

摘要
材料和方法
抗体和细胞
pAbs的生产
图1。
![]()
表达载体的构建及转染
免疫染色
SDS-PAGE和免疫印迹
冷冻断裂电子显微镜
结果
检测Claudin-5/TMVCF抗体的制备
Claudin-5/TMVCF在脑和肺中的作用
图2。
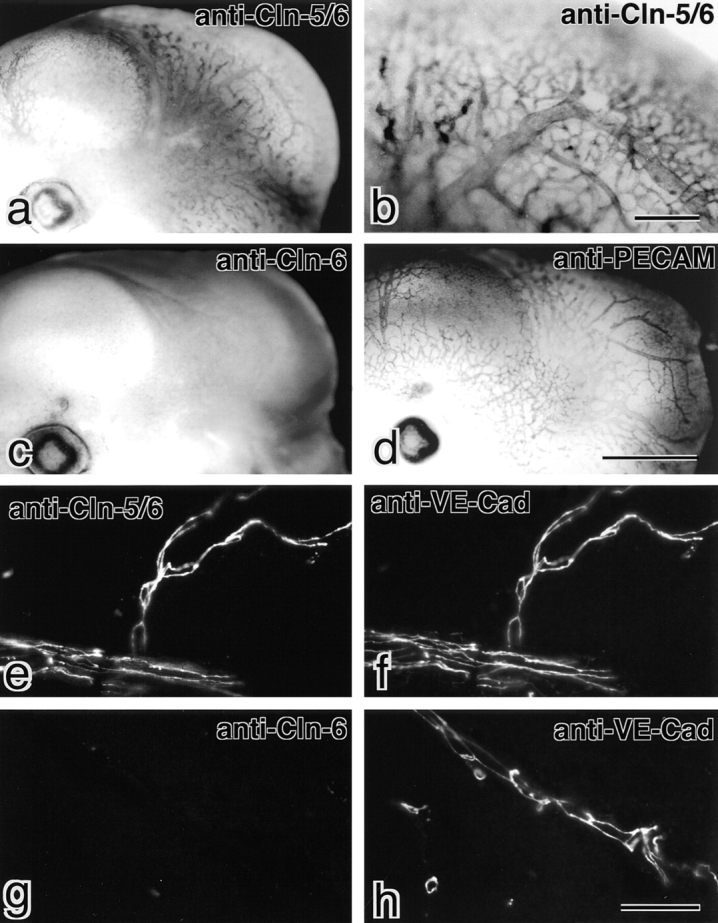
图3。
![]()
图4。
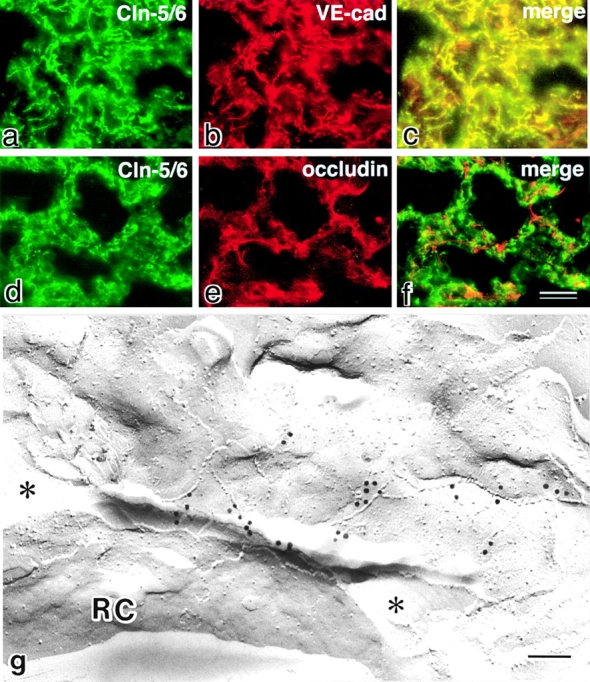
肾脏和其他器官中的Claudin-5/TMVCF
图5。
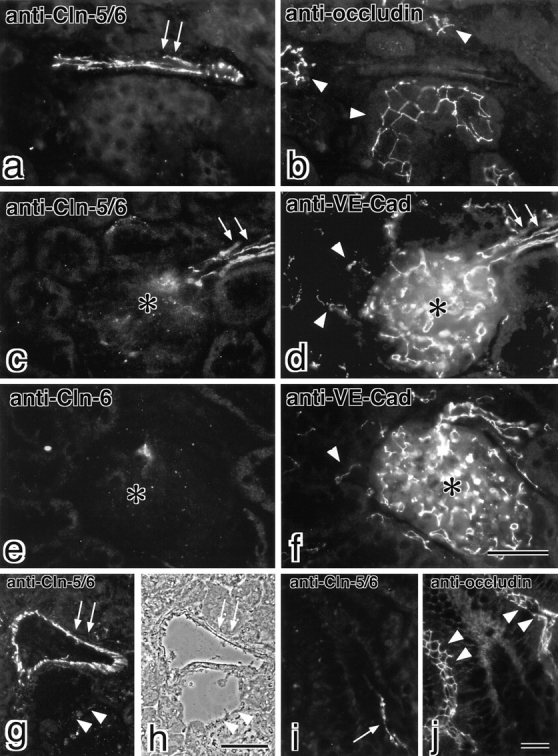
Claudin-5/TMVCF在L成纤维细胞中重组胞外表面相关TJ链
图6。

讨论
致谢
脚注
工具书类
-
Anderson J.M.、Van Itallie C.M.紧密连接和细胞旁通透性调节的分子基础。 美国生理学杂志。 1995; 269:G467–G475。 doi:10.1152/ajpgi.1995.269.4.G467。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ando-Akatsuka Y.、Saitou M.、Hirase T.、Kishi M.,Sakakibara A.、Itoh M.、Yonemura S.、Furuse M.、Tsukita Sh.克隆人类、小鼠、狗和大鼠-袋鼠同源物的闭塞素序列cDNA的种间多样性。 《细胞生物学杂志》。 1996; 133:43–47. doi:10.1083/jcb.133.1.43。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Balda M.S.、González-Mariscal L.、Matter K.、Cereijido M.、Anderson J.M.紧密连接的组装-二酰甘油的作用。 细胞生物学杂志。 1993; 123:293–302. doi:10.1083/jcb.123.2.293。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Balda M.S.、Whitney J.A.、Flores C.、González S.、Cereijido M.、Matter K.通过突变紧密连接膜蛋白的表达,细胞旁通透性和跨上皮电阻的功能分离以及顶基底外侧膜内扩散屏障的破坏。 细胞生物学杂志。 1996; 134:1031–1049. doi:10.1083/jcb.134.4.1031。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Bowman P.D.、du Bois M.、Shivers R.R.、Dorovini-Zis K.内皮紧密连接。 作者:Cereijido M.,编辑。 紧密连接。 CRC出版社; 伦敦:1992年。 第305-320页。 [ 谷歌学者 ] -
Brightman M.W.、Reese T.S.,脊椎动物大脑中紧贴的细胞膜之间的连接。 《细胞生物学杂志》。 1969; 40:648–677. doi:10.1083/jcb.40.3.648。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Chen Y.-H.,Merzdorf C.,Paul D.L.,Goodenough D.A.闭塞蛋白的COOH末端是早期爪蟾胚胎紧密连接屏障功能所必需的。 《细胞生物学杂志》。 1997; 138:891–899. doi:10.1083/jcb.138.4.891。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Chen Z.,Zandonatti M.,Jakubowski D.,Fox H.S.脑毛细血管内皮细胞表达MBEC1,这是一种与产气荚膜梭菌肠毒素受体相关的蛋白质。 实验室投资。 1998; 78:353–363. [ 公共医学 ] [ 谷歌学者 ] -
