1.简介
近年来,人们对含有抗氧化特性的植物和食品的兴趣增加了。蔬菜和水果中具有这些能力的化合物有:维生素C和E、类胡萝卜素和类黄酮。花青素是植物中最重要的类黄酮类物质,是一种含有黄烷阳离子(AH)的色素+)充当酸的结构。这种结构与其抗氧化活性直接相关。花色苷的大部分功能特性和感官质量可以用其化学反应性来解释。花青素的结构和性质取决于不同的因素,如pH值、温度和溶剂,这些因素应加以控制,以便对这些化合物进行抗氧化活性研究[1,2,三,4,5,6,7,8,9]. 自由基、活性氧(ROS)和/或活性氮(RNS)是人体及其器官正常运行所必需的。这些自由基通过我们体内的氧化还原内稳态而处于平衡状态。然而,身体可能偶尔会受到失衡状态导致的氧化应激的影响。这种压力对包括冠心病、癌症和衰老在内的慢性退化性疾病的发展至关重要[2]. 花青素被描述为通过清除自由基和减少氧化应激来防止或抑制氧化的化合物。通常,花青素充当氢原子供体或单电子传递。基于这两种机制提出了不同的分析方法来测定花色苷的抗氧化活性。这些化合物的抗氧化活性取决于其总浓度、结构和环境。本文综述了不同食物中最常见的花青素浓度的文献汇编,以综述花青素的不同来源。 饮食中摄入富含花青素的食物的有益特性(眼睛健康、心血管疾病、抗肥胖、抗糖尿病、抗菌、抗癌或神经保护作用)已在实验模型的研究中得到深入记录。这些健康益处与我们有机体吸收的这些化合物的明显小部分(<1–2%)形成对比[三,4,5]. 在消化过程中,花色苷的pH值发生剧烈变化,加上酶和细菌的作用,花色素会水解并转化为代谢物、共轭产物或简单的酚类化合物[7,10,11,12]. 问题是:花青素如何对健康产生如此大的影响?花青素是其有益作用的唯一原因吗?最近的科学发展突出了母体化合物、代谢物(阶段I和II)、共轭产物和微生物生成代谢物之间的潜在协同效应,以解释这些生物事件[4,11,12,13]. 由于花青素特殊的物理化学特性,其生物利用度很难评估。最初的研究是通过分析血液和尿液来确定摄入富含花青素的食物后的花青素浓度水平[14,15,16]. 然而,获得的低吸收率导致了体外检测(主要使用细胞培养系统),以便于了解其生化和化学变化以及消化步骤的影响。最近的研究强调了微生物群在花色苷转化中的关键作用,这在体外试验中未被考虑,但在体内和体外研究中仍缺乏考虑[14]. 这篇综述旨在强调花青素的抗氧化活性及其摄入后的生物利用度。第一部分阐述了用于体外测定花青素抗氧化活性的最常见抗氧化生物测定方法,以及每种生物测定方法的优缺点。然后,讨论了化学结构和环境对花青素抗氧化能力的影响,并介绍了这些化合物在食品中的不同来源和浓度范围。手稿的第二部分揭示了花青素的生物利用度和代谢,并对花青素对不同疾病的主要治疗作用进行了总结。
2.花色苷的抗氧化生物测定
已经提出了许多抗氧化测试来衡量花青素阻止自然发生的氧化过程的能力。根据花色苷的来源及其性质,在大多数情况下,在进行抗氧化生物测定之前需要进行提取步骤。提取过程是测定抗氧化活性生物测定的关键步骤,由于提取后花青素的稳定性较低,且花青素倾向于与样品基质结合,因此存在挑战。文献中提出了该程序的多种替代方案[17]. 结果表明,温度、pH、溶剂体系、溶剂固比和萃取次数是影响萃取效率的重要因素,应针对每个样品进行优化[18,19]. 一旦花青素处于液体溶液中,就可以通过不同的生物测试来测定其抗氧化活性。总的来说,可以用两种不同的机制来解释花色苷的抗氧化活性:氢原子供体(HAT)和单电子转移(SET)。在HAT机制中,自由基R•从抗氧化剂中去除一个氢原子(AH+)将自由基转化为更稳定的产物。