摘要 自噬是真核细胞降解和再循环大分子和细胞器的主要分解代谢途径。 该途径在环境应激条件下、发育过程中和各种病理情况下被激活。 在这项研究中,我们描述了活性氧(ROS)作为信号分子在饥饿诱导的自噬中的作用。 我们发现饥饿刺激ROS的形成,特别是H 2 O(运行) 2 这些氧化条件对自噬至关重要,因为用抗氧化剂处理可以消除自噬体的形成和蛋白质的降解。 此外,我们确定半胱氨酸蛋白酶HsAtg4是H氧化的直接靶点 2 O(运行) 2 ,并指定位于HsAtg4催化位点附近的半胱氨酸残基为该法规的关键。 这种调节突变体的表达阻止了细胞内自噬体的形成,从而为自噬过程的氧化还原调节提供了分子机制。
介绍 自噬是将蛋白质和细胞器传递到溶酶体/液泡的主要途径,在那里它们被降解和再循环。 自噬在分化和发育以及细胞对应激的反应中起着重要作用。 它在氨基酸缺乏期间被激活,并与神经退行性疾病、癌症、病原体感染和肌病有关(参见 Cuervo,2004年 ; Shintani和Klonsky,2004年 ). 自噬是由新月形的隔离膜/吞噬体在细胞质成分周围启动的,形成一个封闭的双层膜结构,称为自噬体。 最后,自噬体与溶酶体融合成为自溶体,其内容物被溶酶体水解酶降解。
在过去的十年里,自噬的分子机制已经被广泛研究,参与这一过程的基因称为ATG( 一 u个 t吨 奥帕 克 y相关; 克林斯基 等 , 2003 ),被发现从酵母到人类都是保守的( Ohsumi,2001年 ; Huang和Klionsky,2002年 ). 然而,不同基因产物所起的作用及其作用方式有待解决。 自噬过程中的一个标志性事件是Atg8家族蛋白与自噬体膜的可逆结合( 市村真一 等 , 2000 ). 这种泛素样(UBL)蛋白家族与多种细胞过程有关,包括酵母中的Atg8p( 冗长的 等 , 1998 ; 市村真一 等 , 2000 )和GATE-16(16kDa高尔基相关ATP酶增强子; 萨吉夫 等 , 2000 ),LC3(轻链3; Mann和Hammarback,1994年 )GABARAP(GABA受体相关蛋白; 王 等 , 1999 )在哺乳动物中。 所有Atg8同源物(以下称为Atg8s)都是Atg4半胱氨酸蛋白酶家族的底物( 基里萨科 等 , 2000 ; 赫梅拉尔 等 , 2003 ; 马里诺 等 , 2003 ; 谐谑曲-舒瓦尔 等 , 2003 ). Atg4s在保守甘氨酸下游的C末端附近裂解Atg8s。 这种裂解允许Atg8通过暴露的甘氨酸与磷脂酰乙醇胺(PE)结合,这是一个通过泛素化样机制介导的过程。 Atg8-PE是Atg4的另一种底物,它将Atg8裂解并从膜中释放出来。 值得注意的是,这个共轭过程之前必须有另一个类似泛素的过程,将Atg12共轭到Atg5( 谷多 等 ,2004年b ). 在哺乳动物细胞中,氨基酸剥夺诱导LC3脂质化( 卡贝亚 等 , 2000 )以及GATE-16和GABARAP( 卡贝亚 等 , 2004 ). 脂溶性Atg8与吞噬细胞和自噬体结合,并一直存在,直到与溶酶体融合,此时自噬体内的Atg8可能被降解( 卡贝亚 等 , 2000 , 2004 ).
根据与酵母的序列同源性,已经报道了至少四种Atg4哺乳动物同源物 酿酒酵母 (Sc)附件4。 其中两个同源物HsAtg4A和HsAtg 4B能够以不同的效率切割三种哺乳动物的Atg8s:HsAttg4A主要切割GATE-16,而HsAttg 4B切割所有三个同源物(GATE-16、GABARAP和LC3),对LC3的效率最高( 卡贝亚 等 , 2004 ).
Atg4s既起结合酶的作用,也起去结合酶的作用,因此它们的活性预计会受到严格调控。 因此,在自噬过程中,在Atg8样蛋白的初始裂解后,Atg4必须变得不活跃,以确保Atg8与自噬体膜的结合。 随后,当自噬体与溶酶体融合时,Atg4可能会局部重新激活,以脱脂和再循环Atg8。 这项规定是如何实现的? 我们之前已经证明,重组HsAtg4A仅在还原剂DTT存在下作为GATE-16的半胱氨酸蛋白酶具有活性( 谐谑曲-舒瓦尔 等 , 2003 ). 这可能意味着, 体内 ,Atg4的脱脂活性是通过在不同条件和特定亚细胞微环境下发生的氧化还原电位的变化来调节的。 除其他因素外,活性氧(ROS)可能会造成这种调节。 高浓度的活性氧对细胞有害,导致程序性细胞死亡(PCD)( Jabs,1999年 ; 李 等 , 2003 ; Macip公司 等 , 2003 ). 然而,在低水平下,活性氧可以通过多种途径中的氧化因子作为信号分子,从而导致生长和存活。 在这里,我们首次报道ROS作为信号分子参与营养缺乏诱导的自噬,自噬本质上是一种生存途径。 我们发现,在饥饿条件下,细胞形成ROS,特别是H 2 O(运行) 2 对自噬体的形成和自噬降解至关重要。 氧化信号部分依赖于PI3K,导致Atg4的抑制。 使用 在体外 我们可以证明H 2 O(运行) 2 直接调节HsAtg4A。 此外,我们确定了Cys 81 位于活性位点下游的四个氨基酸,是HsAtg4A氧化还原调节的必需残基。 突变的HsAtg4A在Cys中的表达 81 或相应的HsAtg4B突变体,抑制细胞中GATE-16或LC3标记的自噬体的形成。
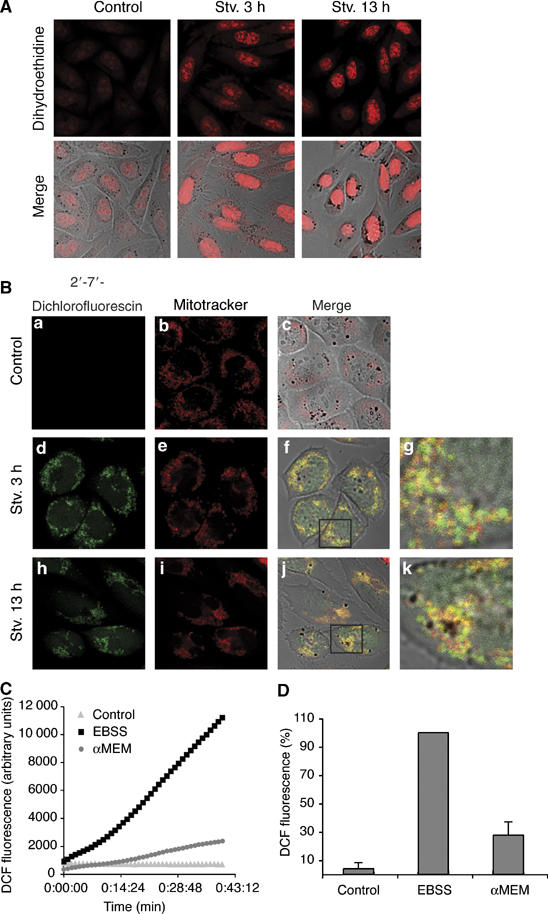
结果 饥饿诱导活性氧的形成 活性氧在多种细胞过程中充当信号分子,包括生长、分化、粘附和PCD。 为了测试活性氧是否在自噬中起作用,我们使用荧光探针二氢乙硫磷(DHE)测量了饥饿状态下活性氧的生成。 DHE与过氧化物反应形成一种化合物,与DNA结合后会发出荧光,可通过共焦荧光显微镜观察( 范登·霍克 等 , 1997 ). 事实上,在营养缺乏时,DHE染色很明显,表明细胞在这些条件下积累过氧化物( 图1A ). 然后我们使用另一个探针,2′,7′-二氯荧光素二乙酸酯(DCF-DA),它主要与H反应 2 O(运行) 2 在相互作用点形成荧光化合物( 卡思卡特 等 , 1983 ; 范登·霍克 等 , 1997 ). 饥饿诱导15分钟后DCF染色已明显( 图1Bd 和 补充图1A ,见下文)。 DCF染色的模式是点状的和细胞溶质的,在通宵饥饿后也很明显( 图1Bh ). 这些结果表明H 2 O(运行) 2 很可能是氧化剂在饥饿的细胞中积累。 为了排除与饥饿相关的氧化信号导致PCD的可能性,我们测试了在营养饥饿条件下生长的细胞的活性。 如所示 补充图1B 营养饥饿5h不影响细胞的存活率,对照组和饥饿组细胞的细胞死亡率同样低。 再加上发现在饥饿诱导数分钟后氧化信号出现,这些结果表明H 2 O(运行) 2 作为自噬途径中的信号分子,而不是导致PCD的氧化应激的一部分。
图1。
细胞在饥饿条件下积累ROS。 ( A类 )CHO细胞在对照培养基中生长(见材料和方法)或饥饿3或13小时,然后在50μM DHE中培养,并按照材料和方法中的详细说明进行可视化。 ( B类 )CHO细胞按照(A)中的方法生长,用5 nM MitoTracker Red在37°C下处理30分钟,清洗,用30μM DCFDA处理,并按照材料和方法中的详细说明进行可视化。 ( C类 )将生长在96周培养板中的HeLa细胞去除血清(αMEM)、完全饥饿(EBSS)或保存在对照培养基(α-MEM,10%FCS)中2 h,然后按照(B)中的方法用DCFDA处理细胞,随后在荧光仪中进行分析,如材料和方法中所述。 ( D类 )根据材料和方法中的详细说明,对从荧光测量中收集的数据进行了分析。
DCF染色的模式与线粒体染色相似,因此我们用DCF和功能线粒体标记MitoTracker Red对细胞进行双重染色。我们发现这两个标记之间存在显著重叠( 图1Bf和j ; 分别以g和k表示的放大)。 所有DCF点要么与MitoTracker染色结构共定位,要么靠近MitoTracker染色结构,表明线粒体是饥饿期间产生的ROS的来源。 这一发现与我们观察到的吞噬细胞一致,吞噬细胞通过Atg5和Atg16双重染色鉴定( 水岛 等 , 2003 ),倾向于与MitoTracker红色染色线粒体共定位( 补充图1C )表明饥饿时线粒体中产生的活性氧与自噬体形成之间存在功能联系。 该结果与最近报告的结果一致( 基索娃 等 , 2006 ).