Citi S.、Sabanay H.、Jakes R.、Geiger B.、Kendrick-Jones J.Cingulin,紧密连接的新外围组件。 自然。 1988; 333:272–276. doi:10.1038/333272a0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Dempsey G.P.,Bullivant S.《冷冻断裂法观察到的内皮细胞膜颗粒极性》,《科学》。 1973; 179:190–192. doi:10.126/science.179.4069.190。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Farquhar M.G.、Palade G.E.各种上皮中的连接复合体。 《细胞生物学杂志》。 1963; 17:375–412. doi:10.1083/jcb.17.2.375。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Farrell C.L.,Risau W.血脑屏障的正常和异常发育。 微型计算机。 Res.Tech.1994; 27:495–506。 doi:10.1002/jemt.1070270604。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Fujimoto K.冷冻断裂复型电镜结合SDS消化用于细胞化学标记整体膜蛋白。 应用于细胞间连接复合体的免疫金标记。 细胞科学杂志。 1995; 108:3443–3449。 doi:10.1242/jcs.108.11.3443。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Furuse M.,Hirase T.,Itoh M.,Nagafuchi A.,Yonemura S.,Sa.Tsukita,Tsukita-Sh.Occludina定位于紧密连接处的新型整体膜蛋白。 《细胞生物学杂志》。 1993; 123:1777–1788. doi:10.1083/jcb.123.6.1777。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Furuse M.、Fujimoto K.、Sato N.、Hirase T.、Sa.Tsukita、Tsukita-Sh。闭塞素(一种紧密连接相关的整体膜蛋白)的过度表达诱导了细胞内具有紧密连接样结构的多层膜小体的形成。 细胞科学杂志。 1996; 109:429–435. doi:10.1242/jcs.109.2.429。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Furuse M.、Fujita K.、Hiiragi T.、Fujiamoto K.、Tsukita Sh。 Claudin-1和-2定位于紧密连接处的新的整合膜蛋白,与闭塞蛋白的序列无相似性 《细胞生物学杂志》。 141 1998 1539 1550年 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Furuse M.、Sasaki H.、Fujimoto K.、Tsukita Sh。 一种单基因产物,claudin-1或-2,在成纤维细胞中重建紧密连接链和新的闭塞素 《细胞生物学杂志》。 143 1998 391 401b个 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Gale N.W.、Yancopoulos G.D.血管发育中通过内皮细胞特异性受体酪氨酸激酶VEGF、血管生成素和肾上腺素作用的生长因子。 基因开发1999; 13:1055–1066. doi:10.1101/gad.13.9.1055。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
很好,D.A.堵住泄漏。 程序。 国家。 阿卡德。 科学。 美国1999年; 96:319–321. doi:10.1073/pnas.96.2.319。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Gumbiner B.上皮紧密连接的结构、生物化学和组装。 美国生理学杂志。 1987; 253:C749–C758。 doi:10.1152/ajpcell.1987.253.6.C749。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Gumbiner B.突破紧密连接屏障。 《细胞生物学杂志》。 1993; 123:1631–1633. doi:10.1083/jcb.123.6.1631。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Gumbiner B.、Lowenkopf T.、Apatira D.与紧密连接蛋白ZO-1结合的160-kDa多肽的鉴定。 程序。 国家。 阿卡德。 科学。 