

纤毛的性质
构建纤毛:鞭毛内运输
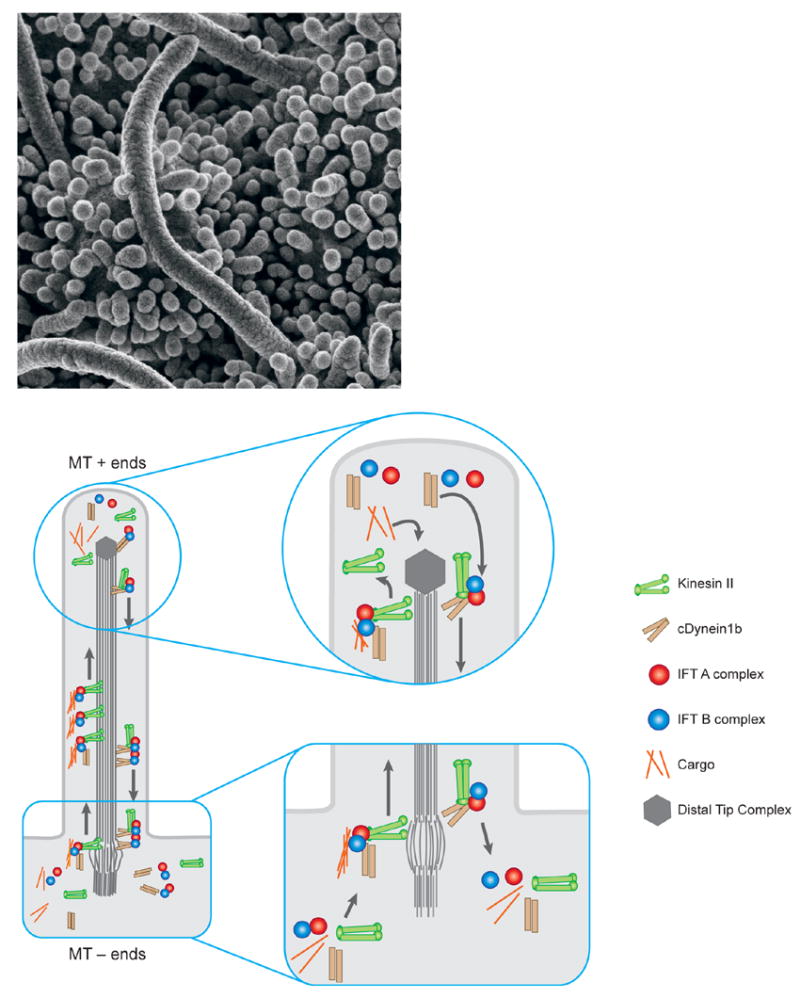
图1。 Cilia和IFT。
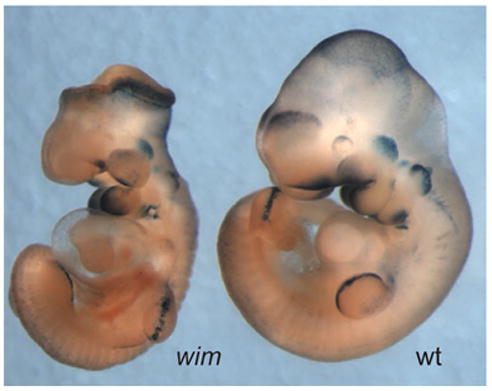
小鼠IFT突变体表明Hh信号与纤毛耦合
表1。
|
|
||||
|
|
||||
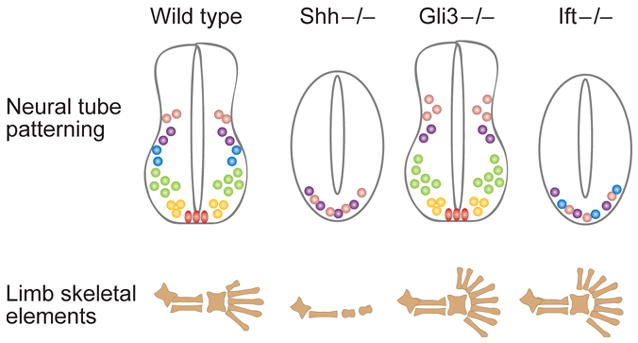
神经管模式分析表明,在细胞质Hh信号转导机制的核心需要IFT蛋白
肢体模式化中的IFT蛋白和Shh信号
图2。 IFT突变体的组织特异性效应取决于组织中Gli激活物和Gli阻遏物的重要性。
声波刺猬和印度刺猬都需要IFT蛋白来传递信号
刺猬路径成分富含纤毛
图3。
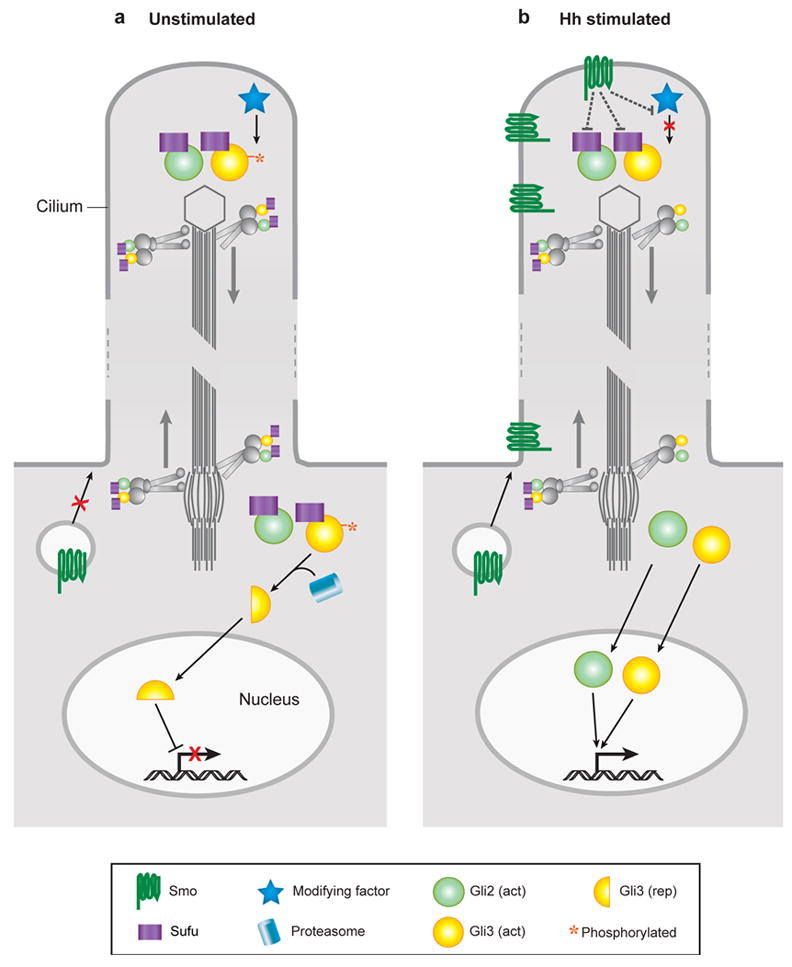
纤毛不仅仅是Hh信号转导机制富集的场所吗?
The phenotypes of the
所有脊椎动物的Hh信号都需要纤毛吗?
还有其他依赖纤毛的发育信号吗?
PDGFRα信号
标准Wnt信号
图4。
非标准Wnt信号
与纤毛相关的人类遗传病是否反映了信号通路的中断?
初级纤毛是信号转导机器吗?
致谢
引用的文献
-
Adler PN。果蝇的平面信号和形态发生。 开发单元。 2002; 2(5):525–35. doi:10.1016/s1534-5807(02)00176-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Afzelius BA。由不动纤毛引起的人类综合征。 科学。 1976; 193(4250):317–9. doi:10.1126/science.1084576。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Afzelius BA。纤毛相关疾病。 病理学杂志。 2004; 204(4):470–7. doi:10.1002/path.1652。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Albrecht-Buehler G.3T3细胞的吞噬动力学轨迹:轨迹片段的方向与包含肌动蛋白或微管蛋白的细胞结构的方向平行。 单元格。 1977; 12(2):333–9. doi:10.1016/0092-8674(77)90109-x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ansley SJ、Badano JL、Blacque OE、Hill J、Hoskins BE等。基础身体功能障碍可能是多效性Bardet-Biedl综合征的原因。 自然。 2003; 425(6958):628–33. doi:10.1038/nature02030。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Avidor Reiss T,Maer AM,Koundakjian E,Polyanovsky A,Keil T等。纤毛功能解码:定义区隔纤毛生物发生所需的专门基因。 单元格。 2004; 117(4):527–39. doi:10.1016/s0092-8674(04)00412-x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Badano JL、Mitsuma N、Beales PL、Katsanis N。纤毛病:一类新兴的人类遗传疾病。 《基因组学与人类遗传学年鉴》。 2006; 7:125–48. doi:10.1146/annurev.genom.708055.115610。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Barnfield PC,Zhang X,Thanabalasingham V,Yoshida M,Hui CC。融合抑制因子通过多种机制对Gli1和Gli2激活物功能的负调控。 区别。 2005; 73(8):397–405. doi:10.1111/j.1432-0436.2005.00042.x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Blacque OE、Li C、Inglis PN、Esmail MA、Ou G等。逆行鞭毛内转运需要WD重复序列蛋白IFTA-1。 分子生物学细胞。 2006; 17(12):5053–62. doi:10.1091/mbc。 E06-06-0571。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Blacque OE,Leroux MR.Bardet-Biedl综合征:细胞内转运的新病理机制。 细胞分子生命科学。 2006; 63(18):2145–61. doi:10.1007/s00018-006-6180-x。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Bostrom H、Willetts K、Pekny M、Leveen P、Lindahl P等。PDGF-A信号是肺泡肌成纤维细胞发育和肺泡形成的关键事件。 单元格。 1996; 85(6):863–73. doi:10.1016/s0092-8674(00)81270-2。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Brailov I、Bancila M、Brisorgueil MJ、Miquel MC、Hamon M、Verge D。大鼠脑神经元纤毛质膜上5-HT(6)受体的定位。 脑研究2000; 872(1–2):271–5. doi:10.1016/s0006-8993(00)02519-1。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Brazelton WJ、Amundsen CD、Silflow CD、Lefebvre PA.bld1突变确定衣原体osm-6同源物是鞭毛组装所需的基因。 当前生物量。 2001; 11(20):1591–4. doi:10.1016/s0960-9822(01)00485-7。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Brito JM、Teillet MA、Le Douarin NM。 来自前肠内胚层的声波刺猬在颌骨发育中的早期作用:确保神经嵴细胞存活。 美国国家科学院院刊2006; 103(31):11607–12. doi:10.1073/pnas.0604751103。