基于同源性的远缘细菌水平基因转移对种内多样性和分化贡献的评估 苛求木霉


摘要
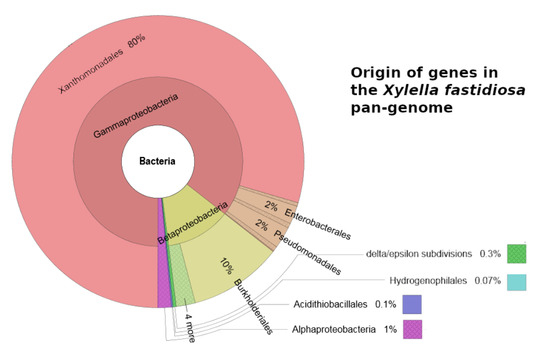
1.简介
2.结果
2.1. 矫形断言
2.2. 核心基因组分析
2.3. 泛基因组分析
2.4. 苛求小叶菌途径活性的基因组分析
2.5. 苛求小叶菌基因组中最近HGT的证据
3.讨论
3.1. BUSCO分析证实苛求小叶菌基因组减少
3.2. 与苛求二叶菌副基因组形状有关的远缘细菌
3.3. 苛求小叶菌从不同的包被中获得不同的功能
3.4. 苛求木霉和木霉液微生物组
4.材料和方法
4.1. 获取泛基因组
4.2. 根据同源性和同源性搜索对基因进行分类
4.3. 功能分配和其他分析
5.结论
补充资料
作者贡献
基金
知情同意书
数据可用性声明
致谢
利益冲突
工具书类
杰格,M。; Caffer,D。; 坎德雷斯,T。; Chatzivassiliou,E。; Dehnen-Schmutz,K。; Gilioli,G。; 格雷戈伊尔,J.C。; Jaques Miret,J.A。; MacLeod,A。; 纳瓦哈斯·纳瓦罗,M。; 等。更新的有害生物分类 苛求木霉 . 欧洲食品安全局J。 2018 , 16 . [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Janse,J.D。; 奥布拉多维奇。 苛求木霉 :其生物学、诊断、控制和风险。 《植物病理学杂志》。 2010 , 92 ,S35–S48。 [ 谷歌学者 ] Chatterjee,S。; 阿尔梅达,R.P.P。; Lindow,S.生活在两个世界:植物和昆虫的生活方式 苛求木霉 . 每年。 植物疗法评论。 2008 , 46 , 243–271. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 霍普金斯,D.L。; A.H.珀塞尔。 苛求木霉 葡萄皮尔斯病和其他紧急疾病的病因。 植物病害。 2002 , 86 , 1056–1066. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] Nunney,L。; 维克曼,D.B。; 布罗姆利,R.E。; Russell,S.A.公司。; Hartman,J.R。; 莫拉诺,L.D。; Stouthamer,R.最近的进化辐射和寄主植物的特化 苛求木霉 原产于美国的亚种。 申请。 环境。 微生物。 2013 , 79 , 2189–2200. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] 陈,J。; Jarret,R.L。; 秦,X。; Hartung,J.S。; Banks,D。; Chang,C.J。; 霍普金斯,D.L.16S rDNA序列分析 苛求木霉 菌株。 系统。 申请。 微生物。 2000 , 23 , 349–354. [ 谷歌学者 ] [ 交叉参考 ] 罗德里格斯-R,L.M。; Grajales,A。; Arrieta-Ortiz,M.L。; 萨拉查,C。; Restrepo,S.公司。; Bernal,A.该属基于基因组的系统发育 黄单胞菌属 . BMC微生物。 2012 , 12 , 43. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] 苏,C.-C。; 邓,W.-L。; 简·F·J。; Chang,C.-J。; 黄,H。; Shih,H.-T。; 陈,J。 台湾小叶藻 sp.nov.,引起梨叶枯病。 国际期刊系统。 进化。 微生物。 2016 , 66 , 4766–4771. [ 谷歌学者 ] [ 交叉参考 ] 西北部沙德。; Postnikova,E。; 莱西,G。; 法特米,M。; Chang,C.-J。 苛求木霉 亚种: 法氏X 第(b)小节。 [修正] 苛求 [校正]子程序。 11月。, 法氏X 第(b)小节。 多路复用 第(b)小节。 11月,和 法氏X 第(b)小节。 波卡 第(b)小节。 11月。 系统。 申请。 微生物。 2004 , 27 , 290–300. [ 谷歌学者 ] [ 交叉参考 ] Marcelletti,S。; 蝎属,M.多个基因组的全基因组比较和分类相关性 苛求木霉 菌株揭示了三个亚种和一个新亚种的出现 二甲苯属 物种。 架构(architecture)。 微生物。 2016 , 198 , 803–812. [ 谷歌学者 ] [ 交叉参考 ] Denancé,北卡罗来纳州。; Briand,M。; 加博里奥,R。; 盖拉德,S。; Jacques,M.-A.《遗传关系和亚种特征的鉴定》 苛求木霉 . BMC基因组。 2019 , 20 , 239. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] Scally,M。; Schuenzel,E.L。; 斯托沙默,R。; Nunney,L.植物病原菌的多焦点序列型系统 苛求木霉 重组和点突变对克隆多样性的相对贡献。 