摘要
我们研究了声波刺猬(Shh)在面部结构发育中的作用,在神经嵴细胞(NCC)迁移至第一鳃弓(BA1)之前,通过剥夺鸡胚这种形态原的最前端来源,包括前肠的前索板和前腹内胚层。整个前额,包括前肠内胚层,在5-10体节期(ss)被切除,导致在7-ss之前进行手术时没有下颌。如果胚胎在8到10秒时被剥夺前额,那么它们随后被赋予较低的喙。在早期手术的胚胎中,NCC正常迁移至BA1,但数小时后发生大量凋亡。在后期进行额头切除时,没有发生细胞死亡。在这种情况下嘘腹侧前肠内胚层向尾部延伸超过切除极限,我们假设没有嘘早期手术胚胎内胚层的产生可能是NCC凋亡和BA1发育失败的原因。因此,我们为7-ss之前手术的胚胎提供了外源性Shh。在这种情况下,下颌骨的发育得到了挽救。因此,来自腹侧前肠内胚层的Shh可以确保NCC在BA1发育的关键阶段存活。
脊椎动物面部骨骼的形成是由复杂的发育过程所导致的,这需要颅神经嵴细胞(NCC)的迁移和一系列上皮-间充质相互作用。来自后间脑和前中脑的NCCs产生额钠骨,而来自后中脑和菱形1和2的NCCs定植于第一鳃弓(BA1),形成上颌骨和下颌骨的骨骼(1–三). 神经嵴的这个吻侧区域被命名为FSNC(面部成骨神经嵴)(4). 后菱形肌产生神经干细胞,参与舌骨软骨内侧和后部的形成(1–三).
在细胞迁移开始之前,切除5至6体节期(ss)鸡胚中的完整FSNC,会导致显著的表型,其中面部过程和骨骼不发育,前脑和中脑形态受到干扰(4). 如果提前(3秒)摘除头部神经干细胞,最前面的头部囊泡不会发育,两眼无晶状体眼在中线融合,导致睫状肌麻痹(5). 这种表型类似于人类前脑无裂畸形(HPE)的最严重形式,该综合征包括多种畸形,如下颌骨完全缺失、前脑半球分裂失败、睫状肌麻痹、长鼻畸形和面部裂伤。
的突变音猬因子(嘘)在人类HPE病例中发现了编码形态原Shh的基因(6,7). 此外嘘在小鼠中发现,该基因是许多形态发生过程所必需的,包括肢体极性的建立、神经系统的背腹模式、前肠、轴骨和头骨的发育(8–12).
最近的研究有助于理解Shh在面部发育中的作用。通过植入分泌抗体的杂交瘤细胞在7-10秒时中和鸡颅骨间质中的Shh作用,导致治疗24小时后FSNC(面部成骨神经嵴)细胞死亡,并导致BA1衍生物丢失(13). 在小鼠NCC中特异性阻断刺猬信号导致类似的细胞死亡动力学,随后出现严重的头骨异常(14). 这两项研究都表明Shh对迁移至BA1的FSNC细胞的存活至关重要。突变小鼠中Shh的缺失导致BA1在发育过程中基因表达强烈异常。在这些小鼠中索氏9型,扭曲、和Fgf8型不存在(15,16). 同样,斑马鱼嘘或第个,共个平滑的(平滑(Smo))导致颅面软骨缺失(17). 然而,这些研究都没有确定对面部骨骼发育至关重要的Shh的来源。可能的候选者是弦前板(PcP)、前肠内胚层、底板(FP)、脊索(No)和面部外胚层。
最近有研究表明,前脑神经外胚层和面部外胚层产生的Shh对上面部和鼻额芽的发育至关重要,而抑制这些Shh来源对BA1衍生物没有影响(18,19)这表明对BA1发育至关重要的Shh的来源可能是咽内胚层。在我们小组之前的一项研究中,NCC向BA1迁移路径中的前肠内胚层移植物诱导了下喙骨骼的复制(20). 相反,切除早期鸡胚中相应的内胚层区域(5-6-ss)阻止了属于BA1的明确软骨成分的发育(20). 其他实验表明,神经干细胞只有与前肠内胚层和上颌外胚层共培养或共移植才能分化为软骨(21–23). 此外,在斑马鱼胚胎的内脏骨骼发育中发现了缺陷卡萨诺瓦(中国科学院)和邦妮和克莱德(波恩)内胚层不发育的突变体(24).
