程度的纤维化(图3A). 相比之下,用5 mg/kg/周rsTGF-βRII-Fc或1或10 mg/kg/周6.3G9治疗的小鼠,其组织学定义的纤维化区域(以%FA计)显著减少。治疗还将组织学分析中任何程度的纤维化小鼠的比例从对照IgG和PBS组的17/24和10/14分别降低到14/24,0.3、1.0和10.0 mg/kg/周6.3G9和rsTGF-βRII-Fc组分别为3/23、3/21和10/27(P(P)= 0.55,P(P)< 0.0001,P(P)=0.0002,以及P(P)分别=0.025,与IgG对照组相比[费希尔精确试验])。
图3。
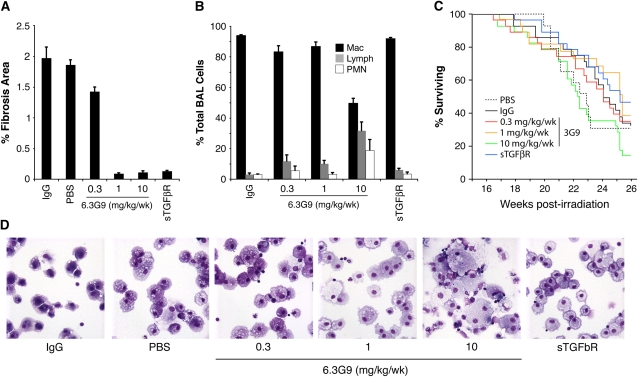
抗αvβ6单克隆抗体(mAb)6.3G9对肺辐射后纤维化、肺部炎症和生存的影响。向小鼠胸部照射14 Gy。照射后16周开始每周腹腔注射对照IgG、对照磷酸盐缓冲液(PBS)、抗αvβ6单克隆抗体(6.3G9)或重组可溶性转化生长因子-β受体II–Fc融合蛋白(rsTGF-βRII–Fc)(每组14–27例)。显示了6.3G9的剂量;rsTGF-βRII-Fc剂量为5 mg/kg/周。(A类)汇总辐照后20至26周死亡小鼠和辐照后26周死亡的小鼠的结果。与IgG对照组相比,用1或10 mg/kg/周6.3G9或rsTGF-βRII-Fc治疗的小鼠的纤维化面积百分比(%FA)显著降低(P(P)与对照IgG相比<0.001)。(B类)照射后26周处死的小鼠支气管肺泡灌洗液中细胞的差异计数。(C类)在治疗阶段,不同治疗组的Kaplan-Meier曲线。在综合分析中,生存曲线没有显著差异(P(P)=0.088(通过log-rank测试)。(D类)BAL细胞的代表性胞浆。巨噬细胞增大和其他炎症细胞类型的增加在用10 mg/kg/周6.3G9处理的小鼠的BAL液中是明显的。误差线表示1个SEM。
为了评估不同治疗组的炎症反应,我们对存活至死亡的小鼠的BAL液进行了细胞计数。各组间细胞总数无显著差异(数据未显示)。然而,差异细胞计数显示,低剂量6.3G9(0.3和1 mg/kg/wk)和rsTGF-βRII-Fc的淋巴细胞百分比无显著增加,而10 mg/kg/ww的淋巴细胞和中性粒细胞显著增加(图3B). 来自高剂量6.3G9小鼠的胞浆含有扩大的泡沫状AM,其外观类似于从肺分离的AMItgb6号机组−/−老鼠(图3D).
我们比较了各组小鼠的Kaplan-Meier生存曲线,发现23-25周时存活率达到50%(图3C)与初始实验中的26周相比(图2C). 各组间无显著差异(P(P)所有组的log-rank检验=0.088),但10 mg/kg/周6.3G9组的存活率明显下降。
这些结果证实了小鼠RILF是TGF-β依赖性的,并且在纤维化阶段之前和期间抑制TGF-αvβ6可以阻止RILF。结果还表明,不同6.3G9剂量的影响存在潜在的重要差异。虽然每周1毫克/千克和10毫克/千克剂量的6.3G9都可以预防RILF,但只有较高剂量的药物与BAL淋巴细胞和中性粒细胞的增加有关。未经治疗的患者肺淋巴细胞(三到四倍)也会增加,但中性粒细胞不会增加Itgb6号机组−/−老鼠(16).