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Bulgakov OV、Eggenschwiler JT、Hong DH、Anderson KV、Li T.FKBP8是神经组织中小鼠声音刺猬信号的负调节因子。 发展。 2004; 131(9):2149–59. doi:10.1242/dev.01122。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Buttita L,Mo R,Hui CC,Fan CM。Gli2和Gli3的相互作用及其在介导Shh依赖性硬结瘤诱导中的需求。 发展。 2003; 130(25):6233–43. doi:10.1242/dev.00851。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Camner P、Mossberg B、Afzelius BA。两名受试者气管支气管内先天性无功能纤毛的证据。 美国Rev Respir Dis。 1975; 112(6):807–9. doi:10.1164/ard.1975.112.6.807。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Cano DA、Murcia NS、Pazour GJ、Hebrok M.Orpk多囊肾病小鼠模型揭示了初级纤毛在胰腺组织组织中的重要作用。 发展。 2004; 131(14):3457–67. doi:10.1242/dev.01189。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Chazaud C,Rossant J.Apc突变体早期近端模式和AVE形成的破坏。 发展。 2006; 133(17):3379–87. doi:10.1242/dev.02523。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Chiang C,Litingtong Y,Lee E,Young KE,Corden JL,等。声波刺猬基因功能缺失小鼠的近视和轴型缺陷。 自然。 1996; 383(6599):407–13. doi:10.1038/383407a0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ciruna B、Weidinger G、Knaut H、Thisse B、ThisseC等。通过生殖系替换生产母体合子突变斑马鱼。 美国国家科学院院刊,2002年; 99(23):14919–24. doi:10.1073/pnas.222459999。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Cole DG、Diener DR、Himelblau AL、Beech PL、Fuster JC、Rosenbaum JL。 衣原体驱动蛋白Ⅱ依赖的鞭毛内转运(IFT):IFT颗粒含有秀丽隐杆线虫感觉神经元中纤毛组装所需的蛋白质。 细胞生物学杂志。 1998; 141(4):993–1008. doi:10.1083/jcb.141.4.993。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Corbit KC、Aanstad P、Singla V、Norman AR、Stainier DY、Reiter JF。脊椎动物初级纤毛的平滑功能。 自然。 2005; 437(7061):1018–21. doi:10.1038/nature04117。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
DasGupta R,Fuchs E.毛囊发育和分化过程中激活的LEF/TCF转录复合物的多重作用。 发展。 1999; 126(20):4557–68. doi:10.1242/dev.126.20.4557。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Dawe HR、Smith UM、Cullinane AR、Gerrelli D、Cox P等。Meckel-Gruber综合征蛋白MKS1和meckelin相互作用,是初级纤毛形成所必需的。 人类分子遗传学。 2007; 16(2):173–86. doi:10.1093/hmg/ddl459。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Dutcher SK.Flagellar大会分为250个简单易行的步骤。 趋势Genet。 1995; 11(10):398–404. doi:10.1016/s0168-9525(00)89123-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Eggenschwiler JT,Espinoza E,Anderson KV。Rab23是小鼠声波刺猬信号通路的重要负调控因子。 自然。 2001; 412(6843):194–8. doi:10.1038/35084089。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Fan S,Hurd TW,Liu CJ,Straight SW,Weimbs T,等。极性蛋白通过驱动蛋白-运动相互作用控制纤毛发生。 当前生物量。 2004; 14(16):1451–61. doi:10.1016/j.cub.2004.08.025。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ferrante MI、Zullo A、Barra A、Bimonte S、Messaddeq N等。初级纤毛形成和左右轴规范需要口腔面部数字I型蛋白。 自然遗传学。 2006; 38(1):112–7. doi:10.1038/ng1684。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Fischer JA、Eun SH、Doolan BT。内吞作用、内体贩运和果蝇发育的调控。 年收入细胞开发生物。 2006; 22:181–206. doi:10.1146/annurev.cellbio.22.010605.093205。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Follit JA、Tuft RA、Fogarty KE、Pazour GJ。 鞭毛内转运蛋白IFT20与高尔基复合体相关,是纤毛组装所必需的。 分子生物学细胞。 2006; 17(9):3781–92. doi:10.1091/mbc。 E06-02-0133。