在本文中,我们分析了前肠内胚层产生的Shh在BA1的模式化及其骨骼衍生物的开发中的作用。在体液期的早期,嘘在No和FP前部的前肠背侧表达。在PcP和前腹内胚层也表达。然而,这种腹侧内胚层表达嘘直到6-ss,仍局限于前脑的横向水平。它仅从7-8秒向前延伸到前脑-中脑边界的尾部。由于时间/空间受到管制嘘在腹侧前肠中表达,我们能够通过检测7-ss前后剥夺前额区的鸡胚来证明前肠内胚层产生的Shh在BA1发育中的作用。我们的结果表明,在前额Shh的不同来源中,前肠内胚层提供BA1发育所需的早期信号。
结果
时空调控嘘5-至25-ss鸡胚头部区域的表达。
要分析嘘在头部发育早期表达,将5~25-ss的鸡胚在整座山上杂交,然后在矢状面和副矢状面连续切片上切割。这些发育阶段包括头部神经干细胞的分层和迁移。
5秒时(图1A类和A类′),嘘转录物存在于No和相关FP中,对应于假定间脑和中脑的水平,与位于前面的PcP连续。这个嘘-阳性PcP毗邻吻腹内胚层,也表达嘘.在6-ss时,嘘腹侧内胚层的转录物几乎没有达到前脑-中脑边界的水平(图1 B类和B类′).
图1。
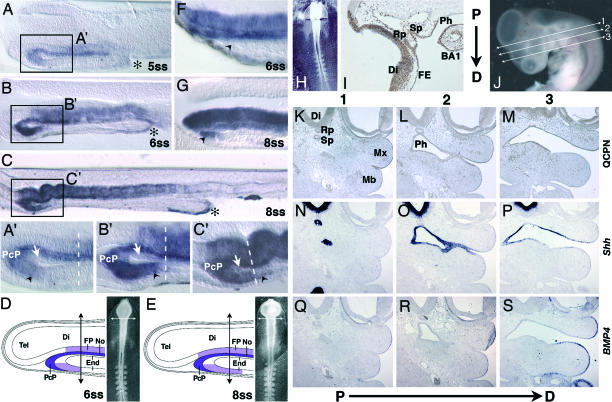
Shh在咽内胚层的表达。(A类和A类′)5-ss鸡胚的矢状切面(50μm)显示嘘在中线细胞、PcP、前腹前肠内胚层(黑箭头)和更后面的AIP(星号)中表达。(B类和B类′)6-ss时,嘘转录物仍然存在于前腹内胚层(黑色箭头)中,但在背前肠内胚层中仍然不存在(白色箭头)。在此阶段,PcP是Shh最前端的来源,与嘴侧腹侧内胚层密切接触。(C和C′)在8-ss时嘘腹侧前肠内胚层出现表达,并越过前脑-中脑边界(虚线)。(D类和E类)的示意图嘘在6和8秒的鸡胚上的表达和前额切除水平(箭头)。末端,内胚层;电话,端脑;Di,间脑;FP,地板;不,脊索。(F类)额头切除后6秒后鸡胚的矢状切面。剩余的腹前肠内胚层(黑色箭头)缺乏嘘抄本。(G公司)在8-ss时,存在表达Shh的前肠内胚层(黑色箭头)。(H(H)–S公司)前额鹌鹑/鸡嵌合体。(H(H))将鹌鹑前额移植到6-ss的鸡胚上后立即进行Chimera。(我)在6-ss处嫁接的E3.5嵌合体的矢状截面(7μm)。QCPN免疫染色(棕色)显示鹌鹑细胞形成腹侧Di和最前面的中脑、面部外胚层(FE)、Rathke囊(Rp)、Sessel囊(Sp)、BA1的背侧和腹侧咽内胚层(Ph)以及BA1的外胚层。(J型)E4.5鹌鹑/鸡嵌合体,具有近端(P→D)连续切片的指示(第1-3行)。(K(K)–S公司)用QCPN处理近端骨骼连续切片(K(K)–M(M))或是就地杂交用于嘘(N个–对)或骨形态发生蛋白4(问–S公司). 在E4.5,鹌鹑细胞(K(K)–M(M))显示E3.5胚胎中观察到的相同分布(我)形成Rathke氏囊上颌芽(Mx)的外胚层、间脑腹侧上皮、Sessel氏囊内胚层和咽前部。(N个–对)嘘mRNA存在于Di、Sp和Ph中(问–S公司)英国标准普尔4mRNA存在于Mx和下颌骨(Mb)的外胚层。
8秒时(图1 C和C′),PcP保持在最前面嘘-表达结构。在这个阶段,它与前腹内胚层形成连续的表达模式,因此很难区分这两种组织之间的边界。从这个阶段开始(8-到10-ss),嘘表达在前腹前肠内部向后进行,而背前肠内胚层仍然被剥夺嘘抄本。嘘早在5-6-ss时就存在于前肠门静脉(AIP)中的表达(图1 A类和B类)当AIP向后移动时,不会扩散到正在生长的腹侧内胚层,因此腹侧前肠内胚层的大部分仍然没有嘘发展期间的成绩单(图1C). 12秒时,未来的口腔外胚层与嘘-产生内胚层,开始表达嘘(未显示数据)。高达16-ss,嘘头部腹侧区域的表达仅限于口腔膜外胚层和前肠内胚层的前部(图4,作为支持信息发表在PNAS网站上)。
第15阶段杂交鸡胚(汉堡和汉密尔顿编号;25-ss)显示嘘在咽部和肠道内胚层均有表达,尤其是在BA1内胚层。嘘在这一阶段也表达于前鼻突出外胚层(图4)。
BA1内胚层的起源。
探讨腹侧内胚层表达的命运嘘在体节早期,并观察是否可能包括内胚层条纹,在我们之前的实验中,内胚层能够诱导额外的下颌发育(20),我们将鸡胚的整个额头区域替换为5-6-ss[胚胎日(E)1.5](图1H(H))与对应的鹌鹑胚胎(n个= 5). 鹌鹑/鸡额嵌合体在手术后2或3天被杀死(E3.5,n个= 3; E4.5、,n个=2),并在用QCPN单抗处理的连续矢状面切片中进行分析,该单抗可识别鹌鹑细胞。实验表明,不仅鹌鹑型的端脑、间脑和中脑最前部以及面部上外胚层(如预期),BA1咽内胚层的主要部分也是由鹌鹑细胞形成的,因此是从移植物中衍生出来的,而BA1外胚层的主要部分为寄主型(图1我).
