D类 二氢吡啶敏感L型钙 2+ 通道(LTCC)是在几乎所有可兴奋细胞的表面膜中发现的多聚蛋白。 LTCC被膜去极化激活,其功能受到PKC等信号分子的调节( 1 ). 这些通道在许多需要钙的信号通路中起着不可或缺的作用 2+ 内流激活细胞内钙 2+ -依赖分子。 在神经元中,Ca 2+ LTCC流入调节基因表达( 2 , 三 )神经生长指导( 4 )和长期增强( 5 )而在肌肉中则有助于收缩。
与神经元和横纹肌相反,动脉平滑肌细胞不会触发动作电位。 在动脉中,管腔内压力的增加会导致平滑肌细胞在生理电位范围(-55至-40 mV)内逐渐去极化。 血管收缩剂,如血管紧张素II( 6 )和UTP( 7 )通过引起膜去极化增加LTCC活性( 8 )PKC的激活( 1 ). 使用传统的成像和电生理技术,已经形成了一个普遍接受的模型,其中膜去极化和PKC增加了单个LTCC的开放概率; 单个LTCC的随机激活增加Ca 2+ 内流,最终导致细胞内游离钙浓度增加([Ca 2+ ] 我 )和收缩( 9 - 11 ). 一个类似的模型已经被提出( 12 ). LTCC介导Ca模型的两个基本原理 2+ 条目是,这些通道的激活是随机的,并且平均而言,所有功能LTCC在特定电压下被激活的概率相似。 然而,到目前为止,由于技术限制,这两个假设尚未经过严格测试。
在本研究中,我们结合电压灯电生理学和消逝场全内反射荧光显微镜观察近膜[Ca的变化 2+ ] 我 (即Ca 2+ 流入)。 由于用这种方法获得的高时间和空间分辨率,我们能够研究LTCC介导的钙的机制 2+ 比以前更详细的条目。 与预期相反,我们观察到相邻LTCC的集群以高开放概率模式运行,这创造了持续Ca的位置 2+ 大量涌入。 我们将这些通道称为“持久LTCC”。有趣的是,需要PKC活动才能打开持久LTCC活动。 PKC激活剂和Bay-K 8644增加Ca 2+ 通过促进额外的LTCC集群在持续Ca中运行而流入 2+ 流入模式。 鉴于PKC和LTCC在可兴奋细胞中普遍存在,Ca 2+ 通过持久性LTCC簇的流入可能是局部和全局Ca生成的一般机制 2+ 信号。
材料和方法 请参见 支持文本 ,发布为 支持信息 在PNAS网站上,获取本节的详细版本。
动脉肌细胞的分离和电生理学。 如前所述,从新鲜解剖的大鼠脑动脉中制备肌细胞( 13 ). 钙 2+ 通过使用Axopatch 200B放大器的全细胞或细胞连接的补丁灯技术配置来记录电流,并使用 pc灯9.0 (Axon Instruments,加利福尼亚州联合市)。 所有实验均在室温(22-25°C)下进行。
全内反射荧光显微镜(TIRF)。 使用配备PlanApo(×60,数值孔径=1.45)透镜和斯坦福光子XR Mega 10增强型电荷耦合器件相机(斯坦福光子公司,加利福尼亚州帕洛阿尔托)的直通式奥林巴斯(纽约州梅尔维尔)TIRF显微镜采集图像。 按照说明校准荧光信号( 14 ). 钙 2+ 火花是通过使用类似于其他人使用的标准来识别的( 15 ).