美国,1991年; 88:3460–3464。 doi:10.1073/pnas.88.8.3460。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Haskins J.、Gu L.、Wittchen E.S.、Hibbard J.、Stevenson B.R.ZO-3是在紧密连接处发现的MAGUK蛋白家族的新成员,与ZO-1和闭塞蛋白相互作用。 《细胞生物学杂志》。 1998; 141:199–208。 doi:10.1083/jcb.141.1.199。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Hirase T.、Staddon J.M.、Saitou M.、Ando-Akatsuka Y.、Itoh M.、Furuse M.、藤本K.、Tsukita Sh.、Rubin L.L.Occludin可能是内皮细胞紧密连接通透性的决定因素。 细胞科学杂志。 1997; 110:1603–1613. doi:10.1242/jcs.110.14.1603。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Keon B.H.、Schäfer S.、Kuhn C.、Grund C.、Franke W.W.Symplekin,一种新型紧密连接斑块蛋白。 《细胞生物学杂志》。 1996; 134:1003–1018. doi:10.1083/jcb.134.4.1003。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Kevil C.G.、Payne D.K.、Mire E.、Alexander J.S.血管通透性因子/血管内皮细胞生长因子介导的通透性通过内皮连接蛋白的紊乱发生。 生物学杂志。 化学。 1998; 273:15099–15103. doi:10.1074/jbc.273.24.15099。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Kniesel U.,Risau W.,Wolburg H.大鼠皮层血脑屏障紧密连接的发育。 《Dev.Brain Res.1996》; 96:229–240. doi:10.1016/0165-3806(96)00117-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Laemmli U.K.噬菌体T4头部组装期间结构蛋白的裂解。 自然。 1970; 227:680–685. doi:10.1038/227680a0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Lampugani M.G.、Corada M.、Caveda L.、Breviario F.、Ayalon O.、Geiger B.、Dejana E.内皮细胞与细胞连接的分子组织斑球蛋白、β-连环蛋白和α-连环素与血管内皮钙粘蛋白(VE-cadherin)的不同联系《细胞生物学杂志》。 1995; 129:203–217. doi:10.1083/jcb.129.1.203。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
McCarthy K.M.、Skare I.B.、Stankewich M.C.、Furuse M.、Tsukita Sh.、Rogers R.A.、Lynch R.D.、Schneeberger E.E.Occludin是紧密连接的功能组件。 细胞科学杂志。 1996; 109:2287–2298. doi:10.1242/jcs.109.9.2287。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Møllgård K.,Saunders N.R.胎儿早期大脑中上皮细胞和内皮细胞的复杂紧密连接。 《神经细胞学杂志》。 1975; 4:453–468. doi:10.1007/BF01261375。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
森田K.、福鲁斯M.、藤本K.、津田Sh。 Claudin多基因家族编码紧密连接链的四个跨膜结构域蛋白组分 程序。 国家。 阿卡德。 科学。 美国。 96 1999 511 516年 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