为了更精确地重建咽外侧内胚层和外胚层的各自起源,我们分析了E4.5处的两个嵌合体。连续冠状序列切片(见图1J型)用QCPN单克隆抗体进行免疫标记(图1 K(K)–M(M))或与嘘(图1 N个–对)或英国标准普尔4(骨形态发生蛋白4) (图1 问–S公司)探针。这些基因分别在BA1的内胚层和远端外胚层中表达。整个嘘-阳性的咽内胚层、塞塞尔氏囊、间脑和Rathke氏囊由鹌鹑细胞组成(图1 K(K)–M(M))(图5,作为支持信息发布在PNAS网站上)。这个英国标准普尔4-下颌骨阳性外胚层为鸡宿主型,与嘘-鹌鹑细胞构成的阳性咽外侧内胚层(图1 M(M),对、和S公司). 其他鹌鹑/鸡前脑嵌合体(n个=2)生长至E10至E11,并检查头部形态。虽然上喙比下喙短,但面部骨骼发育正常(图6,作为PNAS网站上的支持信息发布)。
这些结果表明,在Couly发表的实验中等. (20)2002年,腹侧内胚层的I–III区与Sessel囊、口和第一鳃囊的内胚层相对应。
罗斯特拉尔前肠内胚层对下颌发育至关重要。
我们已经证明,在5-6-ss的胚胎中,嘘嘴侧腹侧内胚层的表达仅限于对应于前脑区的水平。相反,在8秒以后的胚胎中,嘘腹侧内胚层的表达在很大程度上超出了前脑-中脑边界(图1 A类′–C′). 为了研究来自前肠内胚层的Shh在面部骨骼发育中的作用,我们切除了鸡胚的整个前额在ovo中7-ss之前或之后(图1 D类和E类). 当额头在5-6秒时切除时(n个=4),整个吻侧内胚层表达嘘与该形态原的其他来源一起被清除:PcP、No和FP(图1 D类和F类). 手术后48小时(E3.5),胚胎被完全剥夺嘘咽前内胚层和口腔上皮中的转录物(图2G公司和G公司′),与对照胚胎相反(图2 F类和F类′). 当手术在8到10-ss胚胎中进行时(8-ss,n个= 2; 9秒,n个= 1; 10-秒,n个=2),Shh的腹侧内胚层来源维持在切除后的咽前内胚层(图1 E类和G公司),稍后在E3.5,嘘在BA1的前咽内胚层和口腔上皮中发现转录物(图2 H(H)和H(H)')与对照组相同。这些结果表明嘘这些组织中的表达取决于嘘在前腹前肠内胚层。
图2。
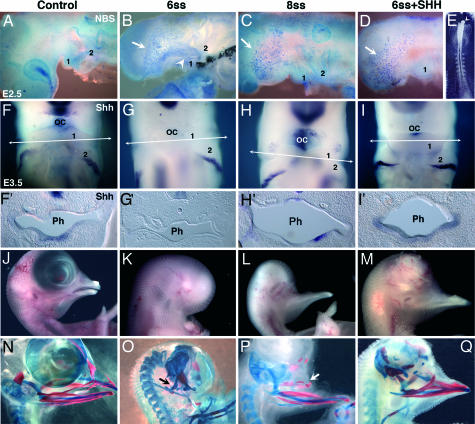
孔内胚层Shh对NCC存活和面部骨骼发育的作用。(A类–D类)硫酸尼罗兰(NBS)对鸡胚的细胞死亡分析全部. (A类)E2.5对照鸡胚胎。(B类–E类)前额在6-ss(白箭头)时切除的胚胎BA1('1')中NBS染色强(B类)但不是在8-ss(C)或在6-ss进行额头切除并用浸有Shh(100μg/ml)的肝素珠(白色箭头)替代后(D类和E类). 然而,在头部前部的剩余区域,细胞大量死亡(B类–D类,白色箭头)。