申请。 环境。 微生物。 2005 , 71 , 8491–8499. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 努尼,L。; 霍普金斯,D.L。; 莫拉诺,L.D。; S.E.罗素。; Stouthamer,R.中的亚种间重组 苛求木霉 美国本土菌株:与未成功入侵相关的新宿主感染。 申请。 环境。 微生物。 2014 , 80 , 1159–1169. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] Nunney,L。; 奥尔蒂斯,B。; Russell,S.A.公司。; Ruiz Sánchez,R。; Stouthamer,R.植物病原体的复杂生物地理学 苛求木霉 :中美洲引种和亚种渗入的遗传证据。 公共科学图书馆 2014 , 9 ,e112463。 [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 科莱塔·菲略,H.D。; 旧金山,C.S。; Lopes,J.R.S。; 穆勒,C。; Almeida,R.P.P.同源重组和 苛求木霉 南美洲的宿主-猪协会。 植物病理学 2017 , 107 , 305–312. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] 波蒂斯,N。; 坎德尔,P.P。; 梅尔法,M.V。; 无网,A.C。; Parker,J.K。; 斯坦格特区。; 阿尔梅达,R.P.P。; Bergsma-Vlami,M。; Westenberg,M。; 科宾,P.A。; 等。亚种间和亚种内同源重组的模式为 苛求木霉 . ISME期刊。 2019 , 13 , 2319–2333. [ 谷歌学者 ] [ 交叉参考 ] 卡斯蒂略,A.I。; Chacón-Díaz,C。; 罗德里格斯-穆里略,N。; 科莱塔·菲略,H.D。; Almeida,R.P.P.。当地人口历史和生态对全球传播病原体进化的影响。 BMC基因组。 2020 , 21 . [ 谷歌学者 ] [ 交叉参考 ] Pieretti,I。; 佩西奇,A。; 彼得拉斯,D。; 罗耶,M。; Suessmuth,R.D.公司。; Cociancich,S.什么使 白色黄单胞菌 在黄色单胞菌中独一无二? 前面。 植物科学。 2015 , 6 , 289. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Perez-Quintero,A.L。; 奥尔蒂兹·卡斯特罗,M。; Lang,J.M。; Rieux,A。; Wu,G。; 刘,S。; 查普曼,T.A。; Chang,C。; 齐格尔,J。; 彭,Z。; 等。新兴人群的基因组采集 血管黄单胞菌 光伏。 脉管 感染了美国和阿根廷的玉米。 植物病理学 2020 , 110 , 1161–1173. [ 谷歌学者 ] [ 交叉参考 ] Pieretti,I。; 罗耶,M。; 巴贝,V。; Carrere,S。; Koebnik,R。; Cociancich,S。; 库鲁,A。; Darrasse,A。; Gouzy,J。; 雅克,硕士。; 等 白色黄单胞菌 为木质部有限的还原基因组进化提供了新的见解 黄单胞菌科 . BMC基因组。 2009 , 10 , 616. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] 阿尔滕霍夫,A.M。; 手套,N.M。; 列车,C.-M。; Kaleb,K。; Warwick Vesztrocy,A。; Dylus博士。; 德法里亚斯,T.M。; Zile,K。; 史蒂文森,C。; Long,J。; 等。2018年OMA正畸数据库:通过更丰富的网络和编程接口检索生命所有领域之间的进化关系。 核酸研究。 2018 , 46 ,D477–D485。 [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] Kaleb,K。; Vesztrocy,A.W。; 阿尔滕霍夫。; Dessimoz,C.扩展Orthologous Matrix(OMA)编程接口:REST API和用于R和Python的OmaDB包。 F1000研究 2019 , 8 , 42. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] 西芒,F.A。; Waterhouse,R.M。; 约阿尼迪斯,P。; Kriventseva,E.V.公司。; Zdobnov,E.M.BUSCO:用单拷贝同源序列评估基因组组装和注释完整性。 生物信息学 2015 , 31 , 3210–3212. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] Medini,D。; Crabtree,J。; 莫拉,M。; 彼得森,J。; 特特林,H。; 安吉奥利,S。; 罗斯,I。; 北卡罗来纳州沃德。; 多纳蒂,C。; 斯卡塞利,M。; 等。