统计。 数据以平均值±SEM表示。使用Student的 t吨 检验或Wilcoxon符号秩检验。 A类 P(P) <0.05的值被认为是显著的。 星号( * )符号用于图。 1 , 2 , 三 , 4 以说明各组之间的显著差异。
图1。
持久性钙 2+ 动脉平滑肌内流入。 ( A类 )显示三种活性Ca的细胞图像的表面图 2+ 火花点([Ca 2+ ] o个 =2 mM)([Ca 2+ ] o个 ,外部钙浓度)。 下图显示了由曲面图中的白色方块所标记区域的展开2D视图。 痕迹显示[Ca的时间进程 2+ ] 我 在a-d区域( B类 )钙的振幅直方图 2+ 2 mM[Ca的火花 2+ ] o个 ( C类 )暴露于20 mM[Ca的两个代表性细胞的图像 2+ ] o个 .痕迹显示[Ca的时间进程 2+ ] 我 在区域a和b中。痕迹c和c′显示[Ca的连续记录 2+ ] 我 在区域c中( D类 )钙的振幅直方图 2+ 20 mM下的火花[Ca 2+ ] o个 .实心黑线最适合(χ 2 =6.0)到具有以下多分量高斯函数的数据: ,其中 一 和 b条 为常数,[Ca 2+ ] 我 和 q个 (37.9 nM)是细胞内钙 2+ 和钙的量子单位 2+ 流入,分别( n个 =2995个事件)。 直方图中的虚线表示振幅阈值。 ( E类 )平均值±SEM的条形图 不适用 秒 钙的 2+ 迷你网站。
图2。
LTCC是持久性钙的基础 2+ 条目。 ( A类 )Ca之间的关系 2+ 火花振幅及其相关Ca 2+ 电流。 实线是数据的线性拟合。 ( 插入 )[Ca的同时记录 2+ ] 我 而我 钙 在-70 mV时( B类 )量子钙的同步记录 2+ 火花和幺正Ca 2+ -90和-70 mV时的通道活度( 左侧 ). ( 居中 )样品Ca 2+ 通道记录来自一个连接了细胞的补丁。 红线表示钙的量子水平 2+ 闪光和全电导和亚电导Ca的单一通道水平 2+ 频道。 Ca的电压依赖性 2+ 火花(pA单位)和亚电导和全电导钙 2+ 通道( 赖特 ). 实线是数据的线性拟合。 ( C类 )样品[Ca 2+ ] 我 沉默的痕迹( 左侧 ),低( 居中 )、和高( 赖特 ) 不适用 秒 Bay-K 8644使用前后的位置。 ( D类 )[加利福尼亚州 2+ ] 我 来自Ca的记录 2+ 硝苯地平前后的火花点。 ( E类 )平均值±SEM 不适用 秒 钙的 2+ 使用Bay-K 8644或硝苯地平前后的火花点。 ( F类 )钙的振幅直方图 2+ 在控制条件下和Bay-K 8644处理后的火花。 实线表示与高斯函数数据的最佳拟合 图1 使用 q个 38.5 nM的值(χ 2 =3.8)和39.2 nM(χ 2 =4.3)用于控制( n个 =818个事件)和Bay-K 8644( n个 =1453个事件)数据。
图3。
PKC诱导钙持续存在 2+ 火花活动。 ( A类 )无声单元格的图像( 左侧 ),低( 居中 )、和高( 左侧 ) 不适用 秒 地点。 每张图片下方的痕迹显示了[Ca的时间进程 2+ ] 我 在前面的绿色圆圈中( 上部 )PDBu(200 nM; 下部 )治疗。 ( B类 )平均值±SEM 不适用 秒 钙的 2+ 在对照条件下和存在PDBu或PKC抑制肽(100μM)的情况下闪点。 ( C类 )钙的振幅直方图 2+ 在控制条件下和PDBu存在下的火花。 实线最适合使用高斯函数的数据,如 图1 ,使用 q个 38.7 nM的值(χ 2 =1.6)和39.4 nM(χ 2 =2.9)用于控制( n个 =779个事件)和PDBu( n个 =2247个事件)数据。 ( D类 )含有[Ca的膜部分的样品痕迹 2+ ] 我 高于阈值(静止[Ca 2+ ]+40 nM),在控制室中( 上部 )和用活化PKC透析的细胞( 下部 ). [加利福尼亚州 2+ ] 我 在进入细胞内部后约1、2或3分钟记录。 图片位于 左侧 在上述时间点拍摄。 图像中的绿线勾勒出单元格的轮廓。