(F类–我)整体安装就地杂交嘘E3.5显示嘘对照胚胎口腔(OC)水平的转录物(F类),胚胎在8秒时切除(H(H))和一个移植了Shh珠的6-ss切除胚胎(我)与非移植胚胎相比(G公司),不显示嘘口腔水平的表达。(F类′–我′)BA1水平的横截面(50μm)(箭头F类–我).嘘mRNA存在于对照胚胎的咽(Ph)内胚层(F类′),胚胎在8-ss时切除(H(H)′)和Shh移植胚胎(我′). 相反,胚胎在6-ss时被切除(G公司')被剥夺嘘BA1咽内胚层中的mRNA。(J型–问)形态学(J型–M(M))和骨骼(N个–问)软骨用阿尔西安蓝染色,骨用茜素红染色,对E11至E12胚胎进行分析。(K(K)和O(运行))6秒时切除的胚胎中没有上下喙。仅保留了Meckel软骨的近端部分(关节方软骨)(O(运行),黑色箭头)。(我和对)8秒时额头切除后,正常的下喙与对照组相似(J型和N个)发育,也有上颌骨发育不良的骨元素(白色箭头)。(M(M)和问)一个E12胚胎,在6-ss时切除并用Shh珠处理;存在与下颌骨和上颌骨相对应的软骨和骨骼。
当手术改变的胚胎长大到E9–E11时,我们可以看到在5到6秒时手术的胚胎没有上下喙。只有方形软骨和关节软骨发育(n个=第8页,共8页)(图2 K(K)和O(运行)). 在8到10秒时手术的胚胎被剥夺了上喙,但有一个下喙(n个=第5页,共5页)(图2 我和对)具有正常的骨骼结构。上喙没有鼻中隔和前颌骨,通常由前鼻芽发育而来。因此,来自前肠内胚层剩余部分的Shh对BA1的发育至关重要。
前肠内胚层的缺失不会干扰头部NCC向BA1的迁移,但会阻止NCC存活。
为了理解为什么在7-ss之前被剥夺了整个前额区域的鸡胚没有发育出颌骨,但通常定植于BA1的NCC没有受到手术的影响,我们使用HNK1单克隆抗体在E2.5观察了BA1中NCC的迁移。7-ss之前手术的胚胎(n个=4)显示了BA1中的许多NCC(图7,作为PNAS网站上的支持信息发布)。然而,与对照胚胎相比,尼罗兰硫酸盐染色显示手术改变胚胎中BA1(但BA2)出现大量细胞死亡(图2 A类和B类). 相比之下,7-ss后进行手术的胚胎发育成正常的BA1,并且只观察到有限数量的细胞死亡(图2C). 在7-ss之前切除的E3.5胚胎中,BA1的大小小于来自对照胚胎和7-ss之后手术胚胎的BA1(图2 F类–H(H)). 在包括对照组在内的所有胚胎中,BA2、BA3和BA4显示出同等数量的细胞死亡,主要位于鳃弓的近端区域(图8,作为PNAS网站上的支持信息发布)。因此,前肠内胚层的Shh对BA1中NCC的迁移不是必需的,但对NCC的存活至关重要。相反,切除前肠内胚层并取走Shh源并不影响后弓的发育,尤其是BA2的发育,正如7-ss之前手术的胚胎舌骨的正常发育所证明的那样(图2O(运行)).