为了更准确地评估活性氧的增加,我们使用设置为485 nm激发和535 nm发射的荧光计测量了两种不同细胞系(CHO和HeLa)中DCF荧光的增加。 细胞要么不含血清,要么完全饥饿(不含血清和营养物质),要么在对照培养基中生长2小时,然后用DCF和整个群体的荧光信号(3×10 4 细胞)。 如所示 图1C 与对照组相比,细胞完全饥饿导致荧光信号随时间急剧增加,而血清饥饿仅对荧光信号产生轻微影响,表明ROS生成略有增加。 进一步处理这些测量值,并将其总结为与试剂孵育20分钟后DCF荧光增加( 图1D ).
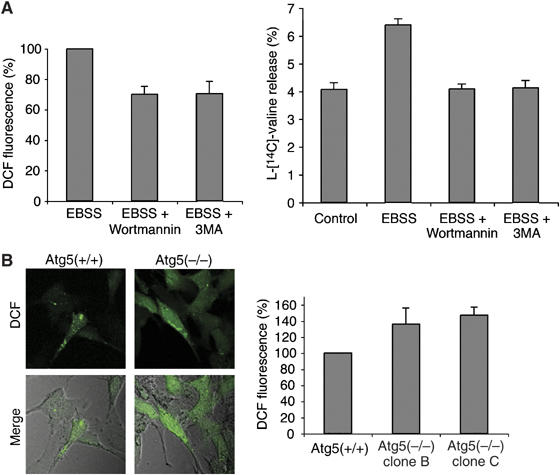
III类PI3K参与依赖于饥饿的ROS形成 为了了解ROS产生在自噬信号通路中的位置,我们监测了两种已知的自噬抑制剂沃特曼和3-甲基腺嘌呤(3MA)对饥饿依赖性DCF染色的影响( 珀蒂奥 等 , 2000 ). 在两种药物作用下,细胞饥饿2小时后,DCF荧光水平降低了25–30%( 图2A ,左侧面板)。 正如预期的那样,经过处理的细胞中的自噬活性通过长寿命蛋白质的降解速率进行测试(已知在自噬过程中会增加,参见 水岛 等 , 2001 ),显著减少( 图2A ,右侧面板)。 沃特曼和3MA是磷脂酰肌醇3激酶(PI3K)抑制剂。 III类PI3K通过与Beclin 1形成基本复合物,在哺乳动物自噬体形成的早期阶段发挥关键作用,因此,抑制其活性可抑制自噬过程( 珀蒂奥 等 , 2000 ; 塔萨 等 , 2003 ). 因此,这一发现表明活性氧的产生部分依赖于PI3K。 事实上,Beclin 1的部分沉默(先前显示会导致部分自噬缺陷; 暗礁 等 , 2006 )或PI3K hVps34导致饥饿诱导的活性氧生成分别减少15%和20%( 补充图2A ). 值得注意的是,hVps34的部分沉默导致每个细胞自噬体数量减少30%( 补充图2B ,右下面板)。
图2。
活性氧的形成部分发生在III类PI3K活化的下游。 ( A类 )左图:HeLa细胞在存在或不存在100 nM沃特曼或10 mM 3MA的情况下饥饿2小时,然后用DCFDA处理并用上述荧光仪进行分析。 右图:在不含或含有100 nM沃特曼或10 mM 3MA的α-MEM培养基或EBSS培养基中培养的细胞中测量长寿命蛋白质的降解率。 数值表示为三次单独测定的平均值±标准差。 ( B类 )来自两个单独克隆的WT MEF或Atg5(−/−)MEF在DCFDA治疗和可视化或荧光分析前饥饿2小时。
接下来,我们评估了因Atg5基因而缺失的细胞系中DCF染色的水平,Atg5的产物对自噬体的形成至关重要,作用于III类PI3K下游( 铃木 等 , 2001 ; 谷多 等 ,2004年b ). 与对照细胞相比,突变细胞中的DCF染色没有减少( 图2B ),但显著增加(40–50%),表明活性氧的产生发生在自噬体形成的Atg5依赖步骤的上游,并表明在缺乏功能性自噬途径的情况下,细胞积累活性氧。
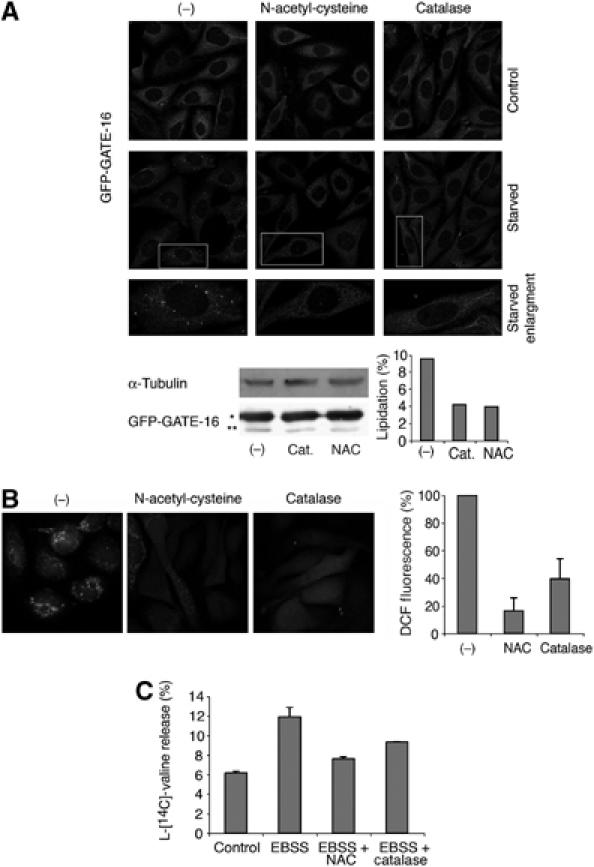
活性氧的形成对自噬至关重要 上述结果表明饥饿会诱导活性氧的形成。 但是活性氧对自噬至关重要吗? 为了解决这个问题,我们测试了 N个 -乙酰基- L(左) -半胱氨酸(NAC),一种通用抗氧化剂,和过氧化氢酶,它能特异性分解H 2 O(运行) 2 以GATE-16和LC3为标记物研究自噬体的形成。 LC3和GATE-16被证明在饥饿条件下标记自噬体( 卡贝亚 等 , 2004 ; 补充图3A ),如中所示 补充图3B ,这两种蛋白质在这些条件下表现出显著的共定位。 向生长培养基中添加NAC或过氧化氢酶( 樱井和Cederbaum,1998年 ; 普雷斯顿 等 , 2001 ; 徐 等 , 2003 )在2小时饥饿期前10分钟,GATE-16和LC3的脂质化减少,并消除了GATE-16-和LC3标记的自噬体的形成( 图3A 和 补充图3C ). 正如预期的那样,相同浓度的抗氧化试剂显著降低了处理细胞中的活性氧水平( 图3B ). 通过测试抗氧化剂对饥饿诱导的蛋白质降解的影响,进一步验证了这些结果( 图3C ). NAC处理对饥饿诱导的降解有60%的抑制作用,过氧化氢酶对饥饿诱导降解有25%的抑制作用。 综上所述,我们得出结论,饥饿条件会诱导活性氧的形成,而活性氧是诱导自噬所必需的。 值得注意的是,用H处理细胞 2 O(运行) 2 单独使用不会导致自噬体的形成(数据未显示)。
图3。
活性氧的积累对自噬至关重要。 ( A类 )上图:稳定转染GFP-GATE-16的CHO细胞在有或无10mM NAC或1000μ/ml过氧化氢酶的情况下预培养10分钟,然后在有或没有这些药物的情况下饥饿2小时,或在含有这些药物的对照培养基中生长2小时。然后固定细胞, 渗透并用抗GFP单克隆抗体染色。 显示了具有代表性的图像。 下面板:HEK 293细胞转染GFP-GATE-16。 在转染后24小时,用NAC或过氧化氢酶处理细胞并如上所述饥饿,在Ripa缓冲液中裂解,并在10%SDS-PAGE上分离100μg每种裂解产物,随后用抗GFP抗体进行分析,以检测转染的GATE-16和抗微管蛋白抗体作为对照。 使用NIH图像程序对数据进行量化,并将其描述为总GATE-16中脂蛋白的百分比。 ( * )表示非lipidated和( ** )表示固定的GFP-GATE-16。 ( B类 )用NAC或过氧化氢酶处理的CHO细胞,并按照(A)中的详细说明饥饿,用DCFDA孵育,用共焦显微镜观察或用荧光仪分析,如 图1 . ( C类 )在α-MEM培养基或EBSS培养基中培养的CHO细胞中,或在用10 mM NAC预处理10分钟或用1000μ/ml过氧化氢酶过夜后,测量长寿命蛋白质的降解率。 给出了一个具有代表性的实验。
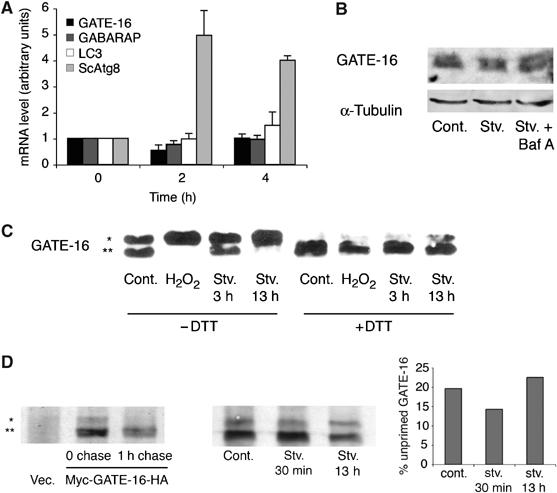
Atg4在饥饿时以氧化还原依赖的方式衰减 接下来,我们寻找饥饿诱导的氧化信号的细胞靶点。 饥饿期间,脂肪型Atg8s的数量显著增加( 卡贝亚 等 , 2004 ). 如所示 图4A ,在哺乳动物Atg8中,这种升高不是由转录增加引起的,如先前对ScAtg8所示( 基里萨科 等 , 1999 ). 一直以来,在这些条件下,我们无法观察到哺乳动物Atg8s蛋白水平的变化( 图4B 和数据未显示)。 因此,我们推断,脂肪化哺乳动物Atg8蛋白对营养饥饿的反应是由于加工酶的翻译后调节。 为了验证这一假设,将细胞饥饿3或13小时,然后对其进行裂解,并测试裂解产物对GATE-16的裂解能力。 在本试验中,重组GATE-16在两个末端标记(His 6 -GATE-16-HA),以便通过其在SDS–PAGE中更快的迁移率来检测裂解的GATE-16。 如所示 图4C 与对照细胞相比,将GATE-16孵育在由短时间饥饿3 h的细胞制备的裂解液中,导致切割活性降低40%。 长时间饥饿细胞13小时(在此期间细胞仍然存活)会进一步抑制卵裂活性。 然而,向裂解物中加入1mM DTT导致裂解活性的恢复,这表明Atg4的抑制是由蛋白质的氧化引起的。 始终用1 mM H处理细胞 2 O(运行) 2 与对照细胞相比,在裂解前1 h,Atg4的裂解活性受到强烈抑制。 向裂解液中添加DTT可恢复大部分失去的活性,证实氧化抑制是可逆的。 值得注意的是,GATE-16特异性蛋白酶HsAtg4A的蛋白质水平在任何测试条件下都没有变化( 补充图4A )支持Atg4活动的调节是翻译后的观点。 使用特异性抗HsAtg4A抗体来确认 图4C 由HsAtg4A介导(有关详细信息,请参阅 补充图4B )表明Atg4是饥饿状态下氧化还原调节的靶点,并表明饥饿状态下脂化Atg8的积累可能是由于抑制Atg4活性所致。 事实上,我们发现用1 mM H 2 O(运行) 2 导致脂质LC3积聚( 补充图4C ).