在SET机制中,抗氧化剂(AH+)给自由基提供一个电子,将氧化的中间产物还原成稳定的形式[20]. 然而,区分HAT和SET反应的难度很高。在大多数情况下,这两个反应同时发生,反应机理由抗氧化剂结构、溶解度、分配系数和溶剂极性决定[21]. 文献中常用不同的抗氧化生物测定来测定花青素的抗氧化活性。花青素最常用的抗氧化生物检测方法总结如下表1。 这些方法中的每一种都提供了特定条件下花色苷抗氧化活性的信息。为了获得更多关于反应机理的信息,对每一种生物测定所遵循的程序进行了更全面的描述,并介绍了它们的主要优缺点。
2.1. DPPH(二苯基-1-苦酰肼)测定
DPPH分析是一种分光光度法,可用于固体和液体样品,但不适用于任何特定的抗氧化剂。因此,它可以用于测定样品的总抗氧化能力。该分析基于自由基(DPPH)的能力•)与氢供体(AH)反应+). 自由基在515nm处的紫外可见光谱区域呈现强烈的衰减,当自由基减少时,515nm的吸光度降低。
花色苷对DPPH的抗氧化活性已由几位作者进行了评估[22,23,24,25,26]. 该方法快速简便,结果可靠。目前,在没有分光光度计的情况下,提出了一种比色法替代方法来扩展该分析的应用[27]. 此外,由于自由基是稳定的,所以每次进行分析时都不必生成自由基。主要缺点可能是分析的复杂性,因为当样品中存在的其他化合物也在515 nm处吸收,并且许多抗氧化剂对DPPH不起作用时,分析变得更加复杂。 2.2。ORAC分析(氧自由基吸收能力)
ORAC分析是一种结合样品(AH)的荧光方法+)使用荧光化合物藻红蛋白(β-PE)和自由基发生器2,2′-偶氮双(2-氨基丙烷)盐酸盐(AAPH)。这种分析是基于荧光化合物被生成的自由基氧化时的损失。当抗氧化剂(AH+)在反应中,荧光化合物免受氧化降解。因此,荧光信号仍然存在。分别在λem=565 nm和λex=540 nm的条件下监测1 h内的荧光信号。几位作者通过ORAC分析评估了花青素的抗氧化活性[26,28,29,30]. 这被认为是一种测定亲水性和疏水性样品抗氧化能力的好方法,因为它很容易适应不同样品改变自由基发生器的情况。一个缺点是荧光化合物的非特异性,即使不添加自由基发生器,也会与失去荧光的样品发生反应。 2.3. TRAP分析(总过氧化物自由基捕获抗氧化剂参数)
TRAP分析是一种化学发光方法,由以下组分组成:(i)不耐热偶氮自由基引发剂(例如,2,2′-偶氮双(2-氨基丙烷)盐酸盐(ABAP)),其产生自由基(R•)与O快速反应2生成过氧自由基(ROO•); (ii)具有化学发光特性的可氧化化合物,以监测反应进程(例如鲁米诺);和(iii)具有抗氧化性能的样品(AH+).
几位作者通过TRAP测定法评估了花青素的抗氧化活性[31,32,33,34]. 该试验对所有已知的断链抗氧化剂都很敏感。然而,需要强调的一个重要缺点是,由于可以使用的不同端点的数量,很难比较实验室之间的结果。这是一种相对复杂的方法,耗时,并且需要高度的经验。此外,如ORAC分析,可氧化化合物是非特异性的,可能会发生其他非自由基连锁反应。 2.4. FCT(硫氰酸铁)测定
FTC分析是一种分光光度法,通过与过氧化物反应将氯化亚铁氧化为铁离子。当样品与乙醇、水、磷酸盐缓冲溶液(pH=7)和亚油酸混合时,在反应过程中会形成过氧化物。然后,将盐酸中的氯化亚铁加入反应中,形成的铁离子与硫氰酸铵结合,生成红色的硫氰酸铁。在500 nm处测量样品的吸光度,直到达到最大值。
该分析用于测量氧化初始阶段产生的过氧化物量。就花青素而言,该方法已用于测定抗氧化活性,以避免多不饱和脂肪酸的过氧化[35]. 该分析方法简单,重复性好。然而,如果样品中的化合物吸收约500 nm,则结果不可靠。这种缺点不仅在这种方法中常见,在其他分光光度分析中也常见。 2.5. 铁还原抗氧化能力(FRAP)测定