重组Shh可以替代BA1中的前肠内胚层。
在5到6个月龄的鸡胚胎中,早期切除前额可以阻止BA1的发育。相反,在后期切除(8-10-ss)(前肠内胚层分泌Shh的区域向尾部延伸)后,下颌发育。我们假设来自腹侧前肠内胚层的Shh是这一过程所需的信号。为了验证这一假设,我们用100μg/ml重组小鼠Shh浸泡在7-ss之前切除的鸡胚中的肝素珠替代前脑区域。Shh珠与剩余腹侧内胚层和外胚层的最前部紧密接触(图2E类). 一些胚胎用PBS-BSA浸泡过的肝素珠作为对照。E2.5处理胚胎的尼罗兰硫酸盐染色显示,正如预期的那样,Shh珠可以拯救BA1中的NCC死亡(n个= 5) (图2D类). 有趣的是,嘘在这些胚胎的BA1内胚层中恢复表达(图2 我和我′). 此外,在E10–E12 Shh处理的胚胎中,喙的形成(n个=第7页,共10页),发生BA1衍生软骨和骨骼(图2 M(M)和问). 相反,对照胚胎移植PBS-BSA浸泡珠(n个=3)显示大量细胞死亡,仅发育BA1的近端结构,如方关节突,7-ss之前未处理的胚胎也一样(未显示)。这些结果表明,前额通过以下途径对下颌发育起关键作用嘘来自前肠内胚层的信号。此外,他们认为Shh在前肠的最初来源于PcP产生的Shh的诱导,而PcP产生的Shh可以被这种信号形态发生素的外源来源所取代。最后,无论是PcP还是浸有Shh的珠子都能在腹侧内胚层诱导和繁殖Shh的产生。
Shh是颌骨发育信号级联的上游。
因为Shh珠子能够恢复嘘在7-ss之前切除的胚胎BA1内胚层中的表达,我们进一步研究了在没有Shh的初始腹侧内胚层来源的情况下基因表达的变化。因此,我们分析了BA1发育标记转录因子的表达坑x1、和,共Fgf8型和英国标准普尔4,参与BA1的近晶模式(25). 在E3.5和E4.5对额头发育胚胎和对照胚胎进行观察。的成绩单Fgf8型,英国标准普尔4、和坑x1在7-ss之前切除的胚胎中,BA1外胚层、口腔上皮和间质中表达下调(图3D类–F类)而它们通常在7-ss后切除的胚胎中表达(图3 G公司–我). 在用Shh珠处理的实验胚胎中Fgf8型,英国标准普尔4、和坑1已在BA1中恢复(图3 J型–我).坑x1在6-ss切除胚胎的视后区E4.5的表达没有下调,但在没有眼睛的情况下,与对照组眼睛相关的两个外侧表达病灶融合在一个横向点上(图3F类)表明该表达独立于内胚层衍生的Shh。这些结果表明,特异性来源于前肠内胚层的Shh对NCC存活具有关键作用,从而允许BA1的模式化和喙的形成。
图3。
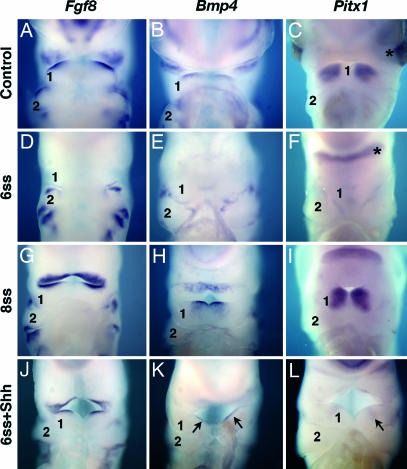
整体安装就地在E3.5至E4.5鸡对照和手术改变的胚胎中进行杂交。(A类–我)Fgf8型,英国标准普尔4、和坑x1对照组BA1的口腔上皮和外胚层中存在mRNA(A类–C)以及8-ss后前额切除的胚胎(G公司–我)但在6-ss时切除额头的胚胎中没有(D类–F类). (J型–我)前额在6-ss时被切除并被Shh珠替代的胚胎显示出这些基因的表达模式(黑色箭头)和发育良好的BA1('1'),而非处理胚胎则显示出BA1尺寸减小和这些基因的错误表达(D类–F类). 星号表示Pitx1在亚轨道区的表达F类在没有眼睛的情况下,两个表达焦点连接在一个横向点上。
讨论
我们在本报告中表明,来自前腹前肠内胚层的Shh是颌骨发育的早期和必要信号。在长达7秒的鸡胚中,对整个前额区域的手术切除不会影响头部NCC向BA1的迁移,但会阻止它们的存活以及上颌骨和下颌骨的形成。前额切除后,用小鼠重组Shh浸泡在咽部内胚层附近的肝素珠挽救了BA1中的细胞死亡,并使下喙和上颌骨及其骨骼成分得以发育。这种治疗能够诱导嘘BA1内胚层和Fgf8型,英国标准普尔4、和坑x1在外胚层。在切除的胚胎中,这些基因的表达下调。
Shh对头部发育的作用最早见于嘘出现前脑无裂、独眼畸形和头骨缺失的突变小鼠(11). 面部畸形也在患有嘘突变(6,7). 尽管这些观察结果表明,如果没有嘘,不可能确定发育中头部的不同Shh来源(PcP、No、FP或肠道内胚层)中的哪一个对面部发育起作用,以及这种作用是通过什么机制和在什么时候发生的。
在我们的工作中,我们首先关注的是嘘前肠腹侧内胚层表达。在5秒时,嘘在与PcP密切接触的假定前脑水平对应的前肠前腹内胚层中检测到转录物。在6-ss时,嘘转录物存在于腹侧前内胚层中,更确切地说,存在于AIP中,但不存在于这两个位点之间。奇怪的是,在这些阶段嘘在腹侧内胚层中,表达域在尾侧方向适度进展,并局限于前肠的前部。它从8秒开始延伸至中脑水平。因此,在颅骨NCC分层和迁移期间(6-至13-ss)(三),腹前肠内胚层由两个区域组成:一个前部区域表达嘘以及更后面的区域嘘(高达16-ss),但AIP除外,AIP始终嘘积极的。这些观察结果可能与之前的研究有关,这些研究表明,前肠内胚层的不同前后区域具有不同的能力来支持骨骼衍生物的发育(20–22). 此外,用鹌鹑替代6-ss鸡胚中的整个前脑区(加上中脑的最前部,包括外胚层、中胚层和内胚层)表明BA1内胚层表达嘘E3.5源自鹌鹑前肠的标记区域。这一结果表明,前肠道内胚层表达嘘产生第一鳃囊的内胚层,包括I–II区和至少部分III区,如参考文献所述。20.