RILF中6.3G9的剂量-反应评估
我们进行了第二次αvβ6抑制实验,以检查额外剂量6.3G9的影响,并重新检查6.3G9对生存的影响。我们对照射后较长时间(28-32周)的小鼠进行了检查,以确定6.3G9治疗是否延迟而不是预防RILF。照射16周后再次开始注射,每组41–44只小鼠接受照射。
我们测量了20周后死亡或28或32周结束时死亡的所有小鼠的%FA。对照组小鼠的典型病变如所示图4A与用对照IgG治疗的小鼠相比,任何剂量的6.3G9治疗组的FA百分比显著降低(图4B). 最低剂量6.3G9(1 mg/kg/wk)的小鼠的%FA为对照组的18%,而较高剂量(3–10 mg/kg/ww)的小鼠只有对照组的3–5%。与之前一样,比较有无纤维化的样本数量时,差异也具有统计学意义。在对照IgG处理的小鼠中,35/42有组织纤维化;对于每周服用1、3、6或10毫克/千克6.3G9的小鼠,只有25/42、10/42、8/42和9/41小鼠出现组织学上明显的纤维化(P(P)=0.029,1 mg/kg/wk 6.3G9和P(P)与对照IgG组相比,其他组<0.0001(Fisher精确试验)。
图4。
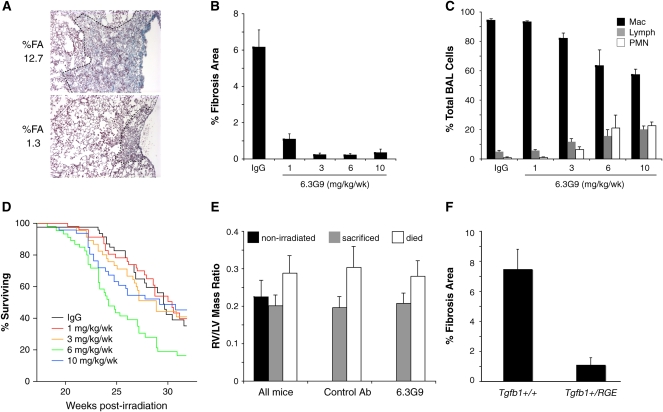
抗αvβ6单克隆抗体(mAb)附加治疗剂量分析。照射后16周开始每周皮下注射抗αvβ6单克隆抗体(6.3G9)或对照IgG(每组14–27)。(A类)两个对照抗体治疗的小鼠的典型纤维化病变。这个虚线表示病变的程度。整个肺切片的纤维化面积百分比(%FA)显示为左边. (B类)汇总辐照后20至32周内死亡的小鼠和28或32周后死亡的小鼠的结果。所有剂量的6.3G9都会显著降低FA的百分比。(C类)照射后28周或32周处死的小鼠BAL液中细胞的差异计数。(D类)不同治疗组在治疗阶段的Kaplan-Meier曲线。在综合分析中,存活曲线存在显著差异(P(P)<0.005(通过log-rank测试)。(E类)右心室(RV)的比较:照射后28至32周死亡的小鼠与存活至32周时死亡的小鼠的左心室(LV)质量比。所有小鼠的结果显示在左边使用来自未辐照控件的数据。打开正确的是用对照IgG或不同剂量的6.3G9处理的小鼠的数据。在所有显示的分组中,与存活小鼠相比,死亡小鼠的RV:LV比率显著增加(P(P)<0.0007)或未辐照(P(P)<0.02)小鼠。(F类)Tgfb1型+/+和Tgfb1型+/RGE公司26周后照射小鼠并处死(n=8和12)。杂合子小鼠的%FA显著降低Tgfb1型突变(P(P)< 0.001).误差线表示1个SEM。
我们在照射后28周杀死了大约等量的小鼠,并收集BAL液进行细胞计数(每组19–22只小鼠,但6 mg/kg/wk 6.3G9组的n=7只除外)。与之前一样,我们观察到淋巴细胞和中性粒细胞的百分比呈剂量依赖性增加,仅在每周6和10毫克/千克的剂量下才显著增加(图4C).
各治疗组的生存曲线如所示图4D:照射后27–31周达到50%的存活率,但6.3G9组的存活率为6 mg/kg/周,其中24周达到50%。存活曲线的差异具有统计学意义(P(P)<0.005,对所有组进行log-rank检验)。
先前的研究表明,右心室肥大(RVH)和肺灌注丧失是肺照射的晚期后遗症(22,23)这可能是呼吸功能不全和死亡的原因。为了在我们的实验中评估RVH和致死率的相关性,我们测量了三组小鼠的RV:LV质量比:照射后28至32周死亡的受照小鼠;存活32周后处死的小鼠;和对照未受辐射的小鼠。与其他两组小鼠相比,无论治疗状态如何,死亡小鼠的RV:LV比率都显著增加(图4E).