FRAP分析是一种比色法,可用于测定花青素的总抗氧化活性。它基于2,4,6-三吡啶-s-三嗪(TPTZ)与六水合三氯化铁(FeCl)的络合物的还原三·6小时2O) 在酸性条件下。还原完成后,溶液会变成略带褐色,形成蓝色的亚铁络合物。在593 nm处测量空白溶液的吸光度。
该方法已广泛应用于测定不同基质中花色苷的抗氧化活性:接骨木[36];玫瑰茄提取物[37];覆盆子、黑莓、红醋栗、醋栗和Cornelian樱桃[38];或者在胡萝卜、卷心菜、花椰菜、土豆、洋葱、芦笋和茄子中[39]. 然而,在大多数情况下,这种方法的结果与其他抗氧化剂检测的结果相比[40]. 用这种方法可以获得快速且可重复的结果。然而,应考虑与FRAP分析相关的一些限制。一方面,样品必须是水性的,pH值至关重要。另一方面,一些没有抗氧化性能的化合物可以减少铁3+至Fe2+导致对样品抗氧化活性的过高估计。 2.6. CUPRAC(铜离子还原抗氧化能力)测定
CUPRAC分析是一种类似于FRAP的分光光度法。在这种方法中,花色苷与CUPRAC试剂(新铜试剂)反应,生成Cu(I)-新铜试剂,它是一种在450 nm处吸收的生色团。该方法已成功应用于各种食品提取物[7]. 该方法的主要优点与CUPRAC试剂的积极特性有关:可用性和易得性、快速性、稳定性、低成本、对硫醇型抗氧化剂的敏感性(与FRAP不同),以及对亲水性和亲脂性抗氧化剂的反应性。然而,检测是基于在450 nm处的吸收,这是非特异性的,样品中存在的其他化合物可能会干扰结果。
2.7. ABTS(2,2′-偶氮-双(3-乙基苯并噻唑啉-6-磺酸)二铵盐)测定
最新的ABTS测定方法是基于脱色技术。它包括通过ABTS与过硫酸钾反应生成稳定的自由基,即蓝色/绿色ABTS发色团。这种稳定的自由基在415nm处具有最大吸光度。当自由基与抗氧化剂反应时,这种化合物的吸光度会下降。这种方法可以测定物质混合物的抗氧化活性,有助于区分添加剂和协同效应。抗氧化活性是根据类似条件下Trolox标准的反应性进行计算的。
该试验经常与DPPH试验相结合,以测定花青素的抗氧化活性[41,42,43]. 该方法简单,不需要大量样品和分析时间。然而,为了获得准确的结果,需要使用标准溶液。 文献中也将该分析描述为TEAC[6-羟基-2,5,7,8-四甲基铬-2-羧酸(Trolox)]等效抗氧化能力。TEAC和ABTS分析均使用相同的自由基。然而,已经提出了不同的试剂来生成绿-蓝ABTS•+,导致不同的TEAC分析。该自由基是由2,2′-叠氮双(3-乙基苯并噻唑啉-6-磺酸;ABTS)氧化产生的。氧化可以通过不同的方式实现:(i)在TEAC分析i中,高铁肌红蛋白与H反应2哦2产生铁酰肌红蛋白自由基,然后与ABTS反应。(ii)在TEAC分析ii中,ABTS•+是由与二氧化锰反应形成的。(iii)在TEAC分析iii中,使用辣根过氧化物酶进行酶反应。其他建议包括应用电化学氧化或使用2,2′-偶氮二-2-氨基丙烷、二氯化氢(AAPH)或过硫酸钾(K2S公司2哦8)作为氧化剂。
当选择合适的溶剂时,这些替代品是可互换的。然而,可以发现分析条件的差异,例如用于检测的反应时间或波长,有时为了避免干扰,734 nm比415 nm更可取。
