膜结合货物的细胞内贩运:一个小舰队的观点


摘要
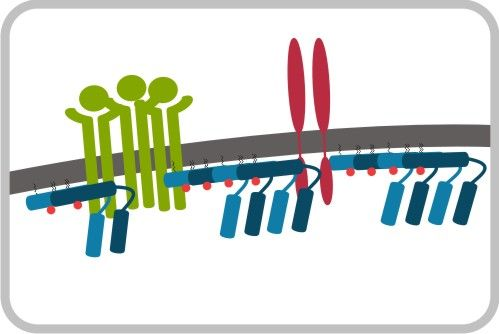
1.脂质微结构域与内分泌
2.Flotillin蛋白家族
3.推测的血小板依赖性内吞途径的发现

{kind=link}
{kind=link}
{kind=link}
4.依赖船队的货物贩运和分拣:超越细胞内吞
4.1. 小群菌在内切体内的事件分类中的作用
4.2. Flotillins在细胞内吞中的间接作用:细胞质膜上的内吞前聚集

5.在信号传递过程中作为内吞货物的小鱼群
6.新时代:从Flotillin依赖到Flotillin辅助的内分泌
7.结论:Flotillins在膜贩运中的应用——获得更大的前景

致谢
作者贡献
利益冲突
参考文献
布朗,D.A。; Rose,J.K.,在转运至顶端细胞表面期间,GPI锚定蛋白到富含糖脂的膜亚结构域的分类。 单元格 1992 , 68 , 533–544. [ 谷歌学者 ] 西蒙斯,K。; Ikonen,E.细胞膜中的功能筏。 自然 1997 , 387 , 569–572. [ 谷歌学者 ] [ 交叉参考 ] 西蒙斯,K。; Sampaio,J.L.膜组织和脂筏。 冷泉港。 透视。 生物。 2011 , 三 . [ 谷歌学者 ] [ 交叉参考 ] D.布朗。; Waneck,G.L.糖基-磷脂酰肌醇锚定膜蛋白。 《美国肾脏学会杂志》。 1992 , 三 , 895–906. [ 谷歌学者 ] 上午Fra。; 威廉姆森,E。; 西蒙斯,K。; Parton,R.G.洗涤剂淋巴细胞中不含小窝的不溶性糖脂微区。 生物学杂志。 化学。 1994 , 269 , 30745–30748. [ 谷歌学者 ] Sargiacomo,M。; 苏多尔,M。; 唐,Z。; Lisanti,M.P.信号转导分子和糖基磷脂酰肌醇连接蛋白在MDCK细胞中形成富含小窝蛋白的不溶性复合物。 《细胞生物学杂志》。 1993 , 122 , 789–807. [ 谷歌学者 ] [ 交叉参考 ] Resh,M.D.Src家族成员的肉豆蔻酰化和棕榈酰化:物质的脂肪。 单元格 1994 , 76 , 411–413. [ 谷歌学者 ] [ 交叉参考 ] Casey,P.J.细胞信号中的蛋白质脂质化。 科学类 1995 , 268 , 221–225. [ 谷歌学者 ] Levental,I。; Grzybek,M。; Simons,K。润滑方式:脂质修饰决定蛋白质与膜筏的结合。 生物化学 2010 , 49 , 6305–6316. [ 谷歌学者 ] [ 交叉参考 ] 迪特里希,C。; Volovyk,Z.N。; 列维,M。; N.L.汤普森。; Jacobson,K.将Thy-1、GM1和交联磷脂类似物分配到支撑模型膜单层中重建的脂筏中。 程序。 国家。 阿卡德。 科学。 美国 2001 , 98 , 10642–10647. [ 谷歌学者 ] Neumann-Giesen,C。; Falkenbach,B。; 贝希特,P。; 克拉森,S。; 卢尔斯,G。; Stuermer,首席执行官。; 赫尔佐格,V。; Tikkanen,R.reggie-1/flotillin-2的膜和筏结合:肉豆蔻酰化、棕榈酰化和寡聚作用以及通过过度表达诱导丝足类。 生物化学。 J。 2004 , 378 , 509–518. [ 谷歌学者 ] [ 交叉参考 ] 巴布克,T。; Ruonala,M。; 梅斯特,M。; 阿马迪,M。; Genzler,C。; 埃斯波西托,A。; Tikkanen,R.reggie-1/flotillin-2和reggie-2/flotillin-1的异构化是其内吞所必需的。 细胞信号。 2009 , 21 , 1287–1297. [ 谷歌学者 ] [ 交叉参考 ] 唐纳森,J.G。