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Forsberg Nilsson K,Behar TN,Afrakhte M,Barker JL,McKay RD。血小板衍生生长因子诱导神经上皮干细胞的趋化性。 1998年神经科学研究杂志; 53(5):521–30. doi:10.1002/(SICI)1097-4547(19980901)53:5<521::AID-JNR2>3.0.CO; 2-B类。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Franco B,Ballabio A.X失活与人类疾病:X连锁显性男性致死性疾病。 2006年通用操作基因开发; 16(3):254–9. doi:10.1016/j.gde.2006.04.012。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Galceran J、Farinas I、Depew MJ、Clevers H、Grosschedl R.Wnt3a−/−样表型和Lef1(−/-)Tcf1(−/-)小鼠的肢体缺陷。 基因开发1999; 13(6):709–17. doi:10.1101/gad.13.6.709。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Greenberg ME、Ziff EB。 刺激3T3细胞可诱导c-fos原癌基因的转录。 自然。 1984; 311(5985):433–8. doi:10.1038/311433a0。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Guo N、Hawkins C、Nathans J.Frizzled6控制小鼠的毛发模式。 美国国家科学院院刊2004; 101(25):9277–81. doi:10.1073/pnas.0402802101。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Hagstrom SA、Adamian M、Scimeca M、Pawlyk BS、Yue G、Li T。试管样蛋白1在视紫红质转运中的作用。 投资眼科视觉科学。 2001; 42(9):1955–62. [ 公共医学 ] [ 谷歌学者 ] -
韩英光、郭博华、柯南MJ。 果蝇需要鞭毛内运输来区分感觉纤毛,而不是精子。 当前生物量。 2003; 13(19):1679–86. doi:10.1016/j.cub.2003.08.034。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Haycraft CJ、Banizs B、Aydin-Son Y、Zhang Q、Michaud EJ、Yoder BK。Gli2和Gli3定位于纤毛,需要鞭毛内转运蛋白极化子进行加工和功能。 公共科学图书馆-遗传学。 2005; 1(4):e53。 doi:10.1371/journal.pgen.0010053。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Haycraft CJ、Zhang Q、Song B、Jackson WS、Detloff PJ等。鞭毛内运输对软骨内骨形成至关重要。 发展。 2007; 134(2):307–16. doi:10.1242/dev.02732。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Hearn T、Spalluto C、Phillips VJ、Renforth GL、Copin N等。ALMS1的亚细胞定位支持中心体和基底体功能障碍参与肥胖、胰岛素抵抗和2型糖尿病的发病机制。 糖尿病。 2005; 54(5):1581–7. doi:10.2337/糖尿病.54.51581。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Heldin CH,Westermark B.血小板衍生生长因子的作用机制和体内作用。 《生理学评论》,1999年; 79(4):1283–316. doi:10.1152/physrev.1999.79.4.1283。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Hoeller D,Volarevic S,Dikic I.生长因子受体信号的分区。 当前操作细胞生物学。 2005; 17(2):107–11. doi:10.1016/j.ceb.2005.01.001。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Hu MC,Mo R,Bhella S,Wilson CW,Chuang PT,等。Gli1,Gli2和肾脏模式基因的GLI3依赖性转录抑制破坏肾脏形态发生。 发展。 2006; 133(3):569–78. doi:10.1242/dev.02220。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Huangfu D,Liu A,Rakeman AS,Murcia NS,Niswander L,Anderson KV。小鼠的刺猬信号需要鞭毛内转运蛋白。 自然。 2003; 426(6962):83–7. doi:10.1038/nature02061。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
皇甫D,安德森KV.Cilia和刺猬在小鼠中的反应性。 美国国家科学院院刊2005; 102(32):11325–30. doi:10.1073/pnas.0505328102。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Huangfu D,Anderson KV。从Smo到Ci/Gli的信号传递:从果蝇到脊椎动物的刺猬途径的保存和分化。 发展。 2006; 133(1):3–14. doi:10.1242/dev.02169。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Hui CC,Joyner AL.灰色头多指综合征小鼠模型:顶外J突变包含Gli3基因的基因内缺失。 自然遗传学。 1993; 3(3):241–6. doi:10.1038/ng0393-241。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ingham PW,美联社麦克马洪。 动物发育中的刺猬信号:范式和原理。 基因开发2001; 15(23):3059–87. doi:10.1101/gad.938601。