剂量滴定实验表明,与控制肺部炎症相比,需要更多的αvβ6激活的TGF-β来引起纤维化。为了以不同的方式验证这个假设,我们使用了敲除基因突变为Tgfb1型消除TGF-β1–LAP的整合素结合位点(14). 这种突变的小鼠纯合子(Tgfb1型RGE/RGE公司)产生正常水平的潜在TGF-β1蛋白,但其表型与Tgfb公司−/−老鼠。杂合小鼠具有较少的整合素激活的潜在TGF-β1,但没有炎症表型。我们辐照过Tgfb1型+/+和Tgfb1型+/RGE公司受照26周后的室友和评估纤维化(图4F). %固定资产Tgfb1型-单倍型小鼠明显少于野生型(P(P)< 0.001).
BAL液蛋白浓度的多元分析
为了进一步表征RILF模型中抗αvβ6治疗的效果,我们通过多分析蛋白谱测定了BAL液中60种选定蛋白的水平。第二次αvβ6抑制实验的小鼠在照射后28或32周被处死,用于这些测量。最引人注目的发现是,与低剂量6.3G9或对照抗体处理的受照小鼠BAL液中的水平相比,受照小鼠的BAL液(每周6.3G9剂量为6和/或10 mg/kg)中的许多蛋白质(主要是炎症介质)的水平显著升高。四种代表性蛋白质的数据如所示图5B和5C在用6.3G9或对照单克隆抗体治疗4周的未受照射小鼠中观察到质量类似的剂量-反应模式(图5A). 与对照组相比,6.3G9组6和10 mg/kg/周组的其他蛋白显著升高,但在6.3G9 1和3 mg/kg/星期组中没有升高,包括基质金属蛋白酶-9、抑癌素M、血管内皮生长因子和肿瘤坏死因子-α,以及趋化因子粒细胞趋化蛋白(GCP)-2,干扰素诱导蛋白-10和巨噬细胞炎症蛋白-1β。与对照组相比,经10 mg/kg/周6.3G9治疗的小鼠的可溶性CD40和巨噬细胞炎性蛋白-2水平显著升高,但在经1、3或6 mg/kg/星期6.3G9处理的小鼠中没有显著升高。这些蛋白质(其中许多是炎症介质)的剂量-反应行为与炎症细胞计数的变化密切相关(如前所述)。
图5。
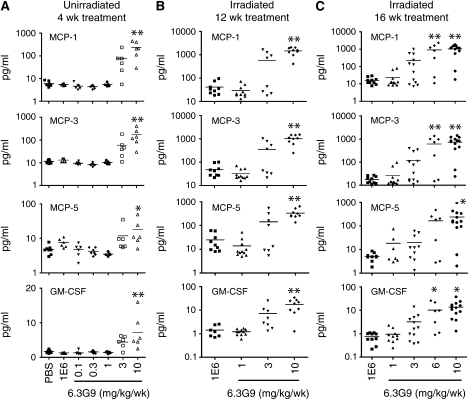
高剂量和低剂量6.3G9对支气管肺泡灌洗(BAL)液蛋白浓度的差异影响。(A类)用对照单克隆抗体(mAb)或指示剂量的6.3G9治疗未受照射的小鼠4周,然后测量BAL液中指示蛋白质的浓度(每组5–8个)。(B类和C类)从照射后16周开始,用对照单克隆抗体或指示剂量的6.3G9治疗受照射小鼠12或16周。然后测量从这些小鼠获得的BAL液中指示蛋白质的浓度*P(P)< 0.05, **P(P)与对照抗体治疗相比,<0.01(每组n=7-13)。
基因表达分析
我们进行了Affymetrix基因芯片分析,分析了按照图3在照射后26周收集肺部进行RNA提取。一种αvβ6依赖性疾病机制的分子特征被鉴定为一组595个基因芯片探针,在辐射和αvβ5阻断的作用下,正常信号强度发生显著变化(看见在线补遗中的表E1)。分层聚类(图6)与PBS和1E6对照组相比,原始对照组和1 mg/kg/周6.3G9组的相应转录表达谱显示基因表达模式有相当大的相似性,而rsTGF-βRII-Fc和低剂量(0.3 mg/kg/wk)与对照损伤(PBS和1E6)组相比,6.3G9剂量对基因表达的影响较小。rsTGFbRII-Fc的作用类似于1 mg/kg/周6.3G9,但没有那么强。这些结果与在1 mg/kg/周6.3G9和可溶性受体治疗组中观察到的纤维化减少相一致。基因列表的功能注释表明,受辐射和6.3G9影响的基因与细胞周期调控机制密切相关,尤其是与p21依赖的G1/S检查点控制相关(数据未显示)。这些结果与辐照细胞的微阵列分析结果相似(24,25). 细胞周期素依赖性激酶抑制剂p21是细胞周期进展的抑制剂和凋亡的介体,是TGF-β的靶点(26),并且在受照小鼠的肺部显示出最高的上调倍数。这些结果表明,通过可溶性受体或阻断αvβ6对TGF-β的抑制可以有效地部分正常化表征辐射损伤反应的基因表达模式,即使在最初的损伤后很久才开始治疗。
图6。

经6.3G9、重组可溶性转化生长因子-β受体II–Fc融合蛋白(rsTGF-βRII–Fc)、对照IgG或PBS照射的小鼠基因表达模式的微阵列分析图3照射16周后开始治疗,26周后处死小鼠。所示的选定基因在未经辐照的小鼠和经辐照的PBS处理的小鼠以及经PBS和1 mg/kg/周6.3G9处理的小鼠中被鉴定为差异表达。基因表达数据的层次聚类显示,PBS、对照IgG或0.3 mg/kg/周6.3G9(一种不能防止纤维化的剂量)治疗的小鼠之间存在相似性。相反,在用1 mg/kg/周6.3G9或rsTGF-βRII–Fc处理的小鼠中,这些基因的表达模式与未经照射的小鼠的表达模式相似。增加的基因以逐渐变亮的阴影表示红色,减少的基因表现为逐渐变亮的阴影绿色和未改变的基因黑色.