; Porat-Shliom,N。; Cohen,L.A.Clathrin非依赖性内吞作用:细胞信号和PM重塑的独特平台。 细胞信号。 2009 , 21 , 1–6. [ 谷歌学者 ] 多尔蒂,G.J。; McMahon,H.T.内吞作用机制。 生物化学年度修订。 2009 , 78 , 857–902. [ 谷歌学者 ] [ 交叉参考 ] H.T.麦克马洪。; Boucrot,E.氯氰菊酯介导的内吞作用的分子机制和生理功能。 自然修订版分子细胞生物学。 2011 , 12 , 517–533. [ 谷歌学者 ] [ 交叉参考 ] Sandvig,K。; 普斯特,S。; 斯科特兰,T。; van Deurs,B.氯氰菊酯非依赖性内吞作用:机制和功能。 货币。 操作。 细胞生物学。 2011 , 23 , 413–420. [ 谷歌学者 ] [ 交叉参考 ] 帕顿,R.G。; del Pozo,M.A.Caveolae担任质膜传感器、保护器和组织者。 自然修订版分子细胞生物学。 2013 , 14 , 98–112. [ 谷歌学者 ] [ 交叉参考 ] 亨利·J·R。; 克鲁格,E.W。; 奥斯瓦尔德,B.J。; McNiven,M.A.Dynamin介导的小窝内化。 《细胞生物学杂志》。 1998 , 141 , 85–99. [ 谷歌学者 ] [ 交叉参考 ] 哦,P。; 麦金托什,民主党人。; 位于小窝颈部的Schnitzer,J.E.Dynamin通过GTP驱动内皮细胞质膜的裂变,介导其发芽形成运输囊泡。 《细胞生物学杂志》。 1998 , 141 , 101–114. [ 谷歌学者 ] [ 交叉参考 ] 格里波夫,O.O。; 布赖特,N.A。; Nichols,B.J.Flotillin-1定义了哺乳动物细胞中不依赖于网格蛋白的内吞途径。 自然细胞生物学。 2006 , 8 , 46–54. [ 谷歌学者 ] [ 交叉参考 ] 弗里克,M。; 布赖特,N.A。; Riento,K。; 布雷,A。; Merrified,C。; Nichols,B.J.浮萍的联合诱导膜微区的形成、膜弯曲和囊泡出芽。 货币。 生物。 2007 , 17 , 1151–1156. [ 谷歌学者 ] [ 交叉参考 ] 梅斯特,M。; Zuk,A。; Tikkanen,R.动力蛋白和甲氰菊酯在船队细胞贩运中的作用。 FEBS J公司。 2014 , 281 , 2956–2976. [ 谷歌学者 ] [ 交叉参考 ] 奥托,G.P。; Nichols,B.J.《船队微域的作用——内吞作用及其他》。 细胞科学杂志。 2012 , 124 , 3933–3940. [ 谷歌学者 ] [ 交叉参考 ] 莫罗,I.C。; 里昂,S。; 马丁,S。; Prior,I.A。; 普罗哈斯卡,R。; Hancock,J.F。; D.E.詹姆斯。; Parton,R.G.Flotillin-1/reggie-2通过一条新的高尔基诱导依赖路径将流量输送至水面筏域。 一种新型膜靶向结构域的鉴定及其在棕榈酰化中的作用。 生物学杂志。 化学。 2002 , 277 , 48834–48841. [ 谷歌学者 ] [ 交叉参考 ] 李毅。; B.R.马丁。; 克雷瓦特,B.F。; Hofmann,S.L.DHHC5蛋白棕榈酰化物flotillin-2,在培养细胞中诱导神经元分化时迅速降解。 生物学杂志。 化学。 2012 , 287 , 523–530. [ 谷歌学者 ] [ 交叉参考 ] 索利斯,G.P。; Hoegg,M。; 蒙德洛,C。; 施洛克,Y。; Malaga-Trillo,E。; Rivera Milla,E。; Stuermer,C.A.Reggie/flotillin蛋白在膜微结构域中被组织成稳定的四聚体。 生物化学。 J。 