图4。
低和高- 不适用 o个 LTCC。 ( A类 )之前低活性LTCC的典型细胞附加记录( 左侧 )以及之后( 赖特 )使用200 nM PDBu(-70 mV;20 mM Ca 2+ ). 虚线表示全电导单通道电流水平。 这些轨迹右侧的图表绘制了平均值±SEM 不适用 o个 PDBu前后的低活性和高活性LTCC。 ( B类 )控制条件下高活性LTCC通道的典型细胞附加LTCC记录( 左侧 )200 nM双吲哚甲酰胺(BIM)后( 赖特 ). 条形图显示平均值±SEM 不适用 o个 BIM前后的高活性LTCC。
结果 动脉肌细胞被保存在含有肌浆网钙的生理盐水中 2+ ATP酶抑制剂thapsigargin(1μM)消除Ca 2+ 从细胞内储存释放。 监测Ca 2+ 细胞内充满“快速”钙 2+ 指示剂Fluo-5F(200μM)和过量的“慢”钙 2+ 缓冲EGTA(10 mM)。 使用这种指示剂/缓冲液组合,Ca 2+ 进入细胞后与Fluo-5F结合,产生荧光信号。 然而,这种荧光是短暂的,因为钙 2+ 迅速转移到较慢、更丰富(且无荧光)的钙 2+ 缓冲液,用作Ca 2+ 水槽。 此过程限制Ca 2+ 钙的指示荧光 2+ 进入事件的时间和空间,从而允许直接观察钙 2+ 流入,流入( 16 ). 以30-90 Hz的频率采集图像。
首先,我们检查了膜下[Ca 2+ ] 我 在用含有生理浓度钙的盐水浸泡的细胞中 2+ (2毫米; 图1 ). 膜电位保持在-70 mV以增加Ca的驱动力 2+ 进入并保持低LTCC开放概率( P(P) o个 ≈ 10 -5 -10 -8 ) ( 17 , 18 ). 图1 A类 表明,尽管大多数质膜是光学沉默的(即缺乏钙 2+ 内流),该代表性细胞有三个高钙区域 2+ 流入(“Ca 2+ 火花”)。 这些Ca的平均面积 2+ 火花为0.81±0.01μm 2 或≈典型动脉肌细胞表面膜的0.08%。
我们调查了Ca 2+ 火花点随机出现在每个细胞的表面膜内。 为此,每个细胞被划分为方形(8×8像素)区域,Ca的数量 2+ 对每个网格单元中检测到的火花进行计数。 每个细胞的所有网格细胞计数生成直方图,χ 2 进行了检验,以检验计数分布遵循泊松分布的无效假设。 尽管Ca的具体位置 2+ 细胞之间的火花不同 2 测试表明存在显著差异( P(P) <0.01)Ca的空间分布 2+ 火花和预测的每个单元中的泊松分布,表明Ca的空间分布 2+ 火花并不是随机的。
Ca的检查 2+ 火花振幅直方图( 图1 B类 )在-70 mV和2 mM外部Ca下 2+ 表明Ca的振幅 2+ 进入事件范围为18至280 nM( n个 = 97). 大多数事件的振幅接近或达到我们的振幅检测阈值18nM,表明存在频繁的阈下事件。 为了避免这个问题,我们增加了Ca的驱动力 2+ 通过提高外部钙流入 2+ 到20 mM。由于我们的内溶液中存在高浓度EGTA(10 mM),提高了外部Ca 2+ 从2到20 mM没有显著增加静息状态下的游离钙水平 2+ 在这些实验中( P(P) > 0.05). 图1 C类 (另请参阅电影1,其发布为 支持信息 PNAS网站上)包含两个代表性细胞的图像,显示离散Ca 2+ 存在20 mM外部Ca时的进入事件 2+ .与浸泡在2 mM Ca中的细胞一样 2+ ,加利福尼亚州 2+ 火花活动高度局限,大多数质膜明显不参与。 钙的振幅直方图 2+ 20 mM Ca的流入事件 2+ 如所示 图1 D类 使用类似于del Castillo和Katz实施的方法( 19 ),我们用Ca量子单位的多分量高斯函数拟合直方图 2+ 海拔37.9nM。 这些数据表明Ca 2+ 进入LTCC本质上是量子的,Ca的大小 2+ 火花取决于激活的量子数。