图4。
内源性Atg4的活性在营养饥饿时受到抑制。 ( A类 )CHO细胞(哺乳动物Atg8s)或 酿酒酵母 (对于ScAtg8)在饥饿培养基(分别为EBSS或SD-N)中培养不同的时间段,如材料和方法中所述,在使用Tri-reagent分离RNA之前所示。 对于每个基因,从非靶细胞获得的数据被设置为任意值1,相应饥饿细胞的结果被相应地归一化。 结果表示三个单独实验的平均值±标准差。 ( B类 )HEK 293细胞在对照培养基中生长,或在没有或存在100 nM巴非霉素A1的情况下在EBSS中饥饿2 h。 在Ripa缓冲液中获得的裂解液(100μg)在12%SDS-PAGE上运行,并使用抗GATE-16或抗管蛋白抗体进行Western blot分析。 ( C类 )CHO细胞在有或无1mM H的对照培养基中生长 2 O(运行) 2 1小时,或饥饿3或13小时。在Ripa缓冲液中获得的裂解产物(10μg)与重组His孵育 6 -GATE-16-HA(0.3μg)置于50 KT反应缓冲液(25 mM Tris,pH 7.4,50 mM KCl)中,在30°C下放置45分钟,存在或不存在1 mM DTT。 通过添加样品缓冲液和煮沸停止反应,然后在15%SDS-PAGE上解析样品,然后使用抗-His单克隆抗体进行Western blot分析。 实验重复六次; 显示了一个具有代表性的斑点。 ( D类 )左图:转染Myc-GATE-16-HA或空白载体作为对照的HeLa细胞被标记为[ 35 S] 蛋氨酸10分钟后立即溶解或在Ripa缓冲液中追赶1小时后溶解。 使用抗Myc抗体对裂解物进行免疫沉淀,并在15%SDS-PAGE上解析免疫沉淀。 中间面板:将转染Myc-GATE-16-HA的HeLa细胞保存在对照培养基中或在EBSS中饥饿30分钟或13小时,然后用[ 35 S] 蛋氨酸10分钟,立即裂解,SDS-PAGE分析,NIH图像程序定量(右侧)。 数值表示为总GATE-16中无质控形式的平均百分比。 ( * )在这幅图的所有部分中,都表示他的 6 -GATE-16-HA或Myc-GATE-16-HA和( ** )表示撕裂的His 6 -GATE-16或Myc-GATE-16。
Atg4在Atg8s的初始处理(启动)和脱脂步骤中都起作用。 先前表明,在富含营养的条件下,启动步骤在翻译后立即发生( 卡贝亚 等 , 2000 ). 为了测试这一步骤在饥饿期间是否也受到抑制,我们用Myc标记的GATE-16和C标记的HA转染细胞,使其饥饿30分钟(这段时间内预计会形成自噬体; 法西斯 等 , 2006 )或13小时,并对其进行[ 35 S] 材料和方法中详述的蛋氨酸脉冲追踪实验( 图4D ). 对照实验表明,脉冲标记后,转染细胞中立即检测到约19kDa的两条带( 图4D ,左侧面板)。 1小时追踪导致上带消失,表明该带对应于未启动的GATE-16(Myc-GATE-16-HA),而下带对应于启动的GATE-16(Myc-GATE-6)。 短暂的饥饿不会导致未致病菌的积累( 图4D ,中间面板),并且这种形式仅在长期饥饿13小时的细胞中积累。
这些结果表明,HsAtg4A在饥饿期间受到氧化还原调节。 由于启动活性不受短期饥饿的影响,我们得出结论,脱脂活性是该调控的主要目标。
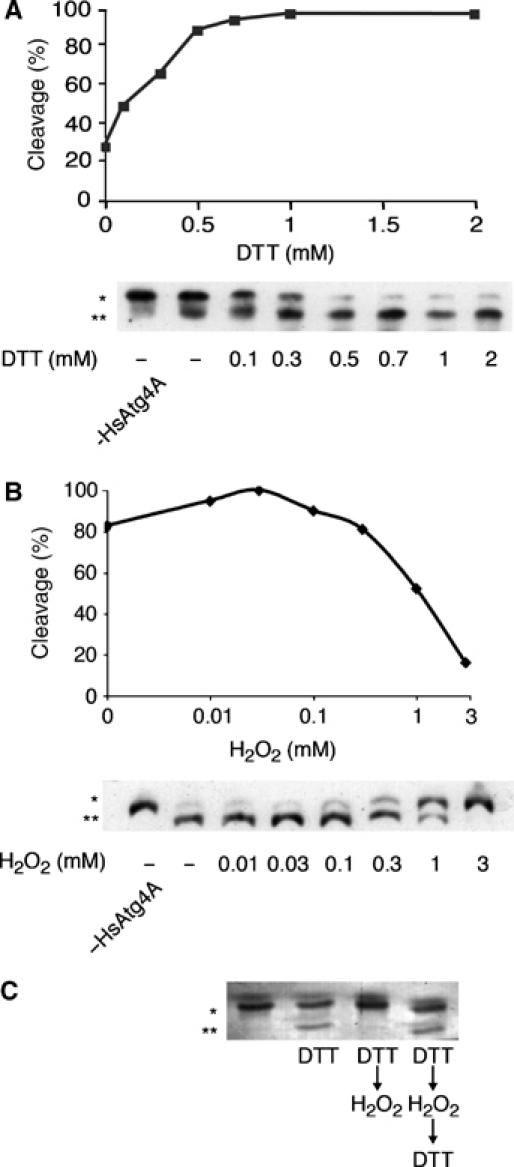
为了表征HsAtg4的氧化还原调节,我们将HsAtg 4A表达为重组蛋白,并在无细胞裂解实验中测试其活性。 如所示 图5A ,蛋白酶在还原反应混合物后逐渐被激活,并且在1mM DTT下活性达到饱和。 接下来,我们测试了H对HsAtg4A活性的抑制作用 2 O(运行) 2 蛋白质在低浓度DTT中预孵育以激活,然后添加H 2 O(运行) 2 与GATE-16孵育5分钟后进行卵裂试验。 HsAtg4A活性随着H 2 O(运行) 2 浓度增加到100μM以上( 图5B ). 在一些实验中,添加30μM H后,检测到蛋白酶活性略有增加 2 O(运行) 2 反应混合物。 最后,我们测试了H对HsAtg4抑制的可逆性 2 O(运行) 2 在本实验中,蛋白质在低浓度DTT中预孵育( 图5C ,车道2),然后添加H 2 O(运行) 2 (第3道),然后添加高浓度DTT(第4道;在添加下一种试剂之前,每个培养5分钟),然后用GATE-16培养。 在这些条件下,HsAtg4A完全恢复其活性。 综上所述,这些结果表明还原条件激活HsAtg4A,而氧化环境抑制其活性。 此外,这些结果表明H 2 O(运行) 2 单独使用足以可逆地抑制HsAtg4A的活性 在体外 ,因此建议H 2 O(运行) 2 也可能直接作用于蛋白酶 体内 .
图5。
H(H) 2 O(运行) 2 直接抑制HsAtgA的活性。 ( A类 )通过培养重组His来测试裂解活性 6 -HsAtg4A(0.1μg)和His 6 -GATE-16-HA(0.3μg)在50 KT反应缓冲液中,30°C,在指定浓度的DTT存在下放置45分钟,然后使用抗-His单克隆抗体进行Western blot分析。 使用Bio-Read Multi-Analyyst程序用密度计对三个单独实验产生的谱带进行量化,并将其表示为总GATE-16中裂解形式的平均百分比。 ( * )表示未裂开的His 6 -GATE-16-HA和( ** )表明劈开了他的 6 -门-16。 ( B类 )他的 6 -HsAtg4A在200μM DTT存在下于4°C孵育10分钟 6 -然后在50 KT中培养HsAtg4A(0.1μg)(以获得15μM DTT),H的指示浓度为 2 O(运行) 2 在25°C下保持5分钟,然后重组His 6 -添加GATE-16-HA(0.3μg),并在30°C下孵育45分钟。对三个单独实验的反应混合物进行了分析,并如(A)所述。 ( C类 )他的 6 -HsAtg4A(0.1μg)与重组His孵育 6 -GATE-16-HA(0.3μg),在以下程序后:不处理(1道); 在25°C下用200μM DTT预处理5分钟(第2车道); 用200μM DTT治疗,然后用1 mM H治疗 2 O(运行) 2 在25°C下保持5分钟(3号车道); 用200μM DTT治疗,然后用1 mM H治疗 2 O(运行) 2 持续5分钟,然后进行2 mM DTT(车道4)。 反应混合物用抗-His单克隆抗体进行Western blot分析。
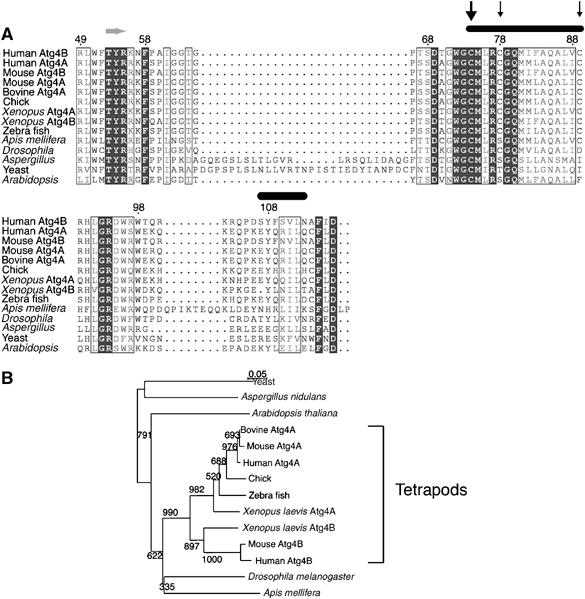
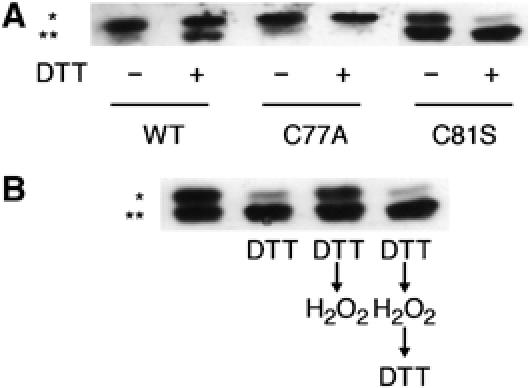