森田K.、佐佐木H.、藤本K.、福鲁斯M.、津田Sh。 睾丸少突胶质细胞有髓鞘和支持细胞Claudin-11/OSP基紧密连接 《细胞生物学杂志》。 145 1999 579 588亿 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Niwa H.,Yamamura K.,Miyazaki J.使用新型真核载体高效选择高效表达转染剂。 基因。 1991; 108:193–200. doi:10.1016/0378-1119(91)90434-d。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Reese T.J.,Karnovsky M.J.血脑屏障对外源性过氧化物酶的精细结构定位。 《细胞生物学杂志》。 1967; 34:207–217. doi:10.1083/jcb.34.1.207。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Risau W.,Wolburg H.血脑屏障的发展。 《神经科学趋势》。 1990; 13:174–178. doi:10.1016/0166-2236(90)90043-a。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Risau W.,Flamme I.血管生成。 每年。 Rev.细胞发育生物学。 1995; 11:73–91. doi:10.1146/annurev.cb.11.10195.000445。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Risau W.、Esser S.、Engelhardt B.血脑屏障内皮细胞的分化。 病态。 《生物》1998; 46:171–175. [ 公共医学 ] [ 谷歌学者 ] -
Rubin L.L.内皮细胞粘附和紧密连接。 货币。 操作。 细胞生物学。 1992; 4:830–833. doi:10.1016/0955-0674(92)90107-n。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Saitou M.、Ando-Akatsuka Y.、Itoh M.、Furuse M.、Inazawa J.、Fujimoto K.、Tsukita Sh.上皮细胞表达和亚细胞分布中的哺乳动物闭塞素。 《欧洲细胞生物学杂志》。 1997; 73:222–231. [ 公共医学 ] [ 谷歌学者 ] -
Saitou M.、Fujimoto K.、Doi Y.、Itoh M.、Benjimoto T.、Furuse M.、Takano H.、Noda T.、Tsukita Sh.闭锁缺陷胚胎干细胞可以分化为具有紧密连接的极化上皮细胞。 《细胞生物学杂志》。 1998; 141:397–408. doi:10.1083/jcb.141.2.397。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Schneeberger E.E.肺内血管系统不同节段细胞间连接的结构。 纽约学院安。 科学。 1982; 384:54–63. doi:10.1111/j.1749-6632.1982.tb21361.x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Schneeberger E.E.,Lynch R.D.细胞紧密连接的结构、功能和调节。 美国生理学杂志。 1992; 262页:L647–L661。 doi:10.1152/ajplung.1992.262.6.L647。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Simionescu M.,Simionesuc N.,Palade G.E.血管内皮细胞连接的分段分化。 微血管。 《细胞生物学杂志》。 1975; 第67:863–885页。 doi:10.1083/jcb.67.3.863。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Simionescu M.,Simionesuc N.,Palade G.E.血管内皮细胞连接的分段分化。 动脉和静脉。 《细胞生物学杂志》。 1976; 第68:705–723页。 doi:10.1083/jcb.68.3.705。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Sirotkin H.