与单通道数据分析类似,我们测定了钙的活性 2+ 通过计算 不适用 秒 每个迷你网站的,其中 n个 是量子能级的数量,并且 P(P) 秒 是给定Ca的概率 2+ 迷你网站处于活动状态。 我们发现Ca的分布 2+ 闪光点 不适用 秒 为双峰型,平均值为0.07±0.01( n个 =39)和1.25±0.16( n个 = 43) ( 图1 E类 ). 基于此分析,我们将Ca分组 2+ 迷你网站分为三类:Silent Ca 2+ 默认情况下,sparket站点具有 不适用 秒 共0个; 低的 不适用 秒 站点的活动水平最低(0< 不适用 秒 <0.2),仅偶尔开放相对较短的时间; 和高 不适用 秒 (>0.2)站点具有几乎连续的Ca 2+ 持续数秒的涌入。 钙的持续时间 2+ 低水位流入事件 不适用 秒 位点范围为11~396ms,平均持续时间为79±8ms( n个 = 29). 钙 2+ 大量涌入事件 不适用 秒 位点的持续时间范围更广(11-5148ms),平均持续时间更长(372±66ms, n个 = 41; P(P) <0.05)比低 不适用 秒 地点。 值得注意的是,尽管Ca的持续时间 2+ 流入事件是可变的,Ca 2+ 流入部位稳定,在实验期间(≈5-20分钟)显示出明显的活动迹象。
火花和全细胞电流的同步记录(参见 图2 A类 和 B类 )发现Ca 2+ 火花总是与相应的向内钙联系在一起 2+ 电流。 钙的振幅之间的关系 2+ 火花及其对应的Ca 2+ 电流是线性的( 图2 A类 ). 我们使用了这种关系的斜率(86 nM/pA; 第页 2 =0.88)转换Ca 2+ 火花振幅(nM)转换为pA。 图2 B类 左侧 和 赖特 显示了量子Ca振幅的稳态电压依赖性 2+ 计算出的pA单元中的火花放电事件随电压线性下降,斜率电导为10.1±0.2 pS。为了进行比较,检查了单个LTCC的稳态电流-电压关系(参见 图2 B类 居中 ). 这些单一Ca 2+ 使用20mM Ca在膜片钳技术的细胞附着配置中记录通道电流 2+ 作为电荷载体。 其他人报告( 20 ),LTCC在全导和亚导状态下运行,斜率电导值分别为10.9±0.2和6.8±0.2 pS。 这些电流被硝苯地平(10μM)阻断,但对非选择性阳离子通道阻滞剂SKF-96365(1μM)不敏感。 此外,使用20 mM Ba 2+ 作为电荷载体导致电流振幅大约加倍(数据未显示)。 综上所述,这些数据支持这些电流的LTCC起源。 注意,全电导LTCC电流的电流-电压关系与量子Ca的电流-电压关系相似 2+ 火花(以计算的pA单位)和相关的Ca 2+ 电流,表明量子Ca 2+ 火花是通过打开单个全导电LTCC或同时打开两个亚导电LTCC产生的( 图2 B类 左侧 ).
为进一步支持Ca 2+ 火花由LTCC产生,我们检测了二氢吡啶拮抗剂和激动剂对钙的影响 2+ 火花。 我们发现LTCC阻断剂硝苯地平(10μM)可以消除钙 2+ 火花( 图2 D类 )和Bay-K 8644(500 nM)增强了它们( 图2 C类 和电影2,发布为 支持信息 在PNAS网站上)。 Bay-K 8644活化静态钙 2+ 沉默部位的通道和增加的钙 2+ 低火花活动 不适用 秒 站点( P(P) < 0.05). Bay-K 8644没有增加高火花活性 不适用 秒 站点( P(P) >0.05),表明这些部位的通道活性最大。 重要的是,Bay-K 8644增加了钙的数量 2+ 在不改变量子事件值(39.2 nM)的情况下,所有振幅的火花。 此时可以得出两个重要结论。 首先,在心里( 21 ),药理和生物物理数据支持钙 2+ 动脉肌细胞中的火花是由二氢吡啶敏感LTCC的开放产生的。 其次,Bay-K 8644的实验表明,LTCC簇可能通过药理学手段被强迫进入持续门控模式。