2007 , 403 , 313–322. [ 谷歌学者 ] [ 交叉参考 ] 斯特劳斯,K。; Goebel,C。; Runz,H。; 莫比乌斯,W。; Weiss,S。; 费斯纳,I。; 西蒙斯,M。; Schneider,A.外显子分泌改善Niemann-Pick C型疾病中胆固醇的溶酶体储存。 生物学杂志。 化学。 2010 , 285 , 26279–26288. [ 谷歌学者 ] 比克尔,P.E。; 谢勒,P.E。; Schnitzer,J.E。; 哦,P。; Lisanti,医学博士。; Lodish、H.F.Flotillin和表皮表面抗原定义了一个新的小窝相关整合膜蛋白家族。 生物学杂志。 化学。 1997 , 272 , 13793–13802. [ 谷歌学者 ] 沃龙特,D。; Galbiati,F。; 李,S。; 西山,K。; 冈本,T。; Lisanti,M.P.Flotillins/cavatellins在细胞和组织中有差异表达,并与小窝蛋白形成异寡聚体复合物 体内试验。 新型flotillin-1单克隆抗体探针的表征和表位映射。 生物学杂志。 化学。 1999 , 274 , 12702–12709. [ 谷歌学者 ] 费诺,I。; 结冰,A。; Tikkanen,R.Reggie-1和Reggie-2定位于上皮细胞的非小窝筏:细胞定位不依赖小窝蛋白的表达。 《欧洲细胞生物学杂志》。 2007 , 86 , 345–352. [ 谷歌学者 ] [ 交叉参考 ] 刘杰。; Deyoung,S.M。; 张,M。; 多尔德,L.H。; Saltiel,A.R.flotillin-1的气孔蛋白/抑制素/flotillin/HflK/C结构域包含指导3T3-L1脂肪细胞质膜定位和蛋白质相互作用的独特序列。 生物学杂志。 化学。 2005 , 280 , 16125–16134. [ 谷歌学者 ] [ 交叉参考 ] Neumann-Giesen,C。; 费诺,I。; 阿马迪,M。; Tikkanen,R.表皮生长因子诱导的reggie-1/flotilin-2酪氨酸磷酸化在细胞扩散和肌动蛋白细胞骨架信号传导中的作用。 细胞科学杂志。 2007 , 120 , 395–406. [ 谷歌学者 ] [ 交叉参考 ] Riento,K。; 弗里克,M。; 谢弗,I。; Nichols,B.J.flotillin-1和flotillin-2的细胞内吞受Fyn激酶调节。 细胞科学杂志。 2009 , 122 , 912–918. [ 谷歌学者 ] 北卡罗来纳州库尔勒。; 约翰·B。; 梅斯特,M。; Tikkanen,R.Flotillins在受体酪氨酸激酶信号传递和胞内作用中的作用:酪氨酸磷酸化和寡聚作用。 在 蛋白质磷酸化与人类健康 ; Huang,C.编辑。; InTech出版社:克罗地亚里耶卡,2012年。 [ 谷歌学者 ] 埃德加,A.J。; Polak,J.M.Flotillin-1:基因结构:人肺CDNA克隆和替代多聚腺苷酸化信号的鉴定。 国际生物化学杂志。 细胞生物学。 2001 , 33 , 53–64. [ 谷歌学者 ] Rivera Milla,E。; Stuermer,C.A。; Malaga-Trillo,E.reggie(flotillin)、reggie-like和其他脂质转运蛋白的古老起源:SPFH结构域的聚合进化。 细胞分子生命科学。 2006 , 63 , 343–357. [ 谷歌学者 ] 阿马迪,M。; 梅斯特,M。; Banning,A。; 托马索维奇,A。; J.穆兹。; 拉贾林加姆,K。; Tikkanen,R.Flotillin-1/reggie-2蛋白在受体酪氨酸激酶/有丝分裂原活化蛋白激酶信号的激活中发挥双重作用。 生物学杂志。 化学。 2012 , 287 , 7265–7278. [ 谷歌学者 ] Banning,A。