赛斯 81 是HsAtg4A氧化还原调节的靶点 HsAtg4A含有12个半胱氨酸,其中7个在Atg4A和Atg4B的四足同源物中高度保守。 后者相对于整个家庭来说具有较高的整体保护水平( 图6 ). 这些保守残基之一是蛋白酶Cys的假定活性残基 77 ( 基里萨科 等 , 2000 ; 谷多 等 ,2004年a ),酵母中唯一保存的半胱氨酸 酿酒酵母 ( 图6A ,大箭头,编号根据HsAtg4B)。 另一保守残基(Cys 81 HsAtg4A)位于活性残渣下游的四个位置。 我们首先试图确定Cys 77 是催化残渣,与ScAtg4和HsAtg4B同源。 如所示 图7A ,重组HsAtg4A,在77位携带半胱氨酸突变为丙氨酸(His 6 -HsHsAtg4A型 C77A型 )没有打开GATE-16 在体外 ,无论是在DTT缺席还是在场的情况下。 基于这个结果和上面描述的序列同源性,我们得出结论:Cys 77 是HsAtg4A活性位点的一部分。
图6。
Atg4的四足同源物共享几个保守的半胱氨酸残基。 ( A类 )Atg4家族成员中活性半胱氨酸残基周围区域的氨基酸排列。 以下登录号的序列对齐:BAB83890、BAB83889、NP_777364、NP_777363、NP_001001171、CAG32326、AAH73017、AAH82660、AAH95617、XP_393739、NP_608563、XP_661074、NP_014176和NP_191554。 使用ClustalX多重校准程序进行校准,并由ESScript 2.0程序描述,用黑色框架表示身份,用灰色背景表示同源性。 编号依据HsAtg4B序列。 根据最近解决的HsAtg4B结构,在该区域发现α-螺旋和β-片( 苏加瓦拉 等 , 2005 ; 库马诺米杜 等 , 2006 )分别标记为黑色椭圆和灰色箭头。 催化半胱氨酸残基用宽黑色箭头标记; 其他保守的半胱氨酸用黑色细箭头标记。 ( B类 )使用NJplot程序根据(A)中所示的比对创建了一个系统发育树( 佩里埃和古伊,1996年 ). 显示了指示树分支可靠性的引导值(最大值为1000)。
图7。
Cys81突变降低HsAtg4A的氧化还原敏感性 在体外 . ( A类 )重组His 6 -HsAtg4A型 重量 ,他的 6 -HsAtg4A型 C77A型 或他的 6 -HsAtg4A型 C81S公司 (0.1μg)与His孵育 6 -GATE-16-HA(0.3μg)在50 KT反应缓冲液中,30°C,在有或无1 mM DTT的条件下放置45分钟。 反应混合物用抗-His单克隆抗体进行Western blot分析。 ( * )表示没有劈开他的 6 -GATE-16-HA和( ** )表明劈开了他的 6 -大门-16。 ( B类 )他的 6 -HsAtg4A型 c81秒 (0.1μg)与重组His孵育 6 -GATE-16-HA(0.3μg)遵循与 图5C 对反应混合物进行了分析,并如(A)所述进行了介绍。
为了研究HsAtg4A氧化还原调节的机制,我们突变了保守的半胱氨酸残基81和92( 图6A ,小箭头),最接近活性残基,最接近丝氨酸(His 6 -HsAtg4A型 C81S公司 和他的 6 -HsAtg4A型 C92S型 并在不同氧化还原条件下对GATE-16的无细胞裂解实验中测试了它们的活性。 如所示 图7A ,Cys突变 81 显著降低HsAtg4A的氧化还原敏感性 6 -HsAtg4A型 C81S公司 即使在没有DTT的情况下也有部分活性。 胱氨酸突变 92 然而,对调节没有影响,蛋白酶在与His相同的条件下保持活性 6 -HsAtg4A型 重量 (未显示数据)。 接下来,我们测试了H氧化的效果 2 O(运行) 2 论他的活动 6 -HsAtg4A型 C81S公司 。我们发现突变蛋白在H存在下保持了显著活性 2 O(运行) 2 ( 图7B )在WT被完全抑制的条件下( 图5C ). 与WT类似,His的部分抑制 6 -HsAtg4A型 C81S公司 按H 2 O(运行) 2 可以通过添加DTT来逆转。 从这些结果中,我们得出结论:Cys 81 对HsAtg4A的氧化还原调节很重要 在体外 .
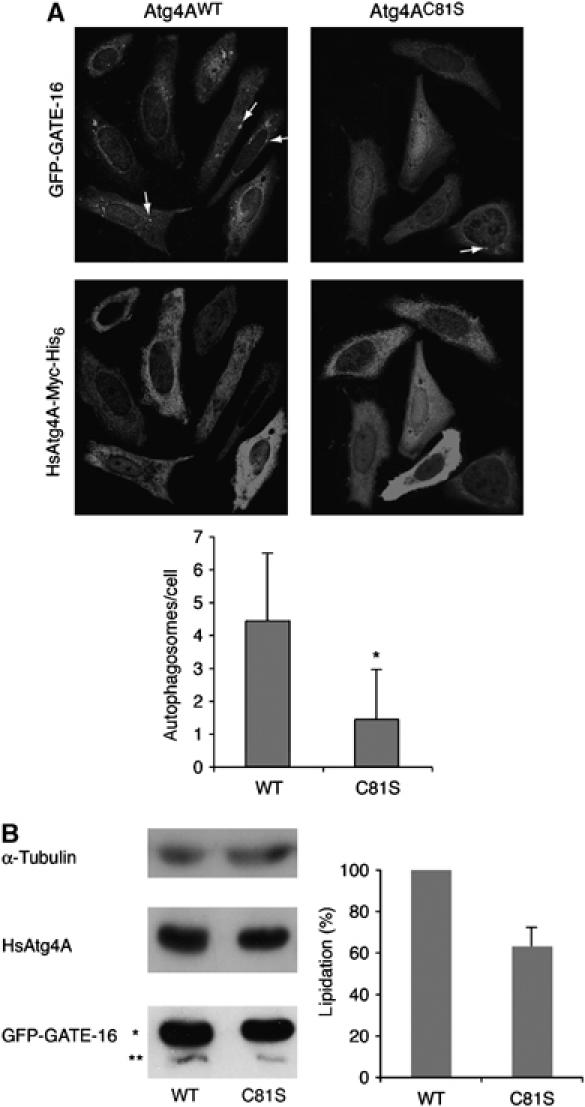
HsAtg4A和HsAtg 4B在自噬过程中均受氧化还原调节 检查Cys是否 81 在HsAtg4A的氧化还原调节中起作用 体内 ,我们监测了转染HsAtg4A突变体的细胞中GFP-GATE-16标记的自噬体的形成。 HsAtg4A的表达 C81S公司 严重损害饥饿细胞中GATE-16标记的自噬体的形成( 图8A )与表达HsAtg4A的细胞相比 重量 ( 图8A )或HsAtg4A C92S型 (未显示数据)处于类似水平。 HsAtg4A中形成的自噬体的平均数量 c81秒 -表达细胞比HsAtg4A低70% 重量 -表达细胞( 图8A ,下面板)。 有人可能认为,在HsAtg4A存在的情况下 C81S公司 ,自噬体形成,但随后迅速降解,因此无法看到。 为了排除这种可能性,我们使用巴非霉素A抑制溶酶体中自噬体的降解( 穆萨维 等 , 2001 ). 这种治疗没有影响突变表型,表明在转染HsAtg4A的细胞中不会形成自噬体 C81S公司 (未显示数据)。 GATE-16标记的自噬体在HsAtg4A中不形成的发现 C81S公司 -GATE-16脂质测定进一步支持了表达细胞。 作为HsAtg4A C81S公司 突变体对氧化不太敏感,我们预计与转染HsAtg4A的细胞相比,转染该突变体的饥饿细胞的脱脂活性更高 重量 事实上,我们发现在转染HsAtg4A的细胞中,GATE-16的脂化形式减少了近40% C81S公司 与转染HsAtg4A的细胞相比 重量 ( 图8B ). 值得一提的是,HsAtg4A的表达 C81S公司 正如预期的那样,HsAtg4A对GATE-16具有特异性,不处理LC3,因此不会影响细胞中LC3的染色模式(数据未显示)。 总之,我们的发现支持了Cys 81 在自噬过程中参与HsAtg4A的氧化还原调节。
图8。
Cys 81是细胞内HsAtg4A氧化还原调节所必需的。 ( A类 )用HsAtg4A瞬时转染稳定表达GFP-GATE-16的CHO细胞 重量 -Myc-His公司 6 或HsAtg4A C81S公司 -Myc-His公司 6 在转染后24小时,将细胞饥饿2.5小时,之后将其固定、透化并与抗Myc单克隆抗体孵育。 上面板:用共焦显微镜观察到的用上述构建物转染的细胞的典型图像; 典型的自噬体用箭头表示。 下面板:量化不同转染剂中每个细胞的平均自噬体数量。 使用尼康日食TE300荧光显微镜观察固定细胞的图像,并用于量化每个细胞的自噬体数量。 给出的结果是三个单独实验中总共100个细胞的平均值±标准差。 ( * )表示在 P(P) <0.001. ( B类 )左面板:HEK 293细胞与GFP-GATE-16和(A)中提到的每个Atg4A结构共同转染。 转染后40小时,将细胞饥饿2.5小时,在Ripa缓冲液中裂解,将100μg每种裂解物加载到10%SDS–PAGE中,随后用抗GFP抗体分析以检测转染的GATE-16,用抗Myc抗体检测转染的HsAtg4A,并用抗微管蛋白抗体作为对照。 ( * )表示非脂化GFP-GATE-16和( ** )表示固定的GFP-GATE-16。 右面板:使用NIH图像程序分析三个单独实验的结果,如左面板所示,并量化如下:计算每个实验中每个突变株GFP-GATE-16总GFP-GATE-16中的脂质量。 HsAtg4A的值 重量 在每个实验中,将其设定为100%,并相应计算转染突变体的细胞的相对脂质化。