以前的报告表明,PKC增加了LTCC的活动( 1 ). 我们测试了PKC激活增加稳态Ca的假设 2+ 通过刺激钙流入 2+ 火花活动。 使用PKC激动剂佛波醇12,13-二丁酸酯(PDBu;200 nM),我们发现PKC刺激增加Ca 2+ 通过激活先前沉默的钙离子注入 2+ 闪光点( 图3 A类 左侧 )并通过增加低 不适用 秒 站点( 图3 A类 居中 ; 电影3,发布为 支持信息 在PNAS网站上)。 与Bay-K 8644类似,PDBu在 不适用 秒 站点( 图3 A类 赖特 )再次表明这些通道的活性最高,对量子事件的振幅没有影响(39.4 nM; 图3 C类 ). 最后,Ca 2+ 在用高特异性PKC抑制肽(100μM)透析的细胞中从未观察到闪光活性,即使在应用PDBu后也是如此( 图3 B类 和图5,发布为 支持信息 在PNAS网站上; P(P) < 0.05, n个 = 7). 综上所述,这些数据表明PKC对持久性Ca是必要的 2+ 火花活动。 然而,值得注意的是,即使在PKC活性很高的条件下(即存在PDBu),Ca 2+ 流入仍然局限于质膜中数量有限的小位点。
PKC募集持续钙的观察 2+ 只有一小部分表面膜的内流位置令人惊讶,因为LTCC可能广泛分布在平滑肌细胞的质膜中( 18 , 22 ). PKC激活期间,这种受空间限制的持续LTCC活性的可能机制是,并非所有的Ca 2+ 通道在功能上耦合到PKC。 为了解决这个问题,我们使用免疫荧光和共焦成像检查动脉肌细胞中LTCC(CaV1.2α亚单位)和PKC(α和β亚型)的空间分布(图6 A类 ,发布为 支持信息 在PNAS网站上)。 先前的研究表明,α和β是脑血管平滑肌表达PKC的主要亚型( 23 ). 正如预期的那样,CaV1.2蛋白沿着动脉肌细胞的表面膜扩散分布。 然而,我们注意到,存在CaV1.2相关荧光略微升高的区域。 这些病灶的来源尚不清楚,但可能反映出膜折叠不均匀或CaV1.2通道分布的微小变化。
与CaV1.2不同,PKC相关荧光高度限制在直径为0.82±0.04μm的簇( n个 =86)(图6 B类 ). 这些PKC簇中的绝大多数(>75%)位于或接近(≤1μm)质膜。 请注意,从细胞表面发现的大于1μm的PKC簇可能位于表面肌膜,因为动脉肌细胞具有细胞膜内折(小窝)。 这些数据表明,PKC的空间局限性分布是PKC激活期间局部持续LTCC活性的基础。
这些发现表明,持续钙的位置和数量 2+ 流入点受PKC而非LTCC的空间分布限制。 为了验证这个假设,我们监测了钙 2+ 用含有PKC催化亚单位(0.1单位/ml)的细胞内溶液透析的细胞内流入。 我们发现活化PKC的内灌注可诱导持续钙 2+ 通过细胞膜的大部分流入(参见 支持文本 有关此分析的详细信息)。 在所示的代表性单元格中 图3 D类 实验开始后约3分钟,绝大多数(>95%)的成像质膜持续钙 2+ 流入。 在七个独立的实验中也得到了类似的结果。 与此形成鲜明对比的是,成像膜中持续钙离子的最大比例 2+ 对照细胞的内流始终很小(10±4%, n个 = 10). 综上所述,这些数据进一步支持了以下观点:功能性LTCC广泛表达于表面膜,持久性Ca的位置和数量 2+ 内流位点由PKC在动脉肌细胞中的差异分布决定。
研究钙增加的机制 2+ 为了响应PKC,我们在细胞贴附的补丁中记录了单个LTCC( 图4 ). 使用含有20mM Ca的移液管溶液在-70 mV下进行实验 2+ 作为电荷载体。 与我们的Ca一致 2+ sparket数据显示,膜贴片含有两种活性水平的LTCC(即。, 不适用 o个 ). 平均值 不适用 o个 低活性和高活性贴片分别为0.0003±0.0002和0.30±0.11( n个 =7)。 我们发现PDBu(200 nM;参见 图4 A类 )增加了 不适用 o个 低活性钙 2+ 通道设置为0.4±0.12( n个 =9),与高活性斑块中观察到的水平相似( P(P) < 0.05). 接下来,我们确定PKC活性是否与钙有关 2+ 通道活性高 不适用 o个 补丁。 我们测试了膜透性PKC抑制剂双吲哚甲酰胺(BIM;500 nM)对 不适用 o个 频道。 图4 B类 表明BIM降低了Ca 2+ 通道活性≈94%( P(P) < 0.05, n个 = 8). 这些数据表明,LTCC在低活性或高活性模式下发挥作用,这取决于局部PKC活性的程度。