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Izraeli S、Lowe LA、Bertness VL、Campaner S、Hahn H等。Sonic Hedgehog反应途径需要Sil的遗传证据。 起源。 2001; 31(2):72–7. doi:10.1002/gene.10004。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Karcher C、Fischer A、Schweickert A、Bitzer E、Horie S等。Pkd1基因敲除胚胎中缺乏侧化表型与结纤毛中缺乏多囊蛋白-1相关。 区别。 2005; 73(8):425–32. doi:10.1111/j.1432-0436.2005.00048.x。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Kartagener M.Zur《支气管哮喘病原体:支气管哮喘bei Situs viscerum inversus》。 Beiträge zum Klinik der Tuberkulose公司。 1933; 83:489–501. [ 谷歌学者 ] -
Kim JC、Badano JL、Sibold S、Esmail MA、Hill J等。Bardet-Biedl蛋白BBS4靶向中心粒周围区域的货物,是微管锚定和细胞周期进展所必需的。 自然遗传学。 2004; 36(5):462–70. doi:10.1038/ng1352。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Kim JC、Ou YY、Badano JL、Esmail MA、Leitch CC等。MKKS/BBS6是一种与肥胖障碍Bardet-Biedl综合征相关的发散性伴侣蛋白样蛋白,是胞质分裂所需的一种新型中心体成分。 细胞科学杂志。 2005; 118(第5部分):1007–20。 doi:10.1242/jcs.01676。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Kimelman D,Xu W.β-catenin破坏复合体:从结构角度的见解和问题。 致癌物。 2006; 25(57):7482–91. doi:10.1038/sj.onc.1210055。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Klinghoffer RA、Hamilton TG、Hoch R、Soriano P。PDGFalphaR基因座的等位序列表明不同信号通路在发育过程中的作用不同。 开发单元。 2002; 2(1):103–13. doi:10.1016/s1534-5807(01)00103-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Kogerman P、Grimm T、Kogerman L、Krause D、Unden AB等。哺乳动物抑制因子调节Gli-1的核质穿梭。 自然细胞生物学。 1999; 1(5):312–9. doi:10.1038/13031。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Kozminski KG、Johnson KA、Forscher P、Rosenbaum JL。 真核生物鞭毛中与鞭毛跳动无关的运动。 美国国家科学院院刊1993; 90(12):5519–23. doi:10.1073/pnas.90.12.5519。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Kozminski KG,Beech PL,Rosenbaum JL。 衣原体驱动蛋白样蛋白FLA10参与与鞭毛膜相关的运动。 细胞生物学杂志。 1995; 131(第6部分第1):1517–27。 doi:10.1083/jcb.131.61517。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Lei Q,Jeong Y,Misra K,Li S,Zelman AK,等。Wnt信号抑制剂调节神经管中对形态发生Shh-Gli信号的转录反应。 开发单元。 2006; 11(3):325–37. doi:10.1016/j.devcel.2006.06.013。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Li JB、Gerdes JM、Haycraft CJ、Fan Y、Teslovich TM等。比较基因组学确定了包含BBS5人类疾病基因的鞭毛和基底体蛋白质组。 单元格。 2004; 117(4):541–52. doi:10.1016/s0092-8674(04)00450-7。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Lin F、Hiesberger T、Cordes K、Sinclair AM、Goldstein LS等。驱动蛋白II的KIF3A亚基的肾特异性失活抑制肾纤毛生成并产生多囊肾病。 美国国家科学院院刊,2003年; 100(9):5286–91. doi:10.1073/pnas.0836980100。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Litingtong Y、Dahn RD、Li Y、Fallon JF、Chiang C.Shh和Gli3对于肢体骨骼的形成是不可或缺的,但可以调节数字和身份。 自然。 2002; 418(6901):979–83. doi:10.1038/nature01033。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Liu P、Wakamiya M、Shea MJ、Albrecht U、Behringer RR、Bradley A.脊椎动物轴形成中Wnt3的要求。 自然遗传学。 1999; 22(4):361–5. doi:10.1038/11932。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Liu A,Wang B,Niswander LA。小鼠鞭毛内转运蛋白调节Gli转录因子的激活和抑制功能。 发展。 2005; 132(13):3103–11. doi:10.1242/dev.01894。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