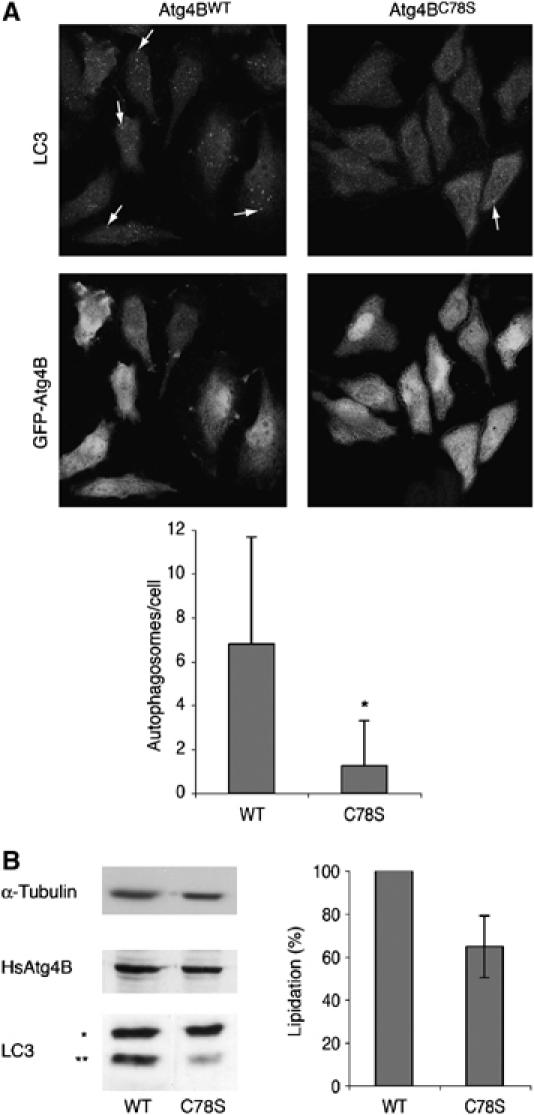
为了将我们的研究扩展到其他Atg4家族成员,我们决定测试LC3特异性蛋白酶HsAtg4B。 为此,我们用GFP-HsAtg4B转染细胞 重量 ,GFP-HsAtg4B C78S公司 或以GFP为对照,测试对LC3脂质化和LC3标记自噬体形成的影响。 我们发现GFP-HsAtg4B的过度表达 重量 和GFP-HsAtg4B C78S公司 减少LC3的自噬体形成和脂质化(数据未显示)。 这一现象以前曾因HsAtg4B的过度表达而被报道 重量 ( 谷多 等 ,2004年a ). 为了克服这个问题,我们添加了H 2 O(运行) 2 到饥饿介质。 在这些条件下,HsAtg4B 重量 被抑制,并观察到正常的自噬表型( 图9 ). HsAtg4B型 C78S公司 然而,对H有抵抗力 2 O(运行) 2 表达这种结构的细胞不能形成脂质LC3和LC3标记的自噬体。 这些结果表明,HsAtg4B与HsAtg 4A类似,是氧化还原调节的。
图9。
饥饿期间,HsAtg4B受到氧化还原调节,其机制与HsAt4A类似。 ( A类 )用GFP-HsAtg4B瞬时转染HeLa细胞 重量 或GFP-HsAtg4B C78S公司 在转染后24小时,细胞在4 mM h的存在下饥饿2.5小时 2 O(运行) 2 然后固定、渗透并与抗LC3多克隆抗体孵育。 如中所述,对细胞进行可视化(上面板)和量化(下面板) 图8 代表性的自噬体在上面板中用箭头表示。 下图中的结果是来自三个独立实验的总共100个细胞的平均值±s.d。 ( * )表示重要性 P(P) <0.001. ( B类 )左图:如(A)中所述转染和处理的HeLa细胞在Ripa缓冲液中裂解,将每种裂解物40μg加载到12%SDS-PAGE中,随后用抗LC3抗体和抗GFP抗体进行分析,以检测转染的HsAtg4B和抗微管蛋白抗体作为对照。 ( * )表示非脂化LC3和( ** )表示脂化LC3。 右侧面板:三个独立实验的结果,如左侧面板中所述,按中所述进行分析 图8B .
讨论 自噬是一种独特的细胞内贩运途径,通过细胞外信号激活。 虽然许多参与这一过程的蛋白质已经被鉴定,但导致自噬激活的信号转导途径尚未完全解决。 在这里,我们首次证明ROS作为信号分子参与饥饿诱导的自噬。 我们发现饥饿会触发活性氧的积累,很可能是H 2 O(运行) 2 ,这是自噬体形成和由此产生的降解途径所必需的。 氧化信号部分依赖于PI3K,位于Atg5依赖性结合机制的上游。 此外,我们确定了H氧化的直接目标 2 O(运行) 2 -蛋白酶Atg4。
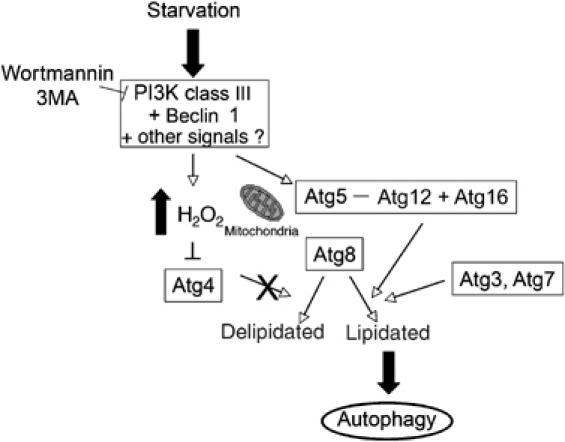
根据我们的结果,我们提出了以下模型,如所示 图10 :营养素缺乏导致III类PI3K和Beclin 1之间的复杂形成,这反过来与其他信号一起导致H的局部升高 2 O(运行) 2 线粒体附近。这种氧化信号导致自噬体形成部位的Atg4失活,从而促进Atg8的脂质化,这是自噬过程中的一个重要步骤。 随着自噬体成熟并与溶酶体融合,其定位明显改变为H 2 O(运行) 2 -环境恶劣,Atg4活跃,可以脱脂并回收Atg8。
图10。
自噬氧化还原调节的拟议模型。
H如何 2 O(运行) 2 管理Atg4? Atg4s是共享几个保守半胱氨酸残基的半胱氨酸蛋白酶。 其中之一是催化残留物,之前在Atg4家族的其他成员中发现,此处显示为Cys 77 在HsAtg4A中。 另一个保守基序是一个半胱氨酸残基,位于催化残基Cys的四个氨基酸中 81 在HsAtg4A中。 值得注意的是,我们发现该残基对HsAtg4A的氧化还原调节至关重要。 该残基突变为丝氨酸显著降低了蛋白质对H的敏感性 2 O(运行) 2 在体外 并阻断细胞内GATE-16标记的自噬体的形成。 通过发现相应的HsAtg4B突变体(HsAtg 4B),进一步证实了该残基对适当氧化还原调节的重要性 c78秒 )表现出类似的自噬缺陷。 我们的 在体外 研究表明H 2 O(运行) 2 直接使Atg4失活。 这可以通过绑定H来实现 2 O(运行) 2 至Cys 77 或至Cys 81 形成可逆的亚硫酸,保护Cys 77 或通过氧化导致其他修饰,如Cys之间形成二硫键 77 和Cys 81 ,再次屏蔽Cys 77 然而,在我们的实验中没有检测到这种二硫键(未公开的数据)。 或者,H 2 O(运行) 2 可能会影响其他站点,当Cys 81 是变异的。 活动站点Cys 77 ,从酵母到人类是保守的,而Cys 81 仅在Atg4A和Atg4B的四足同源物中保守。 低等物种在该残基中含有保守的Ser或Thr,因此选择了C81S突变体。 因此,我们认为氧化还原调节可能通过Cys介导 77 在低等物种中,而四足动物进化出一种更复杂的调节机制,需要两个半胱氨酸 77 和Cys 81 后者为四足动物提供了比低等物种更高的活性氧敏感性。 最近解决的HsAtg4B三维结构( 苏加瓦拉 等 , 2005 ; 库马诺米杜 等 , 2006 )支持我们的模型,如Cys所示 77 和Cys 81 位于α-螺旋线的起点,两者朝向螺旋线的同一平面。 由于两个半胱氨酸之间如此接近,Cys的修饰 81 确实会影响活动站点。 值得注意的是,这两种晶体都是在还原条件下制备的,因此没有关于任何半胱氨酸残基的二硫桥或其他修饰的报告。
半胱氨酸结合蛋白用作氧化还原传感器。 它们可以根据环境氧化条件的变化进行快速、可变、最重要的是可逆的翻译后修饰( Sitia和Molteni,2004年 ). 几种半胱氨酸蛋白酶,包括组织蛋白酶D、组织蛋白酶B和胞浆caspase-3( 钱德勒 等 , 1998 )和钙蛋白酶( 填料 等 , 2001 ),已经证明通过直接修饰活性位点进行氧化还原调节(参见 贾尔斯 等 , 2003 ). 作为信号通路的一部分,Atg4的氧化还原调节可以快速激活和灭活该蛋白酶,从而诱导自噬过程。
越来越多的证据表明,自噬在生存和死亡相关途径中发挥双重作用( Cuervo,2003年 ; Codogno和Meijer,2005年 ; 暗礁 等 , 2006 ). 迄今为止可用的数据将活性氧与自噬联系起来,描述了死亡相关途径,即II型PCD,这是由高水平的活性氧(即氧化应激; 于 等 , 2004 ; 基芬 等 , 2006 ). 最近,有研究表明,由于过氧化氢酶特异性自噬降解,半胱氨酸蛋白酶抑制通过自噬导致细胞死亡涉及活性氧的积累。 然而,在那项研究中,饥饿并没有产生类似的影响( 于 等 , 2006 ). 在我们的系统中,ROS的上升是局部的和可逆的; 它对细胞无害,可氧化特定靶点。 因此,我们的研究首次表明ROS作为生存途径中的信号分子参与饥饿诱导的自噬。
材料和方法 除非另有说明,否则结果部分中的所有实验都至少重复了三次。