额头切除实验的结果因实施阶段的不同而显著不同。最高7秒,当嘘严格限制在前脑区域,没有下颌发育。相反,当手术稍后进行时(8-10-ss),嘘表达已经在尾部进行,下颌发育。当嘘与PcP相关的表达不再与前肠内胚层连续,后者不表达该基因。然而,Shh的这种天然来源可以被外源形态原所取代。与切片的前肠接触的Shh浸泡的肝素珠能够通过腹侧前肠细胞启动Shh的产生。这种治疗足以确保侵袭BA1的NCC的生存和颌骨骨骼的发育。这一结果强烈表明Shh诱导嘘被转录的基因,一项已经在其他各种系统中记录的观察结果(参见参考文献。9和26以及其中的参考)。此外,Shh对神经干细胞的存活作用也在头部发育期间发生(13–15).
在卡萨诺瓦斑马鱼的突变体,被剥夺了内胚层,内脏-颅骨骨骼完全缺失,而神经颅只受到轻微影响(24). 相反,通过中和神经外胚层和面部外胚层中的抗体抑制Shh导致鼻中隔发育不全,但下喙发育正常(18). 综上所述,这些数据和我们的结果表明,上面部的发育受来自神经外胚层和面部外胚层的Shh控制,而下面部的发育则受来自腹侧前肠内胚层的Shh控制。
我们使用Shh珠作为Shh来源的方法提出了该蛋白对BA1形态发生程序的长期作用的问题。Shh珠能够拯救BA1中的颅骨NCC免于死亡,并允许形成下喙。后来,Shh的存在对于确保控制BA1外胚层近端区域化及其规范的基因的正常表达模式至关重要。英国标准普尔4,Fgf8型、和坑x1在6-ss切除胚胎的BA1外胚层和间充质中没有转录物,但在后期手术的胚胎中存在转录物。此外,Shh珠挽救了早期手术胚胎中这些基因的表达模式。这些数据表明嘘嫁接Shh珠后在BA1内胚层表达,对BA1外胚层的图案形成起作用。因此,来自BA1内胚层的Shh具有不同的作用,这取决于所考虑的组织:它作为NCC的生存信号,如鸡和小鼠所示(13,14)也适用于内胚层本身(15)而它是负责诱导BA1外胚层特异基因表达的上游信号。在这些基因中,Fgf8型先前证明对BA1 NCC的增殖和分化至关重要(4). 我们的结果与观察结果一致嘘突变小鼠,表达Fgf8型,英国标准普尔4,Pax1型,扭曲、和Sox9指数在BA1中丢失(15,16). 此外,最近的研究表明,刺猬信号在斑马鱼口的模式形成中起着至关重要的作用,因此对颌骨的发育也起着重要的作用(27). 另一方面,腹侧前肠内胚层之间的接触表达嘘口腔外胚层早在6-ss发生,随后表达嘘从12秒开始在外胚层(28). 稍后,这种相互作用将负责口腔上皮的近端区域化(29).