、Morrow B.、Saint-Jore B.、Puech A.、Das Gupta R.、Patanjali S.R.、Skoultchi A.、Weissman S.M.、Kucherlapati R.识别、表征和精确定位人类基因编码的一种新的膜跨蛋白,该蛋白来自于velo-cardio-face syndrome中缺失的22q11区域。 基因组学。 1997; 42:245–251. doi:10.1006/geno.1997.4734。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Spring K.上皮细胞液体转运的途径和机制。 每年。 生理学评论。 1998; 60:105–119. doi:10.1146/annurev.physiol.60.1.105。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Staehelin L.A.关于冷冻叶紧密连接的精细结构的进一步观察。 细胞科学杂志。 1973; 13:763–786. doi:10.1242/jcs.13.3.763。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Staehelin L.A.细胞间连接的结构和功能。 细胞国际评论。 1974; 39:191–283. doi:10.1016/s0074-7696(08)60940-7。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Stevenson B.R.、Siliciano J.D.、Mooseker M.S.、Goodenough D.A.鉴定与多种上皮细胞紧密连接(闭塞小带)相关的高分子量多肽ZO-1。 《细胞生物学杂志》。 1986; 103:755–766. doi:10.1083/jcb.103.3.755。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Tsukita Sh.、Furuse M.Occludin和claudins在紧密结合点上领导或支持球员? 趋势细胞生物学。 1999; 9:268–273. doi:10.1016/s0962-8924(99)01578-0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Van Itallie C.M.和Anderson J.M.Occludin在成纤维细胞中表达时具有粘附性。 细胞科学杂志。 1997; 110:1113–1121. doi:10.1242/jcs.110.9.1113。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Wolburg H.、Neuhaus J.、Kniesel U.、Krauss B.、Schmid E.M.、Öcalan M.、Farrell C.、Risau W.血脑屏障内皮细胞紧密连接结构的调节。 组织培养、第二信使和共培养星形胶质细胞的作用。 细胞科学杂志。 1994; 107:1347–1357. doi:10.1242/jcs.107.5.1347。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Wong V.,Gumbiner B.M.一种与闭塞素胞外结构域相对应的合成肽干扰紧密连接的通透性屏障。 《细胞生物学杂志》。 1997; 136:399–409. doi:10.1083/jcb.136.2399。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Yap A.S.,Mullin J.M.,Stevenson B.R.紧密连接生理现象和悖论的分子分析。 J.成员。 《生物》1998; 163:159–167. doi:10.1007/s002329900380。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Zhong Y.、Saitoh T.、Minase T.、Sawada N.、Enomoto K.、Mori M.单克隆抗体7H6与一种不同于ZO-1、扣带蛋白和ZO-2的新型紧密连接相关蛋白反应。 《细胞生物学杂志》。 1993; 120:477–483. doi:10.1083/jcb.120.2.477。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]