; 雷根布雷希特,C.R。; Tikkanen,R.在flotillin-2基因敲除小鼠模型中丝裂原活化蛋白激酶途径活性增加。 细胞信号。 2014 , 26 , 198–207. [ 谷歌学者 ] Banning,A。; 北卡罗来纳州库尔勒。; 梅斯特,M。; Tikkanen,R.Flotilins在受体酪氨酸激酶信号传导和癌症中的作用。 细胞 2014 , 三 ,129–149。 [ 谷歌学者 ] 梅斯特,M。; 托马索维奇,A。; Banning,A。; Tikkanen,R.有丝分裂原活化蛋白(MAP)激酶支架蛋白:综述。 国际分子科学杂志。 2013 , 14 , 4854–4884. [ 谷歌学者 ] [ 交叉参考 ] Stuermer,C.A。; Lang,D.M。; Kirsch,F。; 韦切尔斯,M。; Deininger,S.O.公司。; Plattner,H.糖基磷脂酰肌醇锚定蛋白和fyn激酶在reggie-1和-2定义的非泡状质膜微域中组装。 分子生物学。 单元格 2001 , 12 , 3031–3045. [ 谷歌学者 ] [ 交叉参考 ] 索利斯,G.P。; 北卡罗来纳州赫尔斯堡。; 氡,Y。; Katanev,V.L。; 普拉特纳,H。; Stuermer,C.A.Reggies/flotillins在小管上皮细胞循环室与Rab11a和SNX4相互作用,并在转铁蛋白受体和E-cadherin转运中发挥作用。 分子生物学。 单元格 2013 , 24 , 2689–2702. [ 谷歌学者 ] Gagescu,R。; 德马雷克斯,N。; 帕顿,R.G。; Hunziker,W。; 洛杉矶Huber。; Gruenberg,J.马丁·达比犬肾细胞的再循环内体是一个富含筏成分的弱酸性隔间。 分子生物学。 单元格 2000 , 11 , 2775–2791. [ 谷歌学者 ] de Gassart,A。; 基米纳德,C。; Fevrier,B。; 拉波索,G。; Vidal,M.外泌体中脂筏相关蛋白的分类。 血液 2003 , 102 , 4336–4344. [ 谷歌学者 ] [ 交叉参考 ] Dermine,J.F。; Duclos,S。; 加林,J。; St-Louis,F。; 里昂,S。; 帕顿,R.G。; Desjardins,M.Flotillin-1富集的脂筏结构域在成熟的吞噬体上积累。 生物学杂志。 化学。 2001 , 276 , 18507–18512. [ 谷歌学者 ] Phuyal,S。; 北卡罗来纳州赫斯维克。; 斯科特兰,T。; Sandvig,K。; Llorente,A.鞘糖脂和船队对胞外体释放的调节。 FEBS J公司。 2014 , 281 , 2214–2227. [ 谷歌学者 ] 托马索维奇,A。; 特劳布,S。; Tikkanen,R.FGF信号传导中的分子网络:Flotillin-1和cbl-相关蛋白竞争与成纤维细胞生长因子受体底物2的结合。 公共科学图书馆 2012 , 7 ,e29739。 [ 谷歌学者 ] [ 交叉参考 ] 维考特伦,D。; Piest,M。; 范德阿,L.J。; Al Soraj,M。; A.T.琼斯。; Engbersen,J.F。; de Smedt,南卡罗来纳州。; Braeckmans,K.Flotillin依赖性内吞作用和细胞内吞二硫基多胺/DNA多聚酶的类吞噬机制。 生物材料 2011 , 32 , 3072–3084. [ 谷歌学者 ] 佩恩,C.K。; Jones,S.A.公司。; 陈,C。; 庄,X.细胞表面蛋白聚糖和蛋白聚糖结合配体的内化和贩运。 交通 2007 , 8 , 389–401. [ 谷歌学者 ] Ge,L。; 齐,W。; Wang,L.J。; Miao,H.H。