重组蛋白的表达与纯化 重组His 6 -HsAtg4A(WT和中描述的突变体 补充数据 )和他的 6 -GATE-16-HA从pQE-30质粒中表达,并按前述方法纯化( 谐谑曲-舒瓦尔 等 , 2003 ). 他的净化 6 -HsAtg4A(WT和突变体)在10 mMβME存在下进行,以防止非特异性二硫键的形成; 然而,在使用前,蛋白质被稀释至至少30倍,或在缺乏βME的缓冲液中透析。
细胞培养、转染、试剂和免疫荧光 所有细胞系均在添加10%胎牛血清的α-MEM培养基(以色列生物工业公司)中生长,称为对照培养基。 Atg5−/−MEF和抗Atg16多克隆抗体是水岛博士(东京都市医学科学研究所)实验室赠送的礼物。 在1 mg/ml基因素(G418)中选择GFP-GATE-16-、GFP-LC3-或YFP-Atg5-转染CHO细胞的稳定克隆。 饥饿是在EBSS中进行的。 有关组织培养治疗中不同试剂的使用的详细信息,请参见 补充数据 根据制造商的方案,用脂质体(Invitrogen)对CHO细胞进行转染。 用标准磷酸钙技术转染HeLa和HEK 293细胞。 对于免疫荧光,除非另有说明,否则将细胞固定在甲醇中,用丙酮渗透,然后与所示抗体孵育,并用尼康日蚀TE300荧光显微镜或奥林巴斯IX-70共焦显微镜观察。
过氧化氢测量 H(H) 2 O(运行) 2 使用50μMDHE(分子探针)和30μM DCFDA(分子探针。 请参见 补充数据 了解详细信息。 荧光信号由奥林巴斯IX-70共焦显微镜检测。 每个实验至少重复三次,并显示代表性图像。 DCFDA信号也由SPECTRAmax gimini荧光仪测量,设置为485激发和535发射,并在37°C下保持40分钟。从荧光测量中收集的数据分析如 补充数据 .所有荧光测量的结果均为至少三次重复或三次实验的平均值±标准差。
蛋白质降解试验 根据以下描述的方案在CHO细胞上进行检测 法西斯 等 (2006) 必要时,添加10 mM 3MA、100 nM沃特曼、1000μ/ml过氧化氢酶或10 mM NAC。 值得注意的是,在饥饿诱导前加入NAC 10分钟,并在饥饿前14小时加入过氧化氢酶。 [ 14 C] 缬氨酸释放量计算为TCA可溶上清液中放射性相对于总细胞放射性的百分比。
细胞裂解物的制备 将细胞在Ripa缓冲液(0.1M NaCl、5mM EDTA、0.1M Na)中匀浆 2 高性能操作 4 /NaH(氢化钠) 2 PO4、pH 7.5、1%Triton、0.5%DOC、0.1%SDS和蛋白酶抑制剂)在冰上旋转。 匀浆在20000℃下离心15分钟 克 收集含有裂解物的上清液。
mRNA测量 RNA由Tri-reagent(Molecular Research Center Inc.)分离,详见 补充数据 .
脉冲追踪实验 用Myc-GATE-16-HA或空载体瞬时转染HeLa细胞。 然后将细胞保存在完全培养基中24 h,然后将其在无蛋氨酸培养基中培养10 min或在EBSS中饥饿30 min(2×10 cm平板)。 对于长时间饥饿,细胞(三个平板)在转染后10 h转移到EBSS中13 h,然后用新鲜EBSS替换培养基。 然后用0.2 mCi脉冲标记所有细胞[ 35 S] 蛋氨酸在37°C下放置10分钟,或在37°C下的全培养基中追逐1小时,或立即在冰上溶解,在补充了0.5 mM的Ripa缓冲液中 N个 -乙基马来酰亚胺(sigma)。 用抗Myc抗体对裂解液进行免疫沉淀,并在15%SDS-PAGE上进行分析。
致谢 我们感谢弗拉基米尔·基什(Vladimir Kish)对共焦显微镜的宝贵指导,感谢弗里达·西蒙(Frida Shimron)的全力协助。 我们也感谢Avihai Danon的真知灼见。 这项工作得到了以色列科学基金会、两国科学基金会和Minerva中心的部分资助。
工具书类
Cathcart R,Schwiers E,Ames BN(1983)使用荧光二氯荧光素分析法检测过氧化氢的皮摩尔水平。 《Ana Biochem》134:111–116 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Chandler JM、Cohen GM、MacFarlane M(1998)Fas诱导小鼠肝细胞凋亡后caspase-3和caspase-7的不同亚细胞分布。 生物化学杂志273:10815–10818 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Codogno P,Meijer AJ(2005)自噬和信号传导:它们在细胞存活和细胞死亡中的作用。 细胞死亡差异12(增刊2):1509–1518 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Cuervo AM(2003)自噬和寻觅——当“你能吃的东西”都是你自己时。 科学老龄化知识环境2003:pe25。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Cuervo AM(2004)《自噬:疾病与健康》。 趋势细胞生物学14:70–77 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Fass E、Shvets E、Degani I、Hirschberg K、Elazar Z(2006)微管支持饥饿诱导的自噬体的产生,但不支持其靶向性和与溶酶体的融合。 生物化学杂志281:36303–36316 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Giles NM、Watts AB、Giles GI、Fry FH、Littlechild JA、Jacob C(2003)半胱氨酸蛋白功能的金属和氧化还原调节。 化学生物10:677–693 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Glading A,Uberall F,Keyse SM,Lauffenburger DA,Wells A(2001)表皮生长因子受体信号下游的M-calpain激活需要膜近端ERK信号。 生物化学杂志276:23341–23348 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Hemelaar J、Lelyveld VS、Kessler BM、Ploegh HL(2003)单一蛋白酶Apg4B对自噬相关的泛素样蛋白GATE-16、MAP1-LC3、GABARAP和Apg8L具有特异性。 生物化学杂志278:51841–51850 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Huang WP,Klinsky DJ(2002)《酵母自噬:分子机制综述》。 细胞结构功能27:409–420 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Ichimura Y、Kirisako T、Takao T、Satomi Y、Shimonishi Y、Ishihara N、Mizushima N、Tanida I、Kominami E、Ohsumi M、Noda T、Ohsimi Y(2000)泛素样系统介导蛋白质脂质氧化。 