总之,目前的研究表明,特定来源于腹前肠内胚层的Shh是颌骨发育和形态发生的关键和早期信号。的表达式嘘前肠内胚层由一个更前面的来源诱导,可能是PcP。一旦嘘在腹侧前肠内胚层(8-ss)建立表达,即使整个前额区域被切除,下喙也可以发育,这表明在这个阶段,上下面部形态发生是两个独立的过程。Shh的腹侧来源负责BA1发育的至少两个步骤:迁移至BA1的颅骨NCC的存活以及BA1外胚层和口腔上皮的模式化。
材料和方法
显微外科。
仅对鸡胚进行额头手术切除在ovo中5至6秒和8至10秒。在这些阶段,未来前脑和中脑之间的界限尚不明确,切除的区域包括未来中脑的一部分。如果与参考文献中描述的实验进行比较。20,本研究中切除的前肠内胚层对应于I区、II区和III区的一部分。前肠移植是从鹌鹑胚胎到鸡胚进行的在ovo中根据参考文献中描述的技术,在5-7-ss下。30再次孵育手术改变的胚胎,并在E2.5–E4.5下处死,以进行全山或切片免疫化学或就地杂交分析和E9–E12用于形态学和骨骼研究。
Shh切除胚胎的处理。
肝素丙烯酸珠(直径≈120μm;Sigma,St.Louis,MO)植入与5-6-ss切除胚胎的腹内胚层和外胚层接触。肝素珠用100μg/ml小鼠重组Shh(明尼苏达州明尼阿波利斯研发系统公司)浸泡,并在37°C下培养1小时,然后在4°C下过夜,然后使用。用PBS处理过的珠子作为对照。
胚胎加工。
现场全山胚胎或切片的杂交分析Fgf8型(31),嘘(9),英国标准普尔4(32)、和坑x1(33)按照参考文献所述进行。26用QCPN(用于鹌鹑细胞)或HNK1单克隆抗体(用于NCC)对整装制剂和石蜡切片进行免疫细胞化学处理,如参考文献所述。4如参考文献所述,通过尼罗兰硫酸盐染色进行全贴装细胞死亡检测。34必要时,切片上用QCPN单克隆抗体进行免疫细胞化学的软骨结构用阿尔西安蓝进行复染。根据标准染色方案,使用阿尔西安蓝对软骨进行染色,茜素红对骨骼进行染色。
鸣谢
我们感谢索菲·古奈特和米歇尔·弗洛马盖特提供的插图。这项工作得到了医学研究基金会和尼维尔高等教育协会(J.M.B.)以及国家科学研究中心和癌症控制协会的研究金支持。
缩写
- NCC公司
神经嵴细胞
- 嘘
音猬因子
- PcP公司
赤道前板块
- BA1公司
第一鳃弓
- E类n个
胚胎期n个
- 不锈钢
体节期
- AIP公司
前肠门。
工具书类
-
1Couly G.、Grapin-Botton A.、Coltey P.、Le Douarin N.M.Development(英国剑桥),1996年;122:3393–3407. doi:10.1242/dev.122.11.3393。[内政部] [公共医学] [谷歌学者]
-
2Kontges G.,Lumsden A.Development(英国剑桥),1996年;122:3229–3242. doi:10.1242/dev.122.10.3229。[内政部] [公共医学] [谷歌学者]
-
三。Le Douarin N.M.,Kalcheim C.神经嵴。英国剑桥:剑桥大学出版社;1999[谷歌学者]
-
4.Creuzet S.、Schuler B.、Couly G.、Le Douarin N.M.程序。国家。阿卡德。科学。美国2004年;101:4843–4847. doi:10.1073/pnas.0400869101。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
5Etchevers H.C.、Couly G.、Vincent C.、Le Douarin N.M.Development(英国剑桥)1999;126:3533–3543. doi:10.1242/dev.126.16.3533。[内政部] [公共医学] [谷歌学者]
-
6Belloni E.、Muenke M.、Roessler E.、Traverso G.、Siegel-Bartelt J.、Frumkin A.、Mitchell H.F.、Donis-Keller H.、Helms C.、Hing A.V.等人,《Nat.Genet》。1996;14:353–356. doi:10.1038/ng1196-353。[内政部] [公共医学] [谷歌学者]
-
7.Roessler E.、Belloni E.、Gaudenz K.、Jay P.、Berta P.、Scherer S.W.、Tsui L.C.、Muenke M.Nat.Genet。1996;14:357–360. doi:10.1038/ng1196-357。[内政部] [公共医学] [谷歌学者]
-
8Echelard Y.、Epstein D.J.、St-Jacques B.、Shen L.、Mohler J.、McMahon J.A.、McMahon A.P.Cell。1993;75:1417–1430. doi:10.1016/0092-8674(93)90627-3。[内政部] [公共医学] [谷歌学者]
-
9Riddle R.D.、Johnson R.L.、Laufer E.、Tabin C.Cell。1993;75:1401–1416. doi:10.1016/0092-8674(93)90626-2。[内政部] [公共医学] [谷歌学者]
-
10Roelink H.、Augsburger A.、Heemskerk J.、Korzh V.、Norlin S.、Ruiz i Altaba A.、Tanabe Y.、Placzek M.、Edlund T.、Jessell T.M.等人cell。1994;76:761–775. doi:10.1016/0092-8674(94)90514-2。[内政部] [公共医学] [谷歌学者]
-
11Chiang C.、Litingtung Y.、Lee E.、Young K.E.、Corden J.L.、Westphal H.、Beachy P.A.自然。1996;383:407–413. doi:10.1038/383407a0。[内政部] [公共医学] [谷歌学者]
-
12Litingtung Y.、Lei L.、Westphal H.、Chiang C.Nat.Genet。1998;20:58–61. doi:10.1038/1717。[内政部] [公共医学] [谷歌学者]
-
13Ahlgren S.C.,Bronner-Fraser M.货币。《生物学》,1999年;9:1304–1314. doi:10.1016/s0960-9822(00)80052-4。[内政部] [公共医学] [谷歌学者]