; 曲,Y.X。; Li,B.L。; Song,B.L.Flotillins在Niemann-Pick C1样1介导的胆固醇摄取中发挥着重要作用。 程序。 国家。 阿卡德。 科学。 美国 2011 , 108 , 551–556. [ 谷歌学者 ] 福特,M.G。; 皮尔斯,B.M。; 希金斯,M.K。; 瓦利斯,Y。; D.J.欧文。; A.吉布森。; 霍普金斯,C.R。; P.R.埃文斯。; McMahon,H.T.在膜上网格蛋白晶格的成核过程中,通过AP180同时结合PtdIn(4,5)P2和网格蛋白。 科学类 2001 , 291 ,1051–1055。 [ 谷歌学者 ] [ 交叉参考 ] Al Soraj,M。; He,L。; Peynshaert,K。; 库塞尔特,J。; 维考特伦,D。; Braeckmans,K。; de Smedt,S.C.公司。; Jones,A.T.siRNA和内吞途径的药理抑制,以表征大胞饮作用和肌动蛋白细胞骨架对细胞摄取葡聚糖和阳离子细胞穿透肽八精氨酸(R8)和HIV-Tat的不同作用。 J.控制。 发布 2012 , 161 , 132–141. [ 谷歌学者 ] Hansen,G.H。; Dalskov,S.M。; 拉斯穆森,C.R。; Immerdal,L。; 尼尔斯·克里斯蒂安森,L.L。; Danielsen,E.M.霍乱毒素通过脂筏和网格蛋白依赖机制进入猪肠细胞。 生物化学 2005 , 44 , 873–882. [ 谷歌学者 ] Nichols,B.J.含有脂筏的GM1在网格蛋白包被的凹坑内被耗尽。 货币。 生物。 2003 , 13 , 686–690. [ 谷歌学者 ] Kusumi,A。; 小山本田,I。; 铃木,K.从小型不稳定稳定筏中创建刺激诱导稳定筏的分子动力学和相互作用。 交通 2004 , 5 , 213–230. [ 谷歌学者 ] [ 交叉参考 ] Brameshuber,M。; 韦胡贝尔,J。; 鲁普雷希特,V。; 贡博斯,I。; Horvath,I。; 维格·L。; 埃克斯托弗,P。; 亲吻,E。; Stockinger,H。; Schutz,G.J.活细胞质膜中移动长寿命纳米平台的成像。 生物学杂志。 化学。 2010 , 285 , 41765–41771. [ 谷歌学者 ] [ 交叉参考 ] 施耐德,A。; Rajendran,L。; Honsho先生。; 格拉尔,M。; 唐纳特,G。; 沃特斯,F。; 南威尔士赫尔。; Simons,M.Flotillin,淀粉样前体蛋白的依赖性聚集调节其在神经元中的内吞作用和淀粉样蛋白生成过程。 神经科学杂志。 2008 , 28 , 2874–2882. [ 谷歌学者 ] 克雷莫纳,M.L。; 马蒂斯,H.J。; Pau,K。; 鲍顿,E。; 速度,N。; 琵琶,B.J。; 安德森,M。; Sen,N。; Robertson,S.D。; R.A.沃恩。; 等。 Flotillin-1对于PKC触发的DAT的内吞作用和膜微结构域定位至关重要。 自然神经科学。 2011 , 14 , 469–477. [ 谷歌学者 ] [ 交叉参考 ] Gorgens,A。; 贝克曼,J。; 路德维希,A.K。; 莫尔曼,M。; Durig,J。; 霍恩,P.A。; Rajendran,L。; Giebel,B.脂筏再分配和形态细胞极化是可分离的过程,为造血干细胞和祖细胞迁移提供了基础。 国际生物化学杂志。 细胞生物学。 2012 , 44 , 1121–1132. [ 谷歌学者 ] [ 交叉参考 ] Rajendran,L。; 贝克曼,J。; 马吉诺,A。; Boneberg,E.M。; Gaus,K。; 维奥拉,A。; Giebel,B。; Illges,H.Flotilins参与原始和成熟造血细胞的极化。 