自然408:488–492 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Jabs T(1999)活性氧中间体作为植物和动物细胞程序性死亡的介质。 生物化学药理学57:231–245 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Kabeya Y、Mizushima N、Ueno T、Yamamoto A、Kirisako T、Noda T、Kominami E、Ohsumi Y、Yoshimori T(2000)LC3是酵母Apg8p的哺乳动物同源物,加工后定位于自噬体膜。 欧洲工商管理硕士J 19:5720–5728 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Kabeya Y、Mizushima N、Yamamoto A、Oshitani-Okamoto S、Ohsumi Y、Yoshimori T(2004)LC3、GABARAP和GATE16根据II型的形成定位于自噬体膜。 细胞科学杂志117:2805–2812 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Kiffin R,Bandyopadhyay U,Cuervo AM(2006),氧化应激和自噬。 抗氧化剂氧化还原信号8:152–162 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Kirisako T,Baba M,Ishihara N,Miyazawa K,Ohsumi M,Yoshimori T,Noda T,Ohsami Y(1999)自噬体的形成过程用酵母中的Apg8/Aut7p进行追踪。 细胞生物学杂志147:435–446 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Kirisako T、Ichimura Y、Okada H、Kabeya Y、Mizushima N、Yoshimori T、Ohsumi M、Takao T、Noda T、Ohsumi Y(2000)可逆修饰调节自噬所必需的Apg8/Aut7的膜结合状态和细胞质到液泡靶向途径。 细胞生物学杂志151:263–276 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Kissova I、Deffieu M、Samokhvalov V、Velours G、Bessoule JJ、Manon S、Camougran N(2006)《酵母中的脂质氧化和自噬》。 自由基生物医药41:1655–1661 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Klionsky DJ、Cregg JM、Dunn WA Jr、Emr SD、Sakai Y、Sandoval IV、Sibirny A、Subramani S、Thumm M、Veenhuis M、Ohsumi Y(2003)酵母自噬相关基因的统一命名法。 Dev细胞5:539–545 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Kumanomidou T、Mizushima T、Komatsu M、Suzuki A、Tanida I、Sou YS、Ueno T、Kominami E、Tanaka K、Yamane T(2006)人类Atg4b的晶体结构,一种用于自噬体形成修饰物的加工和去结合酶。 分子生物学杂志355:612–618 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Lang T、Schaeffeler E、Bernreuter D、Bredschneider M、Wolf DH、Thumm M(1998)Aut2p和Aut7p这两种新型微管相关蛋白对于将自噬小泡输送到液泡至关重要。 EMBO期刊17:3597–3607 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Lee YJ,Cho HN,Soh JW,Jhon GJ,Cho CK,Chung HY,Bae S,Lee SJ,Lee YS(2003)氧化应激诱导的细胞凋亡是由ERK1/2磷酸化介导的。 实验细胞研究291:251–266 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Macip S,Igarashi M,Berggren P,Yu J,Lee SW,Aaronson SA(2003)诱导活性氧物种对p53介导的细胞命运决定的影响。 摩尔细胞生物学23:8576–8585 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Mann SS,Hammarback JA(1994),轻链3的分子表征。 MAP1A和MAP1B的微管结合亚单位。 生物化学杂志269:11492–11497 [ 公共医学 ] [ 谷歌学者 ]
Marino G,Uria JA,Puente XS,Quesada V,Bordallo J,Lopez-Otin C(2003)人类自噬蛋白,一个可能通过自噬参与细胞降解的半胱氨酸蛋白酶家族。 生物化学杂志278:3671–3678 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Mizushima N,Kuma A,Kobayashi Y,Yamamoto A,Matsube M,Takao T,Natsume T,Ohsumi Y,Yoshimori T(2003)Mouse Apg16L,一种新型WD-重复蛋白,以Apg12–Apg5结合物为自噬隔离膜靶点。 细胞科学杂志116:1679–1688 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Mizushima N、Yamamoto A、Hatano M、Kobayashi Y、Kabeya Y、Suzuki K、Tokuhisa T、Ohsumi Y、Yoshimori T(2001)使用Apg5缺陷小鼠胚胎干细胞解剖自噬体形成。 细胞生物学杂志152:657–668 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Mousavi SA、Kjeken R、Berg TO、Seglen PO、Berg T、Brech A(2001)空泡质子泵抑制剂对肝脏异噬和自噬的影响。 Biochim生物物理学报1510:243–257 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Ohsumi Y(2001)自噬的分子解剖:两个泛素样系统。 