宾夕法尼亚州比奇Lum L,《刺猬反应网络:传感器、交换机和路由器》。 科学。 2004; 304(5678):1755–9. doi:10.1126/science.1098020。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Mak HY、Nelson LS、Basson M、Johnson CD、Ruvkun G.秀丽隐杆线虫脂肪储存的多基因控制。 自然遗传学。 2006; 38(3):363–8. doi:10.1038/ng1739。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Marszalek JR,Ruiz Lozano P,Roberts E,Chien KR,Goldstein LS。缺乏驱动蛋白II的KIF3A亚基的小鼠突变体中的倒位和胚胎纤毛形态发生缺陷。 美国国家科学院院刊1999; 96(9):5043–8. doi:10.1073/pnas.96.9.5043。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
May SR,Ashique AM,Karlen M,Wang B,Shen Y,等。IFT逆行运动的缺失会干扰Smo在纤毛上的定位,并阻止Gli激活和抑制功能的表达。 开发生物。 2005; 287(2):378–89. doi:10.1016/j.ydbio.2005.08.050。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
McMahon AP、Ingham PW、Tabin CJ。 刺猬信号的发育作用和临床意义。 当前最高开发生物。 2003; 53:1–114. doi:10.1016/s0070-2153(03)53002-2。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
McGrath J,Somlo S,Makova S,Tian X,Brueckner M。两个单眼节群在小鼠中引发左右不对称。 单元格。 2003; 114(1):61–73. doi:10.1016/s0092-8674(03)00511-7。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Meguid MM、Fetissov SO、Varma M、Sato T、Zhang L等。下丘脑多巴胺和血清素在食物摄入调节中的作用。 营养。 2000; 16(10):843–57. doi:10.1016/s0899-9007(00)00449-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Methot N,Basler K。融合抑制因子通过阻止Cubitus interrutus激活物形式的核积累来对抗刺猬信号转导。 发展。 2000; 127(18):4001–10. doi:10.1242/dev.127.18.4001。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Mo R、Freer AM、Zinyk DL、Crackower MA、Michaud J等。Gli2和Gli3锌指基因在骨骼模式形成和发育中的特定和冗余功能。 发展。 1997; 124(1):113–23. doi:10.1242/dev.124.1.113。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Montcouquiol M,Sans N,Huss D,Kach J,Dickman JD等。Vangl2和Fz3的不对称定位表明哺乳动物平面细胞极性的新机制。 神经科学杂志。 2006; 26(19):5265–75. doi:10.1523/JNEUROSCI.4680-05.2006。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Murcia NS、Richards WG、Yoder BK、Mucenski ML、Dunlap JR、Woychik RP。左右轴测定需要橡树岭多囊肾(orpk)疾病基因。 发展。 2000; 127(11):2347–55. doi:10.12242/周五.127.11.2347。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Nauli SM,Zhou J.多囊藻毒素与肾纤毛和结纤毛的机械化。 生物论文。 2004; 26(8):844–56. doi:10.1002/bies.20069。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Nishimura DY、Fath M、Mullins RF、Searby C、Andrews M等。Bbs2-null小鼠存在神经感觉缺陷、社会支配缺陷以及与视紫红质定位错误相关的视网膜病变。 美国国家科学院院刊2004; 101(47):16588–93. doi:10.1073/pnas.0405496101。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Nonaka S、Tanaka Y、Okada Y、Takeda S、Harada A等。在缺乏KIF3B运动蛋白的小鼠中,由于节纤毛丢失导致胚胎外液向左流动而导致左右不对称的随机化。 单元格。 1998; 95(6):829–37. doi:10.1016/s0092-8674(00)81705-5。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Nonaka S、Yoshiba S、Watanabe D、Ikeuchi S、Goto T等。通过节纤毛后倾重新形成左右不对称。 《公共科学图书馆·生物》。 2005; 3(8):e268。 doi:10.1371/journal.pbio.0030268。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Nusse R.Wnts和Hedgehogs:脂质修饰的蛋白质和细胞表面信号机制的相似性。 发展。 2003; 130(22):5297–305. doi:10.1242/dev.00821。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ohlemiller KK、Hughes RM、Mosinger-Ogilvie J、Speck JD、Grosof DH、Silverman MS。管状小鼠的耳蜗和视网膜退化。 神经报告。 1995; 6(6):845–9. doi:10.1097/00001756-199504190-00005。