公共科学图书馆 2009 , 4 ,电话8290。 [ 谷歌学者 ] Rajendran,L。; 马萨拉马尼,M。; 所罗门,S。; Tikkanen,R。; Stuermer,C.A。; 普拉特纳,H。; Illges,H。在预装配平台中,船队/reggies的不对称定位赋予造血细胞固有的极性。 程序。 国家。 阿卡德。 科学。 美国 2003 , 100 , 8241–8246. [ 谷歌学者 ] Langhorst,M.F。; 路透社,A。; 卢森霍夫,G。; Boneberg,E.M。; 莱格勒,D.F。; 普拉特纳,H。; Stuermer,C.A.预制reggie/flotillin帽:用于T细胞中大结构域组装的稳定启动平台。 美国财务会计准则委员会J。 2006 , 20 , 711–713. [ 谷歌学者 ] 外遇者,S。; 马蒂内利,S。; 哈恩,J。; 罗西,J。; Niggli,V.趋化因子刺激的人类T淋巴细胞中的flotillins的动态重组。 BMC细胞生物学。 2011 , 12 , 28. [ 谷歌学者 ] [ 交叉参考 ] [ 绿色版本 ] 罗西,J。; Schlicht,D。; Engelhardt,B。; Niggli,V.Flotillins与中性粒细胞中的PSGL-1相互作用,在刺激后迅速组织成膜域,随后在尿足类动物中积累。 公共科学图书馆 2009 , 4 ,e5403。 [ 谷歌学者 ] 路德维希,A。; 奥托,G.P。; Riento,K。; E.哈姆斯。; 法伦,P.G。; Nichols,B.J.Flotillin微域与皮质细胞骨架相互作用,以控制尿足类的形成和中性粒细胞的补充。 《细胞生物学杂志》。 2010 , 191 , 771–781. [ 谷歌学者 ] [ 交叉参考 ] Mathivanan,S。; Simpson,R.J.ExoCarta:外体蛋白质和RNA简编。 蛋白质组学 2009 , 9 , 4997–5000. [ 谷歌学者 ] Saslowsky,D.E。; 赵,J.A。; 钦纳彭,H。; 马索尔,R.H。; Chinnapen,D.J。; Wagner,J.S。; de Luca,H.E。; Kam,W。; 爪,B.H。; Lencer,W.I.霍乱毒素对斑马鱼和哺乳动物细胞的毒性取决于flotillin/reggie蛋白,而不是Derlin-1或-2。 临床杂志。 投资。 2010 , 120 , 4399–4409. [ 谷歌学者 ] [ 交叉参考 ] 普斯特,S。; 戴夫,A.B。; 托格森,M.L。; van Deurs,B。; Sandvig,K.毒素运输和船队定位之间的相互作用。 公共科学图书馆 2010 , 5 ,e8844。 [ 谷歌学者 ] 约翰,文学学士。; 梅斯特,M。; 班宁,A。; Tikkanen,R.Flotillins与β-位点淀粉样前体蛋白切割酶1的二亮氨酸排序基序结合,并影响其内体排序。 FEBS J公司。 2014 , 281 , 2074–2087. [ 谷歌学者 ] [ 交叉参考 ] 拉波索,G。; Stoorvogel,W.细胞外小泡:外泌体、微泡和朋友。 《细胞生物学杂志》。 2013 , 200 , 373–383. [ 谷歌学者 ] [ 交叉参考 ] 拉波索,G。; H.W.奈曼。; 斯托沃格尔,W。; Liejendekker,R。; C.V.哈丁。; 梅利夫,C.J。; Geuze,H.J.B淋巴细胞分泌抗原提呈小泡。 实验医学学报 1996 , 183 , 1161–1172. [ 谷歌学者 ] [ 交叉参考 ] 斯塔巴赫,S。; 拉扎维,H。; Hanisch,F.G.人乳腺癌细胞MCF-7的质膜和外泌体中含有MUC1的脂筏的蛋白质组学。 蛋白质组学 2009 , 9 , 2820–2835. [ 谷歌学者 ] Baietti,M.F。