讨论
TGF-β信号转导是肺纤维化潜在的治疗靶点。减少TGF-β信号传导的一种方法是阻止潜在TGF-α的激活,我们在小鼠RILF模型中测试了这一策略。该模型的一个有趣方面是纤维化的延迟和逐渐发作,可以说,与常用的博来霉素模型相比,纤维化更类似于人类的情况,如特发性肺纤维化。临床上,RILF是肺癌放射治疗的一种潜在的严重并发症,即使对治疗性肺辐射剂量进行了严格限制,但目前尚无有效的治疗方法(27).
与其他器官相比,肺对辐射损伤相对敏感。实际上,辐射会导致DNA损伤,导致细胞损伤和死亡。随后,在暴露后几周到几个月的时间内,可能会发生肺炎阶段。在患者中,肺炎表现为发热、咳嗽和X线片浸润,通常对皮质类固醇有反应。在啮齿动物模型中,TGF-β1的表达在急性期和肺炎期均增加(28–30). 在稍后的时间点(通常小鼠为5-6个月,人类为>6个月),可能会发生纤维化。缓解不利辐射影响的策略可能允许用较高的辐射剂量治疗肺部恶性肿瘤,从而获得更好的结果。
我们发现αvβ6的表达在时间、空间和因果上与RILF相关。照射肺的肺泡上皮中αvβ6的上调发生在纤维化之前,并持续存在于纤维化病变中。此外,Itgb6号机组−/−小鼠不会发生RILF,而在RILF模型的纤维化阶段之前和期间给予的αvβ6的特异性抑制剂可以在正常发病后12周内预防纤维化。可溶性TGF-β受体也可阻止RILF。全局TGF-β抑制剂和抗αvβ6抑制剂的等效抗纤维化疗效进一步证明,αvβ6介导的TGF-β激活是纤维化的主要原因。目前已知αvβ6参与两种不同的肺纤维化模型(博莱霉素和辐射诱导)以及两种肾纤维化模型(18,31).
缺乏αvβ6的小鼠发生淋巴细胞性肺部炎症(16). 我们注意到高剂量6.3G9处理的受照小鼠肺部淋巴细胞和中性粒细胞的百分比增加。我们没有在组织切片中观察到中性粒细胞增加Itgb6号机组−/−辐射暴露后的肺部(数据未显示)。因此,在受照小鼠中,存在与高水平αvβ6抑制相关的未定义炎症刺激,导致肺中性粒细胞积聚,而在αvβ6-null小鼠中不会发生。细胞学检查结果与BAL液中几种炎症相关细胞因子的增加相一致,但剂量仅为6–10 mg/kg/周6.3G9(图5). 然而,值得注意的是,1 mg/kg/周的6.3G9剂量可在不改变BAL细胞计数的情况下预防纤维化(图3和4)或选定的炎症介质(图5).
小鼠RILF模型在纤维化阶段的死亡率很高。Sharplin和Franko的研究表明,三种类型的肺损伤可能会导致死亡:水肿和透明膜阻塞导致功能性腺泡丧失,广泛收缩性纤维化,以及伴随RVH的肺灌注丧失(22,23). 这些异常的相对贡献取决于小鼠的应变和照射后的时间。在没有纤维化的情况下,一些品系的小鼠在肺照射后生存率严重降低,这表明纤维化显然不是该模型中死亡的唯一可能因素(19). 然而,Sharplin和Franko推测,C57BL/6J小鼠的广泛纤维化是该小鼠品系辐射后致死的原因。这一观点间接得到了以下事实的支持:一些干预措施降低了暴露于肺辐射的C57BL/6小鼠的致死率和纤维化(32–35).