这些测定已被用于测定花青素的抗氧化能力,因此在葡萄酒中已经报道了一些结果[44];在玉米中[45];石榴汁[46]或者在蓝莓中[47]. TEAC分析I的优点是简单、重复性和灵活性,可用于测定亲水性和亲脂性食品中的抗氧化能力。然而,在自由基生成之前,样本的成瘾可能会导致对抗氧化能力的过高估计。该试验的其他可能缺点可能是ABTS不是天然存在的,并且任何氧化还原电位低于ABTS的化合物•+可能与基团发生反应。
用于测定花青素抗氧化活性的其他替代品包括使用酶[48]或色谱技术的使用。后者不仅可以提取花青素来测定其抗氧化活性[49],还可测定总花青素浓度(TAC),以及样品中每种花青素存在的识别和量化[50,51]. 本文中暴露的体外测定提供了有关花青素抗氧化活性的信息,花青素的抗氧化活性由其中和氧化过程引发剂(吸收光子、中和ROS或螯合金属离子)的能力决定,从而停止自氧化过程的引发步骤。因此,这些分析是基于花色苷与以下物质的反应:(i)一些有色持久性自由基,例如DPPH测试和TEAC测试中使用的自由基;或(ii)其他氧化剂,如铁3+FRAP测试中使用的离子或铜2+CUPRAC测试中使用的离子。这种中和能力是在特定的温度、光照或两者结合的条件下测量的。其他测量花青素抗氧化能力的分析是基于竞争性探针反应,其中自由基与花青素或探针的竞争性反应在ORAC分析中通过荧光技术进行监测。所有这些方法都提供了实际抗氧化活性的信息。它们捕获自由基,应被视为断链抗氧化剂。只有使用基于抑制自氧化的方法才能正确测量该特性,而其他方法(如DPPH、ABTS)只能提供粗略估计。然而,已经进行了不同的具体研究,以确定其预防性抗氧化能力[52]. 可以在文献中找到其他特定的分析方法,以收集有关抑制脂质底物氧化的信息,同时研究氧化的不同因素和机制[53]. 如上所述,本文中的分析遵循不同的反应机制来测量抗氧化能力。因此,应仔细考虑他们提供的信息及其结果的解释。例如,FRAP和CUPRAC分析与ORAC分析提供的结果就是这样,ORAC分析包含自由基捕获反应,并直接测量中和引发剂的能力。
尽管上述方法是最常用于测定花青素抗氧化能力的方法,但目前有人主张开发不同的替代品,以提供关于生物系统中防止自氧化能力的直接信息。人们提出了多种替代方法来测量对自氧化的抑制。这就是Matera等人所揭露的情况[54]其中,抗氧化活性是通过研究水胶束中被抑制的自氧化来测量的。为此,将亚油酸和Triton-X100在受控pH下与引发剂和提取的花色苷混合。通过使用克拉克型电极监测耗氧量来跟踪反应。Tisuda等人提出了另一种测量花色苷抑制自氧化能力的替代方法[55]其中,通过使用亚油酸在不同体系中的自氧化来测定抗氧化活性;脂质体、兔红细胞膜和大鼠肝微粒体。这类研究提供了有关抗氧化剂检测结果与其在防止自身氧化中的生物活性之间关系的直接信息。目前,欧洲食品安全局(EFSA)发布了一份关于抗氧化剂、氧化损伤和心血管健康相关健康声明的科学要求指南。在本指南中,欧洲健康声明法规建议确定具有抗氧化活性的食品补充剂的生物效应。根据本文件,食品的抗氧化特性基于在模型系统中进行的体外研究清除自由基,目前还没有确定这种能力对人体有有益的生理作用。因此,使用TRAP、TEAC、FRAP和ORAC等方法对血浆进行的体内人体研究并未证明其对人体具有有益的生理作用。因此,保护细胞免受过早老化以促进健康老化与食品抗氧化性能之间的关系并没有得到充分定义,因为它们是根据非特定标准建立的。为了提供有关在特定标准下防止自动氧化的信息,EFSA指南建议对特定生物分子进行一些分析。例如,它建议:(i)监测F2-异前列烷,以确定保护脂质免受氧化损伤的能力;(ii)分析血液、组织和尿液中的8-羟基-2-脱氧鸟苷,以评估DNA的氧化损伤;以及(iii)通过ELISA监测蛋白质羰基,可用于确定蛋白质免受氧化损伤的保护[56]. 本手稿中详细解释的所有测定提供的结果通常将抗氧化活性表示为每千克新鲜重量mmol Trolox当量。然而,这些结果并没有确切报道食品的抗氧化活性。为此,有关饮食中每种食物的大小比例的信息对于评估其抗氧化活性也很重要[57]. 由于这类结果与花青素抗氧化活性有关,美国农业部标准参考营养数据库公布了一个数据库,其中确定了不同水果和蔬菜花青素的总浓度,以及每种水果或蔬菜应摄入的服务量[58]. 3.花色苷的分类和天然来源