; 张,Z。; 莫蒂埃,E。; Melchior,A。; 德盖斯特,G。; Geeraerts,A。; Ivarsson,Y。; Depoortere,F。; 库曼斯,C。; Vermeiren,E。; 齐默尔曼,P。; David,G.Syndecan-syntenin-ALIX调节外显体的生物发生。 自然细胞。 生物。 2012 , 14 , 677–685. [ 谷歌学者 ] [ 交叉参考 ] 何,X。; Chang,W.P。; Koelsch,G。; Tang,J.Memapsin 2(β-分泌酶)胞浆结构域与GGA1和GGA2的VHS结构域结合:对Memapsin2内吞机制的影响。 FEBS通讯。 2002 , 524 , 183–187. [ 谷歌学者 ] [ 交叉参考 ] 巴斯托里诺,L。; 艾金,A.F。; A.C.奈恩。; Pursnani,A。; Buxbaum,J.D.BACE的羧基末端包含一个分拣信号,该信号调节BACE的运输,但不调节总a(β)的形成。 分子细胞神经科学。 2002 , 19 , 175–185. [ 谷歌学者 ] Walter,J。; 弗勒,R。; Hartung,B。; 威廉,M。; Kaether,C。; Capell,A。; Lammich,S。; Multhaup,G。; Haass,C.磷酸化调节β-分泌酶的细胞内转运。 生物学杂志。 化学。 2001 , 276 , 14634–14641. [ 谷歌学者 ] [ 交叉参考 ] Kang,E.L。; 卡梅隆,A.N。; F.广场。; Walker,K.R。; Tesco,G.泛素调节GGA3介导的BACE1降解。 生物学杂志。 化学。 2010 , 285 , 24108–24119. [ 谷歌学者 ] 特易购,G。; Koh,Y.H。; Kang,E.L。; 卡梅隆,A.N。; 达斯,S。; Sena-Esteves,M。; 希尔图宁,M。; Yang,S.H.公司。; 钟,Z。; 沈毅。; 等。 GGA3的耗尽可稳定BACE并增强β-分泌酶活性。 神经元 2007 , 54 , 721–737. [ 谷歌学者 ] [ 交叉参考 ] 索利斯,G.P。; 施洛克,Y。; 北卡罗来纳州赫尔斯堡。; 韦切尔斯,M。; 普拉特纳,H。; Stuemer,C.A.Reggies/flotillins通过影响EGFR贩运来调节E-cadherin介导的细胞接触形成。 分子生物学。 单元格 2012 , 23 , 1812–1825. [ 谷歌学者 ] 索基纳,T。; Caltagarone,J。; Sorkin,A.Flotillins调节多巴胺转运体的膜流动性,但不需要其蛋白激酶C依赖的内吞作用。 交通 2013 , 14 , 709–724. [ 谷歌学者 ] 古,E.H。; Squazzo,S.L.淀粉样β蛋白的产生和释放涉及内吞途径的证据。 生物学杂志。 化学。 1994 , 269 ,17386–17389页。 [ 谷歌学者 ] 古,E.H。; Squazzo,S.L.公司。; D.J.塞尔科。; Koo,C.H.细胞表面淀粉样β蛋白前体的贩运。 标记单克隆抗体检测到的分泌、内吞和再循环。 细胞科学杂志。 1996 , 109 , 991–998. [ 谷歌学者 ] 佩雷斯,R.G。; Soriano,S。; Hayes,J.D。; 奥斯塔舍夫斯基,B。; 夏,W。; D.J.塞尔科。; 陈,X。; 斯托金,G.B。; Koo,E.H.突变识别了β-淀粉样前体蛋白内吞、周转和分泌片段生成的新信号,包括Abeta42。 生物学杂志。 化学。 1999 , 274 , 18851–18856. [ 谷歌学者 ] [ 交叉参考 ] 索基纳,T。; B.R.胡佛。; 新泽西州扎尼塞。; Sorkin,A.