在我们的研究中,对纤维化和致死性的影响是分开的:通过Itgb6号机组缺失或αvβ6抑制不能提高存活率。因此,在这些研究中,纤维化似乎不是死亡的主要原因。这可能与肺灌注减少有关,因为在研究期间死亡的小鼠中发现了RVH,但在存活的小鼠中没有发现RVH,无论治疗如何。因此,我们的证据,根据之前的工作进行解释(23),表明即使在预防纤维化的情况下,C57BL/6小鼠也会发生肺灌注的损失,并且肺灌注的损失至少是该模型中死亡的部分原因。
辐射损伤引发一系列事件,包括血管损伤、炎症、氧化损伤、缺氧和多种信号通路的改变(36,37)这将持续数月。这些事件的一个后果是肺泡上皮细胞αvβ6的表达增加,使更多的TGF-β被激活。其他后果(如血管功能不全)明显发生在αvβ6上调和纤维化的上游或并行。从这个角度来看,αvβ6介导的TGF-β活化是RILF的最直接原因,而不是氧化剂、CD40/CD40L相互作用和血小板衍生生长因子信号传导等因素。我们试图对内皮标记物、血小板内皮细胞粘附分子-1和血管性血友病因子进行染色,但未能进一步了解这些小鼠的血管异常情况(数据未显示)。随着这一过程的描述,确定这些小鼠中假定的血管功能障碍是否是αvβ6介导的纤维化反应所必需的将是一件有趣的事情。
TGF-β信号可能是导致αvβ6表达增加和纤维化的早期事件的一个组成部分,也是纤维化过程中所需的一个因素。放射性损伤后第一天肺和血清TGF-β水平升高(28,30,36). 照射时使用TGF-β拮抗剂进行短期治疗可减少急性肺炎,令人惊讶的是,照射后6个月出现迟发性纤维化(30,38). 同样,在急性肺损伤中,αvβ6激活的TGF-β介导早期毛细血管渗漏(17),并且也是随后的纤维化所必需的。在辐射暴露期间,TGF-β的活化可以通过另一种途径发生,包括电离辐射对LAP的直接影响(10).
该模型中的存活率是毒性的敏感指标,因为小鼠状态脆弱,治疗持续时间长(长达16周)。我们发现高剂量6.3G9有恶化生存率的不一致趋势。还需要进一步研究来确定高剂量6.3G9是否影响生存,以及这种影响是由于炎症加剧还是其他一些过程。
然而,我们的数据表明,当剂量为6.3G9时,可获得抗纤维化效果,且不会导致BAL炎性细胞或存活率发生显著变化。高剂量6.3G9(6–10 mg/kg/wk)会引起BAL细胞计数和巨噬细胞形态的变化,与Itgb6号机组−/−这表明这些剂量会导致αvβ6功能几乎完全丧失。每周1–3毫克/千克的剂量对炎性细胞计数的影响最小,同时仍能防止纤维化。重要的是,尽管产生抗纤维化和促炎作用的抗αvβ6单克隆抗体的注射剂量相差不到10倍,但这些剂量所获得的循环血浆水平的差异明显更大。药代动力学分析(未公布的数据)表明,6.3G9在1 mg/kg剂量下的半衰期明显短于10 mg/kg剂量,这两种剂量下的暴露差异(以浓度-时间曲线下的面积衡量)约为80倍。这些结果表明,当6.3G9的浓度显著低于那些在肺部产生炎症变化的浓度时,可以达到近最大的抗纤维化活性。
TGF-β在不同浓度下引起不同的细胞反应(39). 我们的结果表明,表达αvβ6的上皮细胞产生的少量活性TGF-β足以抑制炎症,而导致纤维化则需要更多的量。这些差异可能是由于靶细胞(如淋巴细胞和成纤维细胞)反应的内在差异,或者是由于从上皮细胞向不同靶细胞主动传递TGF-β的效率不同。无论如何,这一观点得到了以下观察结果的支持:Tgfb1型-单倍体小鼠相对而言受到RILF的保护(图4E)炎症没有明显增加(数据未显示)。
放射治疗18周后αvβ6表达增加;有趣的是,大鼠肺部照射后18周,血浆转化生长因子-β水平急剧升高(29). αvβ6上调也紧随博莱霉素诱导的纤维化(13). 这些相关性并不能证明纤维化需要高水平的αvβ6,但腺病毒介导的正常肺中潜在TGF-β1的过度表达不会导致纤维化(40)表明正常肺的TGF-β活化能力是纤维化的速率限制。αvβ6表达增加可能不是增加αvβ6-依赖性TGF-β活化的唯一途径;例如,蛋白酶激活受体-1信号增强αvβ6功能(41),和潜在的TGF-β结合蛋白-1增强表达αvβ6的细胞对TGF-β的激活(42). TGF-β增加培养细胞中β6的表达(43),但在我们的研究中,我们没有发现TGF-β诱导的αvβ6上调的证据,因为用6.3G9或rsTGF-?RII-Ig治疗的小鼠仍有强烈上调的αv?β6表达(数据未显示)。
抑制TGF-β信号转导可能对纤维化和其他疾病有益,并且在小鼠中耐受性良好(44). TGF-β拮抗剂的早期临床试验正在进行中。然而,来自遗传小鼠模型的数据表明,TGF-β活化或信号转导减少可能导致炎症加重、肺气肿和恶性肿瘤增加。我们的数据表明,最佳的抗TGF-β策略可能需要仔细滴定,部分抑制TGF-?活性,以最大限度地发挥效益,并将潜在毒性降至最低。在人体研究中解决这个问题需要比较不同的给药方案以及治疗和不良反应的定量标记。
总之,我们的数据和其他结果表明,αvβ6–TGF-β轴在各种形式的肺和肾纤维化中起着关键作用(13,18). 直接抑制αvβ6并可能预防其上调是治疗或预防RILF和其他形式纤维化的潜在有用策略。
致谢
作者感谢迪安·谢泼德Itgb6号机组−/−感谢保罗·雷霍恩的实验帮助。
由美国国立卫生研究院资助HL077526和HL063786,并由Biogen Idec(J.S.M.)资助。
最初出版于2007年10月4日,作为DOI:10.1164/rccm.200706-806OC出版