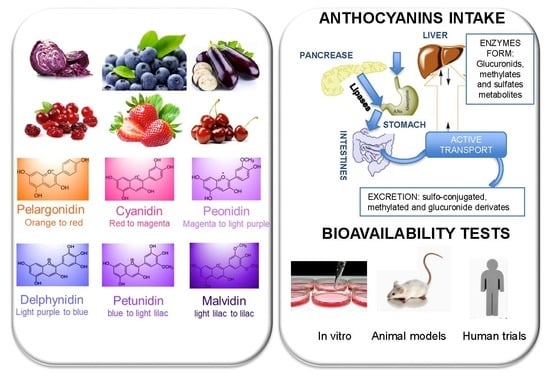
花青素的健康和治疗效果与它们的化学和生化反应有关,这部分是由于它们的抗氧化活性[59,60]. 然而,花青素的抗氧化活性并不一定转化为生物活性,因为对身体的任何作用都取决于生物利用度和细胞分子靶点[61]. 此外,并非所有蓝色、红色和紫色的水果、蔬菜和花卉都具有相同的花青素成分和浓度,因此具有相同的抗氧化活性。花青素含量最高的水果有浆果、葡萄干、葡萄和一些热带水果。在可食用蔬菜中,叶菜类蔬菜、谷物、根和块茎的花青素浓度也最高[62]. 此外,可以在植物的不同部位,如茎、叶和贮藏器官中检测到花青素的存在。 在自然界中,这些色素通常以花青素的形式存在,花青素以糖苷的形式存在;花青素也称为苷元(一种或多种糖与苷元结合)。花色苷的基本结构如所示图1如今,自然界中鉴定出的花色苷数量超过600个[6]. 在这些花色苷中,糖苷形式的飞燕草苷、花青素、牵牛花苷、芍药苷、马来黄素和天竺葵苷最为丰富[54,55,62,63,64,65].表2显示了在不同水果、蔬菜和食用花卉中发现的最常见的花青素,以及分配给每个花青素的数字代码,以在手稿中引用它们。 在补充信息,表S1–S6显示了不同水果、蔬菜和花卉中最常见的花青素,根据其化学结构进行分类:飞燕草素及其衍生物(表S1)、氰化物及其衍生物(表S2),牵牛花苷(表S3)、芍药苷(表S4)、丙氨酸(表S5)天竺葵素及其衍生物(表S6). 每张表还说明了花青素的天然来源、所用提取物的类型、用于识别和量化花青素所用的色谱法以及用于确定抗氧化能力的抗氧化分析。 主要结论来自表S1飞燕草苷3-葡萄糖苷(代码4)是水果和食用花卉中最常见和最丰富的飞燕草素。然而,这类花青素在蔬菜中并不常见,其中最常见的是翠雀碱。结果显示在表S2指出水果和可食用花朵中最常见的花青素是花青素3-葡萄糖苷(代码11),也是水果中丰富的花青素3-半乳糖苷(代码10)。在蔬菜中,花青素(代码8)是最常见和最丰富的,但一些谷物中也富含花青素3-葡萄糖苷(代码11)。表S3结果表明,在几乎所有研究的水果中都能鉴定出以下花色苷:矮牵牛苷3-阿拉伯糖苷、矮牵牛素3-半乳糖苷和矮牵牛蛋白3-葡萄糖苷(代码30、31、32),其中矮牵牛肽3-葡萄糖甙(代码32)是花中最丰富的也是唯一鉴定出的矮牵牛甙。牵牛花苷(代码29)是在所研究的蔬菜中唯一鉴定出的这类花青素。表S4发现在花中没有发现芍药苷。在水果中,芍药苷3-葡萄糖苷(代码37)含量最高,但大多数水果中也鉴定出芍药苷-3-半乳糖苷(代码36)。在蔬菜中,这类花青素中最常见的是芍药苷(代码35)。来自malvidin组(表S5)可以得出结论,水果中最丰富的是malvidin3-半乳糖苷(代码51),其次是malvidian-glucoside(代码52)。最后一个阶段的花也是最丰富的,其次是malvidin 3,5-二葡萄糖苷(代码53)。在蔬菜中,malvidin是该组中唯一量化的花青素。表S6结果表明,果实中天竺葵素3-葡萄糖苷(代码55)含量最高,其次是天竺葵苷3-芸香苷(代码56)。天竺葵素3,5-二葡萄糖苷(代码57)在花中含量最高,天竺葵苷(代码54)是研究蔬菜中唯一鉴定出的这类花色苷。 