组分和蛋白激酶C诱导的多巴胺转运体内化是由氯菊酯依赖机制介导的。 交通 2005 , 6 ,157–170。 [ 谷歌学者 ] [ 交叉参考 ] 西格斯蒙德,S。; Argenzio,E。; 托索尼,D。; 卡瓦拉罗,E。; 马球,S。; di Fiore,P.P.氯氰菊酯介导的内化对于持续的EGFR信号传导至关重要,但对于降解来说是不必要的。 开发单元 2008 , 15 , 209–219. [ 谷歌学者 ] 西吉斯蒙德,S。; Woelk,T。; 普里,C。; 马斯佩罗,E。; 塔切蒂,C。; Transidico,P。; 迪菲奥雷,P.P。; Polo,S.对泛素化货物的非依赖性内吞作用。 程序。 国家。 阿卡德。 科学。 美国 2005 , 102 , 2760–2765. [ 谷歌学者 ] [ 交叉参考 ] 普里,C。; 托索尼,D。; 科迈,R。; 拉贝利诺,A。; Segat,D。; 卡内娃,F。; Luzzi,P。; 迪菲奥雷,P.P。; Tacchetti,C.表皮生长因子受体信号传导能力和胞内竞争性膜微域之间的关系。 分子生物学。 单元格 2005 , 16 , 2704–2718. [ 谷歌学者 ] [ 交叉参考 ] Roitbak,T。; 沃德,C.J。; 哈里斯,P.C。; 巴卡拉奥,R。; Ness,S.A.公司。; Wandinger-Ness,A.多囊肾病细胞中的多囊蛋白-1多蛋白复合物被破坏。 分子生物学。 单元格 2004 , 15 , 1334–1346. [ 谷歌学者 ] Ge,L。; Wang,J。; 齐,W。; Miao,H.H。; 曹,J。; 曲,Y.X。; Li,B.L。; Song,B.L.胆固醇吸收抑制剂ezetimibe通过阻止固醇诱导的NPC1L1内化发挥作用。 单元格元数据。 2008 , 7 , 508–519. [ 谷歌学者 ] [ 交叉参考 ] Abrami,L。; Bischofberger,M。; 昆兹,B。; 格鲁,R。; van der Goot,F.G.炭疽毒素的细胞内吞作用由网格蛋白、肌动蛋白和非常规适配器介导。 《公共科学图书馆·病理学》。 2010 , 6 ,e1000792。 [ 谷歌学者 ] [ 交叉参考 ] Abrami,L。; Kunz,B。; 范德古特(van der Goot),F.G.炭疽毒素触发src样激酶的激活,以调节其自身的摄取。 程序。 国家。 阿卡德。 科学。 美国 2010 , 107 , 1420–1424. [ 谷歌学者 ] [ 交叉参考 ] Abrami,L。; 刘,S。; Cosson,P。; 莱普拉,S.H。; van der Goot,F.G.炭疽毒素通过脂筏介导的氯氰菊酯依赖过程触发其受体的内吞作用。 《细胞生物学杂志》。 2003 , 160 , 321–328. [ 谷歌学者 ] [ 交叉参考 ] Deinhardt,K。; O·伯宁豪森。; Willison,H.J。; 霍普金斯,C.R。; Schiavo,G.破伤风毒素通过脂质微结构域内启动的连续网格蛋白依赖性机制内化,不依赖于epsin1。 《细胞生物学杂志》。 2006 , 174 , 459–471. [ 谷歌学者 ] [ 交叉参考 ] Van Dam,E.M。; Stoorvogel,W.通过内体衍生的网格蛋白涂层囊泡回收动力蛋白依赖性转铁蛋白受体。 分子生物学。 单元格 2002 , 13 , 169–182. [ 谷歌学者 ] [ 交叉参考 ] Van Dam,E.M。; Ten Broeke,T。; Jansen,K。; Spijkers,P。; Stoorvogel,W.内吞转铁蛋白受体通过不同的动力蛋白和磷脂酰肌醇3-激酶依赖性途径进行再循环。 生物学杂志。 化学。 2002 , 277 , 48876–48883. [ 谷歌学者 ]