利益冲突声明:K.P.与与本手稿主题相关的商业实体没有财务关系。N.H.与与本手稿主题相关的商业实体没有财务关系。S.C.J.与对本手稿主题感兴趣的商业实体没有财务关系。E.B.与与本手稿主题相关的商业实体没有财务关系。Z.Z.与与本手稿主题相关的商业实体没有财务关系。Z.Y.与对本手稿主题感兴趣的商业实体没有财务关系。M.L.D.与对本手稿主题感兴趣的商业实体没有财务关系。G.S.H.是Biogen Idec的员工,领取工资和股票的年薪。P.H.W.是Biogen Idec的员工和股东。M.E.L.是Biogen Idec的员工,领取工资和股票的年薪。S.M.V.是Biogen Idec的员工,领取工资和股票的年薪。K.S.G.与与本手稿主题相关的商业实体没有财务关系。C.C.与与本手稿主题相关的商业实体没有财务关系。S.C.F.与与本手稿主题相关的商业实体没有财务关系。2004年至2007年,J.S.M.获得了Biogen Idec的研究资助。
工具书类
-
1.查普曼HA。肺基质重塑障碍。临床投资杂志2004;113:148–157.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
2佛兰德斯KC。Smad3作为纤维化反应的介质。国际实验病理学杂志2004;85:47–64.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
三。Branton MH,Kopp JB公司。TGF-β与纤维化。1999年《微生物感染》;1:1349–1365.[内政部] [公共医学] [谷歌学者]
-
4Hyytainen M,Penttinen C,Keski-Oja J.潜在TGF-β结合蛋白:细胞外基质关联和在TGF-?激活中的作用。Crit Rev Clin Lab Sci 2004;41:233–264.[内政部] [公共医学] [谷歌学者]
-
5Annes JP、Munger JS、Rifkin DB。了解潜在的TGFβ激活。《细胞科学杂志》2003;116:217–224.[内政部] [公共医学] [谷歌学者]
-
6Kojima S、Nara K、Rifkin DB。激活牛内皮细胞中潜在转化生长因子-β对谷氨酰胺转胺酶的需求。细胞生物学杂志1993;121:439–448.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
7Yu Q,Stamenkovic I.细胞表面局部基质金属蛋白酶-9蛋白水解激活TGF-β并促进肿瘤侵袭和血管生成。基因开发2000;14:163–176.[PMC免费文章] [公共医学] [谷歌学者]
-
8Crawford SE、Stellmach V、Murphy-Ullrich JE、Ribeiro SM、Lawler J、Hynes RO、Boivin GP、Bouck N.血栓反应蛋白-1是TGF-β1的主要激活剂体内.单元格1998;93:1159–1170.[内政部] [公共医学] [谷歌学者]
-
9Murphy-Ullrich JE,Poczatek M.血小板反应蛋白-1对潜在TGF-β的激活:机制和生理学。细胞因子生长因子Rev 2000;11:59–69.[内政部] [公共医学] [谷歌学者]
-
10Jobling MF、Mott JD、Finnegan MT、Jurukovski V、Erickson AC、Walian PJ、Taylor SE、Ledbetter S、Lawrence CM、Rifkin DB、,等.活性氧物种对潜在转化生长因子β(LTGF-β)的等形态特异性激活。辐射研究2006;166:839–848.[内政部] [公共医学] [谷歌学者]
-
11Mu D、Cambier S、Fjellbirkeland L、Baron JL、Munger JS、Kawakatsu H、Sheppard D、Broaddus VC、Nishimura SL。整合素αvβ8通过MT1-MMP-依赖性TGF-β1激活介导上皮内稳态。细胞生物学杂志2002;157:493–507.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
12Annes JP、Rifkin DB、Munger JS。整合素αvβ6结合并激活潜在的TGFβ3。FEBS Lett 2002;511:65–68.[内政部] [公共医学] [谷歌学者]
-
13.Munger JS、Huang X、Kawakatsu H、Griffiths MJ、Dalton SL、Wu J、Pittet JF、Kaminski N、Garat C、Matthay MA、,等整合素αvβ6结合并激活潜在的TGF-β1:一种调节肺部炎症和纤维化的机制。细胞1999;96:319–328.[内政部] [公共医学] [谷歌学者]
-
14Yang Z、Mu Z、Dabovic B、Jurukovski V、Yu D、Sung J、Xiong X、Munger JS。体内缺乏整合素介导的TGFβ1激活,重述了TGFβ1-null小鼠的表型。细胞生物学杂志2007;176:787–793.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