多种致病菌株的基因组分析 无乳链球菌 :微生物“泛基因组”的含义。 程序。 国家。 阿卡德。 科学。 美国 2005 , 102 , 13950–13955. [ 谷歌学者 ] 医学博士De Carvalho。; Loreto,E.L.S.全基因组中转座元件水平转移检测方法。 遗传学。 分子生物学。 2012 , 35 , 1078–1084. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 劳伦斯,J.G。; Ochman,H.细菌基因组的改良:变化率和交换率。 《分子进化杂志》。 1997 , 44 , 383–397. [ 谷歌学者 ] [ 交叉参考 ] 维登贝克,J。; Cohan,F.M.通过水平遗传转移和适应新生态位的细菌多样性起源。 FEMS微生物。 版次。 2011 , 35 , 957–976. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Vandroemme,J。; 科廷,B。; Baeyen,S。; 德沃斯,P。; Maes,M.草拟基因组序列 芳香黄单胞菌 揭示了还原进化和不同的毒力相关基因含量。 BMC基因组。 2013 , 14 . [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Shvetsov,A.V.公司。; 列别捷夫,D.V。; Chervyakova,D.B。; 巴赫拉诺娃,I.V。; Yung,I.A。; 拉杜列斯库,A。; 库克林,A.I。; 拜廷,D.M。; Isaev-Ivanov,V.V.带有突触前RecA纤维的RecX蛋白复合物的结构:分子动力学模拟和小角中子散射。 FEBS通讯。 2014 , 588 , 948–955. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] R.T.西尔兹。; Chin,J.K。; 安德斯特区。; de Crécy-Lagard,V。; W.A.克雷格。; Romesberg,F.E.抑制突变和对抗抗生素耐药性的演变。 《公共科学图书馆·生物》。 2005 , 三 ,e176。 [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Beaber,J.W。; Hochhut,B。; Waldor,M.K.SOS反应促进抗生素耐药基因的水平传播。 自然 2004 , 427 , 72–74. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] 斯图德霍姆,D.J。; 科门,E。; 麦克莱恩,D。; Schornack,S。; Aritua,V。; Thwaites,R。; 格兰特,M。; 史密斯,J。; Jones,J.D.G.基因组测序数据揭示了香蕉中的毒力因子 黄单胞菌属 枯萎。 FEMS微生物。 莱特。 2010 , 310 , 182–192. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] R.G.伯奇。 白色黄单胞菌 以及疾病控制的抗致病途径。 分子植物病原体。 2001 , 2 , 1–11. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] 帕克,H.-J。; Jung,B。; Lee,J。; Han,S.-W.DNA甲基转移酶EadM的功能特性 轴索黄单胞菌 光伏。 甘氨酸 通过蛋白质组和表型分析。 科学。 代表。 2019 , 9 , 2446. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 拉杰什瓦里,R。; Jha,G。; Sonti,R.V.在植物中表达的木聚糖酶的作用 米黄色单胞菌 光伏。 大米 促进水稻的毒力。 分子-植物-微生物相互作用。 2005 , 18 , 830–837. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 韦塞尔,M。; Klüsener,S。; Gödeke,J。; 弗里茨,C。; Hacker,S。; Narberhaus,F.根癌农杆菌的毒力需要细菌膜中的磷脂酰胆碱。 摩尔微生物。 2006 , 62 , 906–915. [ 谷歌学者 ] [ 交叉参考 ] Farberg,A.M。; 哈特,W.K。; Johnson,R.J.来自剧毒细菌的毒性相关丝氨酸水解酶的异常底物特异性, 土拉伦弗朗西斯 . 生物化学。 生物物理学。 代表。 2016 , 7 , 415–422. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Kulasakara,H。; 李,V。; Brencic,A。; 北卡罗来纳州利伯拉蒂。; 乌尔巴赫,J。; 宫田,S。; Lee,D.G。; 尼利,A.N。; Hyodo,M。; Hayakawa,Y。; 等.分析 铜绿假单胞菌 二鸟苷酸环化酶和磷酸二酯酶揭示了双(3′-5′)环GMP在毒力中的作用。 