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Omori Y、Malicki J.oko meduzy和相关碎屑基因是脊椎动物胚胎顶端细胞特征的决定因素。 当前生物量。 2006; 16(10):945–57. doi:10.1016/j.cub.2006.03.058。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Orentas DM、Hayes JE、Dyer KL、Miller RH。脊髓少突胶质细胞前体出现时需要声波刺猬信号。 发展。 1999; 126(11):2419–29. doi:10.1242/dev.126.11.2419。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Orozco JT、Wedaman KP、Signor D、Brown H、Rose L、Scholey JM。沿纤毛运动的电机和货物。 自然。 1999; 398(6729):674. doi:10.1038/19448。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ou G、Blacque OE、Snow JJ、Leroux MR、Scholey JM。鞭毛内运输马达的功能协调。 自然。 2005; 436(7050):583–7. doi:10.1038/nature03818。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Paces-Fessy M、Boucher D、Petit E、Paute-Briand S、Blanchet-Tournier MF。Gli的负调控因子,即融合抑制因子(Sufu),与SAP18、半乳糖凝集素3和其他核蛋白相互作用。 生物化学杂志2004; 378(第2部分):353–62。 doi:10.1042/BJ20030786。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Pan Y、Bai CB、Joyner AL、Wang B.Hedgehog信号通过抑制Gli2的加工和降解来调节Gli2转录活性。 分子细胞生物学。 2006; 26(9):3365–77. doi:10.1128/MCB.26.9.3365-3377.2006。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Pan X,Ou G,Civelekoglu-Scholey G,Blacque OE,Endres NF,等。驱动蛋白II和OSM-3马达协同作用下秀丽线虫纤毛中IFT颗粒的转运机制。 细胞生物学杂志。 2006; 174:1035–45. doi:10.1083/jcb.200606003。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Park TJ、Haigo SL、Wallingford JB。 缺乏内翻或模糊功能的胚胎中的纤毛发生缺陷与平面细胞极性和Hedgehog信号的失败有关。 自然遗传学。 2006; 38(3):303–11. doi:10.1038/ng1753。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Pazour GJ、Dickert BL、Vucica Y、Seeley ES、Rosenbaum JL等。衣原体IFT88及其小鼠同源物多囊肾病基因tg737是纤毛和鞭毛组装所必需的。 细胞生物学杂志。 2000; 151(3):709–18. doi:10.1083/jcb.151.3.709。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
佩德森LB、盖默S、罗森鲍姆JL。 衣原体鞭毛内转运的分子机制剖析。 当前生物量。 2006; 16(5):450–9. doi:10.1016/j.cub.2006.02.020。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Perkins LA、Hedgecock EM、Thomson JN、Culotti JG。 秀丽隐杆线虫的突变感觉纤毛。 开发生物。 1986; 117(2):456–87. doi:10.1016/0012-1606(86)90314-3。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Pietras K、Sjoblom T、Rubin K、Heldin CH、Ostman A.PDGF受体作为癌症药物靶点。 癌细胞。 2003; 3(5):439–43. doi:10.1016/s1535-6108(03)00089-8。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Piontek KB、Huso DL、Grinberg A、Liu L、Bedja D等。体内可条件灭活的Pkd1功能性漂浮等位基因。 《美国肾脏病杂志》。 2004; 15(12):3035–43. doi:10.1097/01.ASN.0000144204.01352.86。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Piperno G、Siuda E、Henderson S、Segil M、Vaananen H、Sassaroli M。逆行鞭毛内转运(IFT)的不同突变体具有相似的形态和分子缺陷。 细胞生物学杂志。 1998; 143(6):1591–601. doi:10.1083/jcb.143.61591。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Pledger WJ、Hart CA、Locatell KL、Scher CD。血小板衍生生长因子调节蛋白:转化细胞系的组成合成。 美国国家科学院院刊1981; 78(7):4358–62. doi:10.1073/pnas.78.7.4358。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Romio L、Fry AM、Winyard PJ、Malcolm S、Woolf AS、Feather SA。OFD1是一种中心体/基础体蛋白,在人类肾脏发生的间质-上皮转变过程中表达。 《美国肾脏病杂志》。 2004; 15(10):2556–68. doi:10.1097/01.ASN.0000140220.46477.5C。