讨论 本研究中的数据揭示了之前未描述的钙的机制 2+ 进入对二氢吡啶敏感的LTCC,其中看似耦合的小簇通道以高开放概率持续运行,从而产生持续钙的位点 2+ 大量涌入。 这种“持久”的LTCC活动需要激活PKC。 此外,我们的数据表明Bay-K 8644和PKC激活剂增加了Ca 2+ 通过促进额外的LTCC集群在持续Ca中运行而流入 2+ 流入模式。
持续钙的生理意义 2+ 通过持久性LTCC集群的流入意义深远。 持久性钙中的LTCC 2+ 流入模式增加局部[Ca 2+ ] 我 达到与Ca相似的程度 2+ 火花( 图1 ) ( 24 , 25 ). 这表明LTCCs在持续性Ca 2+ 流入模式可能导致[Ca持续升高 2+ ] 我 足够激活附近的Ca 2+ -钙调素、表面膜钙等敏感蛋白 2+ -激活离子通道和ryanodine受体。 确实,持久性钙中的LTCC 2+ 内流模式可能允许肌浆网钙的空间限制性调节 2+ 通过提供局部钙源来加载和释放 2+ 这可能解释了钙复发部位的观察结果 2+ 心室肌细胞的火花活动( 26 )和平滑肌细胞( 27 ). 因此,很有意思的推测是,持久性LTCC调节Ca 2+ 从细胞内储存释放。
虽然多重Ca的振幅和持续时间 2+ 火花和钙 2+ 火花相似,这些Ca之间有重要区别 2+ 信号。 首先,Ca 2+ 火花的持续时间可以比Ca长 2+ 火花( 24 , 25 ). 其次,与钙不同 2+ 火花,钙的振幅 2+ 火花是电压依赖型和多模式的。 第三,Ca 2+ 火花和Ca 2+ 火花具有独特的药理学特征。 例如,硝苯地平可用于阻断钙 2+ 不影响Ca的火花 2+ 火花。 或者,可以使用丁卡因、thapsigargin或ryanodine选择性地阻断Ca 2+ 火花。 未来的研究可以利用这些差异作为潜在的策略来区分这两种根本不同的钙 2+ 信号。
本研究的一个重要观察结果是,高活性钙中持久性LTCC的活性 2+ 火花点比细胞的其他区域大。 为了量化这种差异,我们进行了以下分析。 如果持续LTCC的打开时间≈10 ms( 28 ),的 不适用 秒 在高图像采集率(90 Hz)下获得的高活性位点值应近似于 不适用 o个 这些站点中的LTCC。 这意味着 P(P) o个 如果 不适用 秒 以及频道数量( n个 )在sparket站点中是已知的。 典型的心肌细胞(≈1000μm 2 )每μm有5-10个LTCC 2 ( 18 )因此,我们计算出在一个典型的Ca中有4到8个LTCC(LTCC的密度×火花区) 2+ 迷你网站。 这个值与在大多数闪光点观察到的量子水平的数量一致(大约七个)。 因此,使用 n个 七个通道的值,我们得到 P(P) o个 Ca的 2+ 典型高压通道 不适用 秒 火花位置可能≈0.18( 不适用 秒 / n个 ; 1.25/7)。因为 P(P) o个 该电位下典型低活性LTCC的 -8 ( 17 ),我们的数据表明 P(P) o个 在一个高活跃度的sparket站点中,LTCC的数量可能高达1.8×10 7 倍增(1.8×10 -1 /10 -8 )低于-70 mV的平均信道。我们的单信道数据支持这一结论。 假设有四个功能渠道( n个 )在中所示的修补程序中 图4 B类 ,的 P(P) o个 这个高活性补丁中的通道( 不适用 o个 =0.3)为0.08(0.3/4),接近10 7 高于 P(P) o个 (10 -8 )-70 mV时平均LTCC的( 17 ).