Nat Rev Mol细胞生物学2:211–216 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Perriere G,Gouy M(1996)WWW-query:生物序列库的在线检索系统。 生物化学78:364–369 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Petiot A、Ogier-Denis E、Blommaart EF、Meijer AJ、Codogno P(2000)不同类别的磷脂酰肌醇3′-激酶参与控制HT-29细胞宏观自噬的信号通路。 生物化学杂志275:992–998 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Preston TJ、Muller WJ、Singh G(2001)《细胞外H的清除》 2 O(运行) 2 过氧化氢酶通过诱导应激反应抑制HER-2/Neu-transformed rat-1成纤维细胞的增殖。 生物化学杂志276:9558–9564 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Reef S、Zalckvar E、Shifman O、Bialik S、Sabanay H、Oren M、Kimchi A(2006)p19ARF的短线粒体形式诱导自噬和半胱天冬酶非依赖性细胞死亡。 分子细胞22:463–475 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Sagiv Y、Lesses-Miller A、Porat A、Elazar Z(2000)GATE-16是一种膜转运调节剂,与NSF和高尔基v-SNARE GOS-28相互作用。 EMBO期刊19:1494–1504 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Sakurai K,Cederbaum AI(1998)表达细胞色素的HepG2细胞中亚硝酸盐铁诱导的氧化应激和细胞毒性 P(P) 450 2E1。 摩尔药理学54:1024–1035 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Scherz-Shouval R,Sagiv Y,Shorer H,Elazar Z(2003)GATE-16(高尔基内转运调节剂)的COOH末端被人类半胱氨酸蛋白酶HsApg4A切割。 生物化学杂志278:14053–14058 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Shintani T,Klinsky DJ(2004)《健康与疾病中的自噬:一把双刃剑》。 科学306:990-995 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Sitia R,Molteni SN(2004),应激,蛋白质(mis)折叠和信号:氧化还原连接。 科学STKE 2004:pe27。 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Sugawara K、Suzuki NN、Fujioka Y、Mizushima N、Ohsumi Y、Inagaki F(2005)人类Atg4B特异性和催化作用的结构基础,负责哺乳动物自噬。 生物化学杂志280:40058-40065 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Suzuki K、Kirisako T、Kamada Y、Mizushima N、Noda T、Ohsumi Y(2001)APG基因协同功能组织的自噬前结构对自噬体的形成至关重要。 EMBO期刊20:5971–5981 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Tanida I、Sou YS、Ezaki J、Minematsu Ikeguchi N、Ueno T、Kominami E(2004a)HsAtg4B/HsAppg4B/自噬蛋白-1切割三种人Atg8同源物的羧基末端,并使微管相关蛋白轻链3-和GABAA受体相关蛋白磷脂偶联物脱水。 生物化学杂志279:36268-36276 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Tanida I,Ueno T,Kominami E(2004b)哺乳动物自噬中的LC3结合系统。 国际生物化学细胞生物学杂志36:2503–2518 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Tassa A,Roux MP,Attaix D,Bechet DM(2003)III类磷脂酰肌醇3-激酶-Beclin1复合物介导C2C12肌管中氨基酸依赖性的自噬调节。 生物化学杂志376:577–586 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
Vanden Hoek TL、Li C、Shao Z、Schumaker PT、Becker LB(1997)在再灌注前缺血期间,分离的心肌细胞产生大量氧化剂。 心脏分子细胞杂志29:2571–2583 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Wang H,Bedford FK,Brandon NJ,Moss SJ,Olsen RW(1999)GABA(A)受体相关蛋白连接GABA(A)受体和细胞骨架。 自然397:69–72 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Xu J,Yu S,Sun AY,Sun GY(2003)氧化剂介导的星形胶质细胞AA释放涉及cPLA(2)和iPLA(2)。 自由基生物医药34:1531–1543 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Yu L,Alva A,Su H,Dutt P,Freundt E,Welsh S,Baehrecke EH,Lenardo MJ(2004)通过半胱氨酸蛋白酶-8调节ATG7-beclin 1自噬细胞死亡程序。 科学304:1500-1502 [ 内政部 ] [ 公共医学 ] [ 谷歌学者 ]
Yu L,Wan F,Dutta S,Welsh S,Liu Z,Freundt E,Baehrecke EH,Lenardo M(2006)选择性过氧化氢酶降解引起的自噬性程序性细胞死亡。 美国国家科学院院刊103:4952–4957 [ 内政部 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
关联数据 本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。