15Morris DG、Huang X、Kaminski N、Wang Y、Shapiro SD、Dolganov G、Glick A、Sheppard D。整合素αvβ6介导的TGF-β活化缺失导致MMP12依赖性肺气肿。自然2003;422:169–173.[内政部] [公共医学] [谷歌学者]
-
16Huang XZ,Wu JF,Cass D,Erle DJ,Corry D,Young SG,Farese RV,Sheppard D。整合素β6亚基基因的失活揭示了上皮整合素在调节肺部和皮肤炎症中的作用。细胞生物学杂志1996;133:921–928.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
17Pittet JF、Griffiths MJ、Geiser T、Kaminski N、Dalton SL、Huang X、Brown LA、Gotwals PJ、Koteliansky VE、Matthay MA、,等TGF-β是急性肺损伤的重要介质。临床投资杂志2001;107:1537–1544.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
18Hahm K、Lukashev ME、Luo Y、Yang WJ、Dolinski BM、Weinreb PH、Simon KJ、Chun Wang L、Leone DR、Lobb RR、,等αvβ6整合素调节阿尔波特小鼠的肾纤维化和炎症。《美国病理杂志》2007年;170:110–125.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
19Haston CK、Zhou X、Gumbiner-Russo L、Irani R、Dejournett R、Gu X、Weil M、Amos CI、Travis EL。通用和辐射特异性基因座影响小鼠对辐射诱导肺纤维化的敏感性。2002年癌症研究;62:3782–3788.[公共医学] [谷歌学者]
-
20Reddy GK,Enwemeka CS公司。生物组织中羟脯氨酸的简化分析方法。临床生物化学1996;29:225–229.[内政部] [公共医学] [谷歌学者]
-
21Weinreb PH、Simon KJ、Rayhorn P、Yang WJ、Leone DR、Dolinski BM、Pearse BR、Yokota Y、Kawakatsu H、Atakilit A、,等功能块整合素αvβ6单克隆抗体:不同的类配体和非类配体。生物化学杂志2004;279:17875–17887.[内政部] [公共医学] [谷歌学者]
-
22Sharplin J,Franko AJ。定量组织学研究早期辐射对小鼠肺部影响的应变依赖性差异。辐射研究1989;119:1–14.[公共医学] [谷歌学者]
-
23Sharplin J,Franko AJ。定量组织学研究中后期小鼠肺部辐射效应的应变依赖性差异。辐射研究1989;119:15–31.[公共医学] [谷歌学者]
-
24Zhou T、Chou JW、Simpson DA、Zhou Y、Mullen TE、Medeiros M、Bushel PR、Paules RS、Yang X、Hurban P、,等电离辐射损伤的人类二倍体成纤维细胞中的全局基因表达谱显示G1检查点在G0样静止状态下的同步性。《2006年环境健康展望》;114:553–559.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
25Jen KY,Cheung VG。电离辐射反应中新的p53靶基因的鉴定。2005年癌症研究;65:7666–7673.[内政部] [公共医学] [谷歌学者]
-
26Datto MB、Li Y、Panus JF、Howe DJ、Xiong Y、Wang XF。转化生长因子β通过p53依赖机制诱导细胞周期素依赖性激酶抑制剂p21。1995年美国国家科学院院刊;92:5545–5549.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
27Kong FM,Ten Haken R,Eisbruch A,Lawrence TS。非小细胞肺癌治疗相关的肺毒性:放射性肺炎和纤维化的最新进展。Semin Oncol 2005;32:S42–S54。[内政部] [公共医学] [谷歌学者]
-
28Rubin P、Johnston CJ、Williams JP、McDonald S、Finkelstein JN。辐射后细胞因子的持续级联导致肺纤维化。国际放射肿瘤生物学杂志1995;33:99–109.[内政部] [公共医学] [谷歌学者]
-
29Vujaskovic Z,Down JD,van Waarde MA,van Assen AJ,Szabo BG,Konings AW.大鼠半胸照射后血浆TGFβ水平。Radiother Oncol 1997;44:41–43.[内政部] [公共医学] [谷歌学者]
-
30Haiping Z、Takayama K、Uchino J、Harada A、Adachi Y、Kura S、Caichun Z、Tsuzuki T、Nakanishi Y。通过重组腺病毒介导转移可溶性TGF-βII型受体基因预防放射性肺炎。癌症基因疗法2006;13:864–872.[内政部] [公共医学] [谷歌学者]