然而,请注意,在超极化膜电位下,全细胞LTCC活性较低( 18 ). 因此,持续性LTCC的数量在-70 mV时必须非常少。我们的全内反射荧光图像显示Ca 2+ sparket活性仅限于质膜的小区域,支持这一结论。 我们通过划分 不适用 o个 在-70 mV(1.21±0.23; n个 =7)由 不适用 o个 (0.3)或 不适用 秒 (1.25)个具有相同电位的高活性LTCC。 这表明,一个(1.21/1.25)到四个(1.21/0.3)持久性LTCC位点可能是稳态Ca 2+ 假设每个火花位点有7个通道,这些数据表明,7至28个(≈5000-10000)LTCC的持续开口决定了全局[Ca 2+ ] 我 在这些细胞中。
生理条件下的动脉膜电位(血管内压在60至120毫米汞柱之间)范围为-55至-40毫伏( 29 , 30 ). 在此电压范围内 P(P) o个 LTCC的变化从≈10 -5 到10 -3 ( 17 , 18 ). 如上所述 P(P) o个 Ca的 2+ 高活性Ca通道 2+ 火花点在-70 mV时约为0.18。因此,即使在从-70 mV到-55到-40 mV的电位范围内去极化期间这些持续性通道的活性没有增加 P(P) o个 持久性钙 2+ 此处报告的通道≈180到10 4 -比平均Ca高倍 2+ 这些电位的通道。 该分析表明Ca 2+ 通过持续性LTCC的内流可能在调节[Ca 2+ ] 我 在膜电位的生理范围内。
我们的数据表明,血管平滑肌中LTCC簇的持续空间受限激活可能是一种机制。 我们发现尽管LTCC广泛分布于这些细胞的表面膜( 18 , 22 ),PKC靶向质膜处或附近的小簇。 PKC激动剂可能会增加持续钙的数量 2+ 通过激活这些PKC簇并诱导形成新的PKC簇而进入位点。 我们的数据表明,未与PKC功能耦合的LTCC可以转换为持久性Ca 2+ 通过与PKC的强制相互作用形成流入模式。 因此,我们的研究结果支持这样的观点,即区域PKC激活增加了 P(P) o个 少量LTCC和多量子Ca 2+ 高程可能来自相邻LTCC随机重叠的开口 P(P) o个 .
目前,尚不清楚为什么我们没有检测到大量钙 2+ 具有可归因于处于亚导电状态的开口LTCC的振幅的火花。 一种可能性是,正如奎尔所建议的那样 等。 ( 17 ),次电导Ca的开路概率 2+ 通道开口一般较低,但持久性钙含量特别低 2+ 迷你网站。 未来的研究应该检验PKC调节LTCC电导的有趣可能性。
我们的发现迫使对描述LTCC介导稳态[Ca的当前随机模型进行修改 2+ ] 我 在动脉肌细胞中( 10 , 11 ). 我们建议稳态Ca 2+ 典型细胞内的内流是由小簇的连续开口产生的( 1 - 4 )除了个别LTCC的不频繁和随机的电压依赖性开口之外,看似耦合的持久LTCC的开口。 LTCC激动剂如Bay-K 8644和PKC激活剂可增加Ca 2+ 通过促进额外的LTCC集群在持续Ca中运行而流入 2+ 流入模式和增加孤立LTCC的开放概率。 由于PKC和LTCC在各种组织中的普遍性和重要性,Ca 2+ 通过持续性LTCC的流入可能是一种广泛的机制,产生持续的钙 2+ 许多细胞类型的信号。 事实上,持续性LTCC、相关PKC分子和由此产生的高亚质膜钙的微结构域 2+ 可能构成局部自给Ca 2+ 能够调节多个细胞过程的信令模块。
致谢 感谢W.J.Lederer博士、Keith W.Dilly博士、Charles F.Rossow博士、Carmen A.Ufret-Vincenty博士、Fred Rieke博士和Bertil Hille博士阅读本手稿。 这项工作得到了国立卫生研究院HL077115、HL07828和HL07312的支持。
作者贡献:L.F.S.、G.C.A.和M.F.N.设计的研究; M.F.N.、G.C.A.和L.F.S.进行了研究; M.F.N.、G.C.A.、V.S.V.和L.F.S.分析数据; L.F.S.、V.S.V.、M.F.N.和G.C.A.撰写了这篇论文。
这篇论文是直接提交给PNAS办公室的(第二轨道)。
缩写:[Ca 2+ ] 我 细胞内游离钙浓度; LTCC,L型Ca 2+ 渠道; PDBu,佛波醇12,13-二丁酯。
工具书类
1 Fish,R.D.、Sperti,G.、Colucci,W.S.和Clapham,D.E.(1988)Circ。 第62号决议 , 1049-1054. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
2 Deisserth,K.、Heist,E.K.和Tsien,R.W.(1998)《自然》392 , 198-202. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
三。 Dolmetsch,R.E.,Pajvani,U.,Fife,K.,Spotts,J.M.&Greenberg,M.E.(2001)《科学》294 , 333-339. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
4 Nishiyama,M.、Hoshino,A.、Tsai,L.、Henley,J.R.、Goshima,Y.、Tessier-Lavigne,M.,Poo,M.和Hong,K.(2003)《自然》423 , 990-995. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
5 Huang,Y.Y.和Malenka,R.C.(1993)《神经科学杂志》。 13 , 568-576. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
6 Seki,T.、Yokoshiki,H.、Sunagawa,M.、Nakamura,M.和Sperelakis,N.(1999)《普吕格尔斯建筑》。 437 , 317-323. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
7 Urquilla,P.R.(1978)中风9 , 133-136. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
8 Jaggar,J.H.和Nelson,M.T.(2000)美国生理学杂志。 279 , C1528年至C1539年。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
9 Amberg,G.C.、Rossow,C.F.、Navedo,M.F.和Santana,L.F.(2004)《生物杂志》。 化学。 279 , 47326-47334. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