程序。 国家。 阿卡德。 科学。 美国 2006 , 103 , 2839–2844. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] 秋田,H。; 木村,Z.I。; Hoshino,T。 腐殖假单胞菌 11月,从叶土中分离得到。 架构(architecture)。 微生物。 2019 , 201 , 245–251. [ 谷歌学者 ] [ 交叉参考 ] Bai,Y。; 米勒,D.B。; 斯里尼瓦斯,G。; 加里多·奥特(Garrido-Oter,R.)。; 波托夫,E。; 罗特,M。; Dombrowski,N。; 慕尼黑,P.C。; Spaepen,S。; Remus-Emsermann,M。; 等 拟南芥 叶和根微生物群。 自然 2015 , 528 , 364–369. [ 谷歌学者 ] [ 交叉参考 ] Hayes,F.毒素-抗毒素:质粒维持、程序性细胞死亡和细胞周期阻滞。 科学类 2003 , 301 , 1496–1499. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] M.W.李。; Tan,C.C。; 罗杰斯,E.E。; Stenger,D.C.毒素-抗毒素系统mqsR/ygiT和dinJ/relE 苛求木霉 . 生理学。 分子植物病原体。 2014 , 87 , 59–68. [ 谷歌学者 ] [ 交叉参考 ] Kaneko,T。; Y.中村。; 佐藤,S。; Asamizu,E。; 加藤,T。; Sasamoto,S。; 渡边,A。; Idesawa,K。; 石川,A。; 川岛,K。; 等.充氮共生细菌的完整基因组结构 洛蒂中根瘤菌 . DNA研究。 2000 , 7 , 331–338. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 雷迪,J.D。; Reddy,S.L。; 霍普金斯,D.L。; Gabriel,D.W.TolC对以下疾病的致病性是必需的 苛求木霉 在葡萄藤中。 分子-植物-微生物相互作用。 2007 , 20 , 403–410. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 吉尔哈贝特,M.R。; 不列颠哥伦比亚省柯克帕特里克 苛求木霉 抗病毒基因:血凝素粘附素有助于生物膜成熟 法氏X 以及定植和减弱毒力。 分子植物微生物相互作用。 2005 , 18 , 856–868. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] Van Sluys,文学硕士。; de Oliveira,医学博士。; Monteiro-Vitorello,C.B。; 宫崎骏,C.Y。; 弗兰,L.R。; 洛杉矶卡马戈。; 达席尔瓦,A.C.R。; Moon,D.H。; Takita,文学硕士。; Lemos,E.G.M。; 等。Pierce’s病和柑橘杂色黄化病株的完整基因组序列的比较分析 苛求木霉 . 《细菌学杂志》。 2003 , 185 , 1018–1026. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 罗珀,M.C。; 格雷夫,L.C。; 沃伦,J.G。; 拉巴维奇,J.M。; 不列颠哥伦比亚省柯克帕特里克。 苛求木霉 需要多聚半乳糖醛酸酶才能在 葡萄 葡萄藤。 分子-植物-微生物相互作用。 2007 , 20 , 411–419. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Purcell,A.H.公司。; 霍普金斯,D.L.苦味木质部限制细菌植物病原体。 每年。 植物疗法评论。 1996 , 34 , 131–151. [ 谷歌学者 ] [ 交叉参考 ] 阿查里,G.A。; Ramesh,R.茄科作物木质部驻留细菌的多样性、生物控制和植物生长促进能力。 国际微生物杂志。 2014 . [ 谷歌学者 ] [ 交叉参考 ] 詹姆斯,E.K。; Gyaneshwar,P。; Mathan,N。; 巴拉圭,W.L。; 下午Reddy。; 伊安内塔,P.P.M。; 奥利瓦雷斯,F.L。; Ladha,J.K.植物生长繁殖细菌对水稻幼苗的感染和定殖 血吸虫 Z67号。 分子-植物-微生物相互作用。 2002 , 15 , 894–906. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Compant,S.公司。; Reiter,B。; Sessitsch,A。; Nowak,J。; Clément,C。; Barka,E.A.内生定殖 葡萄 由植物生长繁殖细菌引起 伯克氏菌属 sp.菌株PsJN。 申请。 环境。 微生物。 2005 , 71 , 1685–1693. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] Compant,S。; 卡普兰,H。; Sessitsch,A。