-
31Ma LJ,Yang H,Gaspert A,Carlesso G,Barty MM,Davidson JM,Sheppard D,Fogo AB。转化生长因子-β依赖性和非依赖性途径诱导β6肾小管间质纤维化−/−老鼠。《美国病理学杂志》2003;163:1261–1273.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
32Adawi A、Zhang Y、Baggs R、Rubin P、Williams J、Finkelstein J、Phipps RP。阻断CD40–CD40配体相互作用可防止辐射诱导的肺部炎症和纤维化。临床免疫病理学1998;89:222–230.[内政部] [公共医学] [谷歌学者]
-
33.Rabbani ZN、Anscher MS、Folz RJ、Archer E、Huang H、Chen L、Golson ML、Samulski TS、Dewhirst MW、Vujaskovic Z。细胞外超氧化物歧化酶的过度表达可降低急性辐射诱导的肺毒性。BMC癌症2005;5:59.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
34Tabata C、Kadokawa Y、Tabata R、Takahashi M、Okoshi K、Sakai Y、Mishima M、Kubo H。全反式维甲酸可预防辐射或博莱霉素诱导的肺纤维化。Am J Respir Crit Care Med 2006;174:1352–1360.[内政部] [公共医学] [谷歌学者]
-
35Abdollahi A、Li M、Ping G、Plathow C、Domhan S、Kiessling F、Lee LB、McMahon G、Grone HJ、Lipson KE、,等血小板衍生生长因子信号传导抑制可减轻肺纤维化。《实验医学杂志》2005;201:925–935.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
36希尔RP。辐射对呼吸系统的影响。BJR Suppl 2005;27:75–81.15975876[谷歌学者]
-
37.Vujaskovic Z、Anscher MS、Feng QF、Rabbani ZN、Amin K、Samulski TS、Dewhirst MW、Haroon ZA。辐射诱导的缺氧可能使晚期正常组织损伤永久化。国际放射肿瘤生物物理杂志2001;50:851–855.[内政部] [公共医学] [谷歌学者]
-
38Anscher MS、Thrasher B、Rabbani Z、Teicher B和Vujaskovic Z。抗转化生长因子-β抗体1D11可改善高剂量辐射引起的正常组织损伤。国际放射肿瘤生物物理杂志2006;65:876–881.[内政部] [公共医学] [谷歌学者]
-
39.Wu DT、Bitzer M、Ju W、Mundel P、Bottinger EP。TGF-β浓度规定了足细胞生长停滞/分化和凋亡的不同信号模式。《美国肾脏学会杂志》2005;16:3211–3221.[内政部] [公共医学] [谷歌学者]
-
40Sime PJ,Xing Z,Graham FL,Csaky KG,Gauldie J.腺病毒介导的活性转化生长因子-β1基因转移诱导大鼠肺长期严重纤维化。《临床投资杂志》1997年;100:768–776.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
41Jenkins RG、Su X、Su G、Scotton CJ、Camerer E、Laurent GJ、Davis GE、Chambers RC、Matthay MA、Sheppard D。蛋白酶激活受体1的连接增强αvβ6整合素依赖性TGF-β的激活并促进急性肺损伤。临床投资杂志2006;116:1606–1614.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
42Annes JP、Chen Y、Munger JS、Rifkin DB。整合素αvβ6介导的潜在TGF-β激活需要潜在TGF--β结合蛋白-1。细胞生物学杂志2004;165:723–734.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
43Wang A,Yokosaki Y,Ferrando R,Balmes J,Sheppard D。生长因子对气道上皮整合素的差异调节。美国呼吸细胞分子生物学杂志1996;15:664–672.[内政部] [公共医学] [谷歌学者]
-
44Yang YA、Dukhanina O、Tang B、Mamura M、Letterio JJ、MacGregor J、Patel SC、Khozin S、Liu ZY、Green J、,等终身暴露于可溶性TGF-β拮抗剂可保护小鼠免受转移,而不会产生不良副作用。临床投资杂志2002;109:1607–1615.[内政部] [PMC免费文章] [公共医学] [谷歌学者]
关联数据
本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。