10 Knot,H.J.和Nelson,M.T.(1998)《生理学杂志》。 508 , 199-209. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
11 Fleischmann,B.K.、Murray,R.K.和Kotlikoff,M.I.(1994)Proc。 国家。 阿卡德。 科学。 美国91 , 11914-11918年。 [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
12 Stern,M.D.(1992)《生物物理学》。 期刊63 , 497-517. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
13 Amberg,G.C.和Santana,L.F.(2003),循环。 第93号决议 , 965-971. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
14 Maravall,M.、Mainen,Z.F.、Sabatini,B.L.和Svoboda,K.(2000)《生物物理学》。 约78 , 2655-2667. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
15 Cheng,H.、Song,L.S.、Shirokova,N.、Gonzalez,A.、Lakatta,E.G.、Rios,E.&Stern,M.D.(1999)《生物物理学》。 约76 , 606-617. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
16 Zenisek,D.、Davila,V.、Wan,L.和Almers,W.(2003)《神经科学杂志》。 23 , 2538-2548. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
17 Quayle,J.M.、McCarron,J.G.、Asbury,J.R.和Nelson,M.T.(1993)《美国生理学杂志》。 264 , H470-H478。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
18 Rubart,M.、Patlak,J.B.和Nelson,M.T.(1996)J.Gen.Physiol。 107 , 459-472. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
19 del Castillo,J.&Katz,B.(1954)《生理学杂志》。 124 , 560-573. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
20 Cloues,R.K.和Sather,W.A.(2000)《生理学杂志》。 524 第1部分, 19-36. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
21 Wang,S.Q.,Song,L.S.,Lakatta,E.G.&Cheng,H.(2001)《自然》第410页 , 592-596. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
22 Moore,E.D.、Voigt,T.、Kobayashi,Y.M.、Isenberg,G.、Fay,F.S.、Gallitelli,M.F.和Franzini-Amstrong,C.(2004)《生物物理学》。 J.87号 , 1836-1847. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
23 Wickman,G.、Lan,C.和Vollrath,B.(2003)巡回法院。 第92号决议 , 809-816. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
24 Cheng,H.、Lederer,W.J.和Cannell,M.B.(1993)《科学》262 , 740-744. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
25 Nelson,M.T.、Cheng,H.、Rubart,M.、Santana,L.F.、Bonev,A.D.、Knot,H.J.和Lederer,W.J.(1995)《科学》270 , 633-637. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
26 Terentyev,D.、Viatchenko-Karpinski,S.、Valdivia,H.H.、Escobar,A.L.和Gyorke,S.(2002)Circ。 第91号决议 , 414-420. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
27 Gordienko,D.V.、Bolton,T.B.和Cannell,M.B.(1998)《生理学杂志》。 507 , 707-720. [ DOI程序 ] [ PMC免费文章 ] [ 公共医学 ] [ 谷歌学者 ]
28 Hess,P.、Lansman,J.B.和Tsien,R.W.(1984)《自然》311 , 538-544. [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
29 Harder,D.R.、Gilbert,R.和Lombard,J.H.(1987)《美国生理学杂志》。 253 , F778-F781。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
30 Wellman,G.C.、Cartin,L.、Eckman,D.M.、Stevenson,A.S.、Saundry,C.M.、Lederer,W.J.和Nelson,M.T.(2001)《美国生理学杂志》。 281 , H2559-H2567。 [ DOI程序 ] [ 公共医学 ] [ 谷歌学者 ]
关联数据 本节收集本文中包含的任何数据引用、数据可用性声明或补充材料。


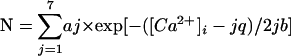
{kind=link}
{kind=link}