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Rosenbaum JL、Witman GB。 鞭毛内运输。 Nat Rev Mol细胞生物学。 2002年11月; 3(11):813–25. doi:10.1038/nrm952。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Ross AJ、May-Simera H、Eichers ER、Kai M、Hill J等。Bardet-Biedl综合征纤毛蛋白的破坏扰乱脊椎动物的平面细胞极性。 自然遗传学。 2005; 37(10):1135–40. doi:10.1038/ng1644。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Rubin LL,de Sauvage FJ。针对癌症中的Hedgehog途径。 Nat Rev药物发现。 2006; 5(12):1026–33. doi:10.1038/nrd2086。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Saadi Kheddouci S、Berrebi D、Romagnolo B、Cluzeaud F、Peuchmaur M等。表达β-连环蛋白基因激活突变体的转基因小鼠中多囊肾病的早期发展。 致癌物。 2001; 20(42):5972–81. doi:10.1038/sj.onc.1204825。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Santagata S、Boggon TJ、Baird CL、Gomez CA、Zhao J等。通过管状蛋白的G蛋白信号传导。 科学。 2001; 292(5524):2041–50. doi:10.1126/science.1061233。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Schneider L、Clement CA、Teilmann SC、Pazour GJ、Hoffmann EK等。PDGFRalphaalpha信号通过成纤维细胞中的初级纤毛进行调节。 当前生物量。 2005; 15(20):1861–6. doi:10.1016/j.cub.2005.09.012。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Sekimizu K、Nishioka N、Sasaki H、Takeda H、Karlstrom RO、Kawakami A。斑马鱼鬣蜥基因座编码Dzip1,这是一种新的锌指蛋白,是正确调节刺猬信号所需的。 发展。 2004; 131(11):2521–32. doi:10.1242/dev.01059。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Shiratori H,Hamada H。小鼠的左右轴:从起源到形态学。 发展。 2006; 133(11):2095–104. doi:10.1242/dev.02384。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Simons M、Gloy J、Ganner A、Bullerkotte A、Bashkurov M等。Inversin,一种在II型肾病综合征中突变的基因产物,起到Wnt信号通路之间的分子开关的作用。 自然遗传学。 2005; 37(5):537–43. doi:10.1038/ng1552。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Singla V,Reiter JF。作为细胞天线的初级纤毛:在感觉器官发出信号。 科学。 2006; 313(5787):629–33. doi:10.1126/science.1124534。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Soriano P.PDGFα受体是神经嵴细胞发育和体节正常模式所必需的。 发展。 1997; 124(14):2691–700. doi:10.1242/dev.124.14.2691。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Stiles CD、Capone GT、Scher CD、Antoniades HN、Van Wyk JJ、Pledger WJ。 生长激素和血小板衍生生长因子对细胞生长的双重控制。 美国国家科学院院刊1979; 76(3):1279–83. doi:10.1073/pnas.76.3.1279。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
St-Jacques B、Hammerschmidt M、McMahon AP。 印度刺猬信号调节软骨细胞的增殖和分化,对骨形成至关重要。 基因开发1999; 13(16):2072–86. doi:10.1101/gad.13.16.2072。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Sun Z、Amsterdam A、Pazour GJ、Cole DG、Miller MS、Hopkins N。斑马鱼的基因筛查确定纤毛基因是囊性肾的主要病因。 发展。 2004; 131(16):4085–93. doi:10.1242/dev.01240。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Supp DM、Brueckner M、Kuehn MR、Witte DP、Lowe LA等。靶向删除左右动力蛋白的ATP结合域证实了其在左右不对称发展中的作用。 发展。 1999; 126(23):5495–504. doi:10.1242/dev.126.23.5495。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Svard J、Heby-Henricson K、Persson-Lek M、Rozell B、Lauth M等。融合抑制因子的基因消除揭示了哺乳动物刺猬信号通路中的重要阻遏物功能。 开发单元。 2006; 10(2):187–97. doi:10.1016/j.devcel.2005.12.013。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