-
14Jeong J.、Mao J.、Tenzen T.、Kottmann A.H.、McMahon A.P.基因开发2004;18:937–951. doi:10.1101/gad.1190304。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
15Moore-Scott B.A.,Manley N.R.Dev.Biol公司。2005;278:323–335. doi:10.1016/j.ydbio.2004.10.027。[内政部] [公共医学] [谷歌学者]
-
16Washington Smoak I.、Byrd N.A.、Abu-Issa R.、Goddeeris M.M.、Anderson R.、Morris J.、Yamamura K.、Klingensmith J.和Meyers E.N.Dev.Biol。2005;283:357–372. doi:10.1016/j.ydbio.2005.04.029。[内政部] [公共医学] [谷歌学者]
-
17Chen W.、Burgess S.、Hopkins N.Development(英国剑桥),2001年;128:2385–2396. doi:10.1242/dev.128.12.2385。[内政部] [公共医学] [谷歌学者]
-
18Marcusio R.S.、Cordero D.R.、Hu D.、Helms J.A.Dev.Biol。2005;284:48–61. doi:10.1016/j.ydbio.2005.04.030。[内政部] [公共医学] [谷歌学者]
-
19Cordero D.、Marcucio R.、Hu D.、Gaffield W.、Tapadia M.、Helms J.A.J.Clin。投资。2004;114:485–494. doi:10.1172/JCI19596。[内政部] [PMC免费文章] [公共医学] [谷歌学者]
-
20Couly G.、Creuzet S.、Bennaceur S.、Vincent C.、Le Douarin N.M.Development(英国剑桥),2002年;129:1061–1073. doi:10.1242/dev.129.4.1061。[内政部] [公共医学] [谷歌学者]
-
21格雷夫森A.C.,阿姆斯特朗J.B.差异化。1987;35:16–20. doi:10.1111/j.1432-0436.1987。tb00146.x。[内政部] [公共医学] [谷歌学者]
-
22Seufert D.W.,Hall B.K.细胞差异。1990年开发;32:153–165. doi:10.1016/0922-3371(90)90109-a。[内政部] [公共医学] [谷歌学者]
-
23Bee J.,Thorogood P.开发生物学。1980;78:47–62. doi:10.1016/0012-1606(80)90317-6。[内政部] [公共医学] [谷歌学者]
-
24David N.B.、Saint-Etienne L.、Tsang M.、Schilling T.F.、Rosa F.M.Development(英国剑桥),2002年;129:4457–4468. doi:10.1242/dev.129.19.4457。[内政部] [公共医学] [谷歌学者]
-
25Wall N.A.,Hogan B.L.机械。1995年开发;53:383–392. doi:10.1016/0925-4773(95)00453-x。[内政部] [公共医学] [谷歌学者]
-
26Charrier J.B.、Lapointe F.、Le Douarin N.M.、Teillet M.A.Development(英国剑桥),2002年;129:4785–4796. doi:10.1242/dev.129.20.4785。[内政部] [公共医学] [谷歌学者]
-
27Eberhart J.K.、Swartz M.E.、Crump J.G.、Kimmel C.B.Development(英国剑桥),2006年;133:1069–1077. doi:10.1242/dev.02281。[内政部] [公共医学] [谷歌学者]
-
28Withington S.、Beddington R.、Cooke J.Development(英国剑桥),2001年;128:309–320. doi:10.1242/dev.128.3.309。[内政部] [公共医学] [谷歌学者]
-
29Haworth K.E.、Healy C.、Morgan P.、Sharpe P.T.Development(英国剑桥),2004年;131:4797–4806. doi:10.1242/dev.01337。[内政部] [公共医学] [谷歌学者]
-
30马蒂诺维奇P.N.,巴甫洛维奇M.R.自然。1958;182:571–572. doi:10.1038/182571b0。[内政部] [公共医学] [谷歌学者]
-
31Crossley P.H.、Minowada G.、MacArthur C.A.、Martin G.R.Cell。1996;84:127–136. doi:10.1016/s0092-8674(00)80999-x。[内政部] [公共医学] [谷歌学者]
-
32Francis P.H.、Richardson M.K.、Brickell P.M.、Tickle C.Development(英国剑桥),1994年;120:209–218. doi:10.1242/dev.120.1.209。[内政部] [公共医学] [谷歌学者]
-
33Henrique D.、Adam J.、Myat A.、Chitnis A.、Lewis J.、Ish Horowicz D.自然。1995;375:787–790. doi:10.1038/375787a0。[内政部] [公共医学] [谷歌学者]
-
34Jeff P.、Jaques K.、Osmond M.Anat。胚胎。1992;185:583–588. doi:10.1007/BF00185617。[内政部] [公共医学] [谷歌学者]
关联数据
本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。