; Nowak,J。; 艾特·巴尔卡(Ait Barka),E。; Clément,C.内生定殖 葡萄 L.依据 伯克霍尔德菌 菌株PsJN:从根际到花序组织。 FEMS微生物。 经济。 2008 , 63 , 84–93. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 拉卡瓦,P.T。; 阿泽维多,J.L。; 米勒,T.A。; Hartung,J.S.的相互作用 苛求木霉 柑橘内生细菌研究进展。 树用于。 科学。 生物技术。 2009 , 三 , 40–48. [ 谷歌学者 ] Deyett,E。; Rolshausen,P.E.高皮尔斯病压力下葡萄树液微生物组的时间动力学。 前面。 植物科学。 2019 , 10 . [ 谷歌学者 ] [ 交叉参考 ] Kung,S.H。; 无网,A.C。; Kwan,J.Y。; Almeida,R.P.P.DNA大小对转基因和重组效率的影响 苛求木霉 . 申请。 环境。 微生物。 2013 , 79 , 1712–1717. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] 范霍夫,M。; 无网,A.C。; Sicard,A。; Rieux,A.等人。; 科莱塔·菲略,H.D。; de La Fuente,L.公司。; 斯坦格特区。; Almeida,R.P.P.基因组多样性和重组 苛求木霉 亚种。 申请。 环境。 微生物。 2019 , 85 ,e02972-18。 [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] Larkin,文学硕士。; Blackshields,G。; 棕色,N.P。; Chenna,R。; McGettigan,P.A。; McWilliam,H。; 瓦伦丁,F。; I.M.华莱士。; Wilm,A。; 洛佩兹,R。; 等。Clustal W和Clustal X版本2.0。 生物信息学 2007 , 23 , 2947–2948. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Wasukira,A。; Tayebwa,J。; Thwaites,R。; Paszkiewicz,K。; Aritua,V.公司。; 库比里巴,J。; 史密斯,J。; 格兰特,M。; Studholme,D.J.全基因组测序揭示了遗传单型病原体的两个主要亚系 campestris黄单胞菌 病理变种 马齿苋属 . 基因 2012 , 三 , 361–377. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] 马格洛特,D。; 奥斯特尔,J。; K.D.普鲁特。; Tatusova,T.Entrez基因:NCBI以基因为中心的信息。 核酸研究。 2011 , 39 ,D52–D57。 [ 谷歌学者 ] [ 交叉参考 ] 阿尔滕霍夫,A.M。; 吉尔,M。; Gonnet,G.H。; Dessimoz,C.从同源基因对推断等级同源群。 公共科学图书馆 2013 , 8 ,e53786。 [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] Deorowicz,S。; Debudaj-Grabysz,A。; Gudys,A.FAMSA:巨大蛋白质家族的快速准确多序列比对。 科学。 代表。 2016 , 6 , 33964. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] 韦尔塔·塞帕斯,J。; 塞拉,F。; Bork,P.ETE 3:系统发育数据的重建、分析和可视化。 分子生物学。 进化。 2016 , 33 , 1635–1638. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] Ondov,B.D。; 新罕布什尔州伯格曼。; Phillippy,A.M.网络浏览器中的交互式基因组可视化。 BMC生物信息。 2011 , 12 , 385. [ 谷歌学者 ] [ 交叉参考 ] [ 公共医学 ] [ 绿色版本 ] 森雅,Y。; 伊藤,M。; Okuda,S。; 吉泽,A.C。; Kanehisa,M.KAAS:自动基因组注释和路径重建服务器。 核酸研究。 2007 , 35 ,W182–W185。 [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] 三通,H。; Waldmann,J。; 伦巴多特,T。; Bauer,M。; Glöckner,F.O.TETRA:一个用于分析和比较DNA序列中四核苷酸使用模式的网络服务和独立程序。 BMC生物信息。 2004 , 5 , 163. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ]




{kind=link}
{kind=link}
{kind=link}
{kind=link}