武田S、Yonekawa Y、Tanaka Y、Okada Y、Nonaka S、Hirokawa N。左右不对称性和驱动蛋白超家族蛋白KIF3A:通过KIF3A−/−小鼠分析确定偏侧性和中胚层诱导的新见解。 细胞生物学杂志。 1999; 145(4):825–36. doi:10.1083/jcb.145.4.825。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
te Welscher P、Zuniga A、Kuijper S、Drenth T、Goedemans HJ、Meijlink F、Zeller R。通过SHH介导的GLI3反作用促进脊椎动物肢体发育。 科学。 2002年10月25日; 298(5594):827–30. doi:10.1126/science.1075620。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Tempe D,Casas M,Karaz S,Blanchet Tournier MF,Concordet JP。多位点蛋白激酶A和糖原合成酶激酶3β磷酸化通过SCFbetaTrCP导致Gli3泛素化。 分子细胞生物学。 2006; 26(11):4316–26. doi:10.1128/MCB.02183-05。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
搔痒C.脊椎动物肢体上的数字图案。 Nat Rev Mol细胞生物学。 2006年1月; 7(1):45–53. doi:10.1038/nrm1830。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Tsujikawa M,Malicki J.鞭毛内转运基因对脊椎动物感觉神经元的分化和存活至关重要。 神经元。 2004; 42(5):703–16. doi:10.1016/s0896-6273(04)00268-5。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Tucker RW、Pardee AB、Fujiwara K.Centriole纤毛与3T3细胞的静止和DNA合成有关。 单元格。 1979年a; 17(3):527–35. doi:10.1016/0092-8674(79)90261-7。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Tucker RW,Scher CD,Stiles CD。与3T3细胞对生长因子的早期反应有关,但与SV40无关。 单元格。 1979年b; 18(4):1065–72. doi:10.1016/0092-8674(79)90219-8。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Uren A,Yu JC,Ghoma NS,Pierce JH,Heidaran MA。α-PDGFR酪氨酸激酶通过趋化性和趋化性介导两种不同细胞类型的运动。 生物化学与生物物理研究委员会。 1994; 204(2):628–34. doi:10.1006/bbrc.1994.2505。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Vieira AR、Avila JR、Daack-Hirsch S、Dragan E、Felix TM等。非综合征性唇腭裂候选基因的医学测序。 公共科学图书馆-遗传学。 2005; 1(6):e64。 doi:10.1371/journal.pgen.0010064。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Walther Z、Vashishtha M、Hall JL。 衣原体FLA10基因编码一种新的驱动蛋白同源蛋白。 细胞生物学杂志。 1994; 126(1):175–88. doi:10.1083/jcb.126.1.175。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Wang B,Fallon JF,Beachy PA。Hedgehog调节的Gli3加工在发育中的脊椎动物肢体中产生前/后阻遏物梯度。 单元格。 2000; 100(4):423–34. doi:10.1016/s0092-8674(00)80678-9。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Wang B,Li Y.直接参与的证据 {β}TrCP 在Gli3蛋白质加工中。 美国国家科学院院刊2006; 103(1):33–8. doi:10.1073/pnas.0509927103。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Wang Y,Nathans J.脊椎动物的组织/平面细胞极性:新见解和新问题。 发展。 2007; 134(4):647–58. doi:10.1242/dev.02772。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Watanabe D、Saijoh Y、Nonaka S、Sasaki G、Ikawa Y等。左右决定簇Inversin是单眼节和其他9+0纤毛的成分。 发展。 2003; 130(9):1725–34. doi:10.1242/dev.00407。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ] -
Wigley WC、Fabunmi RP、Lee MG、Marino CR、Muallem S等。蛋白酶体机械与中心体的动态关联。 细胞生物学杂志。 1999; 145(3):481–90. doi:10.1083/jcb.1453.481。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Wolff C、Roy S、Lewis KE、Schauerte H、Joerg Rauch G等。Iguana编码一种新的锌指蛋白,该蛋白具有对斑马鱼胚胎中Hedgehog信号转导至关重要的卷曲螺旋结构域。 基因开发2004; 18(13):1565–76. doi:10.1101/gad.296004。 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ] -
Zeng L、Fagotto F、Zhang T、Hsu W、Vasicek TJ等。小鼠融合基因座编码Axin,Axin是Wnt信号通路的抑制剂,可调节胚胎轴的形成。 单元格。 1997; 90(1):181–92. doi:10.1016/s0092-8674(00)80324-4。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]