这些颜料对氧化过程的保护作用取决于它们的结构。并不是所有它们都具有相同的清除活性氧或氮物种的活性。花青素的抗氧化能力取决于环的取向,因为它将决定捐献质子的意愿以及传递和电子的能力。吡喃环周围的游离羟基数量及其位置也对抗氧化活性起着关键作用[2]. 在主体结构中存在其他类型的自由基对抗氧化活性也有重要作用。因此,花色素苷查尔酮和与酮基团共轭的双键醌基是清除自由基的有效抗氧化剂。此外,花色苷的糖基化B环结构有助于提高抗氧化活性,其中邻羟基化和甲氧基化显著提高抗氧化活性。此外,与文献报道的花青素相比,花青素具有更高的抗氧化活性。原因可能是由于花青素的结构使其稳定性低于花青素,从而使花青素具有高度活性[66]. 花青素与一种或多种酚酸的酰化反应可显著提高抗氧化活性[54,67],但糖基化导致活性降低[66,68]. 不同的花青素清除不同自由基的功效不同。天竺葵苷-3-葡萄糖苷、花青素-3-葡萄苷和飞燕草苷-3-葡糖苷及其标准苷元在脂质体系统中具有较强的抗氧化活性,并通过UVB辐射减少丙二醛的形成[55,69]. 此外,结果还表明,该药物对脂质过氧化和氧的抑制作用最强2•飞燕草素和飞燕草苷-3-葡萄糖苷的清除活性,其次是花青素和天竺葵素[2]. 相反,天竺葵素对羟自由基清除活性的抑制作用最强[66]. 此外,一项研究表明,与其他酚酸、花青素和花青素苷元相比,对铜(II)诱导的低密度脂蛋白(LDL)氧化花青素及花青素-3-葡萄糖苷的抑制作用最强,而飞燕草素具有中等功效[70]. 另一项研究分析了malvidin-3-葡萄糖苷的抗氧化活性,结果表明,与儿茶素、malvidin和白藜芦醇相比,malvidin3-葡萄糖甙的醌基和假碱显著抑制亚油酸的过氧化作用[71]. 然而,分配给这些花色苷的氧化活性取决于活性物种的类型,因此也取决于为测定抗氧化活性而进行的抗氧化分析的类型。因此,FRAP和TEAC分析报告了分别在牵牛花苷和马来酸苷的5或3和5位通过转氨酶显著降低抗氧化活性[72]. 另一个与花色苷的反应性和抗氧化活性有关的因素是pH值。在文献中,不同的研究已经证明了pH值对不同来源花色苷抗氧化能力的影响;罗塞尔[26],葡萄酒[71,72],黑米与环直链淀粉复合[73,74]、棕榈汁[75]、和红叶槿[76]. pH值是一个重要因素,应加以控制,以确定花色苷的反应性。花青素结构的酸性如所示图2这种酸性性质是由于主结构环中的双键与C4′、C5和C7处的羟基发生共轭。C7处的羟基是最强的酸。脱质子化可以在酸性pH~4下进行,生成中性醌碱,该碱通过与C5处的羟基互变异构而稳定。C4处的羟基也容易在较高的pH~7下脱质子,从而产生阴离子碱。如果pH值仍然上升到基本pH值,高于8,则在C5中发生脱质子反应,生成可导致查尔酮阴离子的二离子碱[1]. 因此,溶液的pH值控制着质子化和去质子化水合花青素和异构体花青素的比例,从而影响其反应性。因此,在花青素的提取过程中以及抗氧化生物测定过程中都应该控制pH,因为它们的结果是pH依赖性的[2,26]. 一旦对不同食物中最常见的花青素进行了鉴定,并讨论了结构和环境对花青素抗氧化能力的影响,下一步就是确定这些花青素的生物利用度及其影响,以增进人类健康。
4.花青素的生物利用度
花青素的每日摄入量可以通过食物数据库进行估算,由于评估方法的差异,以及受调查人群的营养、文化和社会差异的影响,花青素摄入量从每人几毫克到数百毫克不等[8]. 口服后花青素的模式是独特的,与其他类黄酮不同[77]. 花青素的生物利用度明显较低,只有1–2%的摄入花青素在生物体内保持其亲本C6–C3–C6结构。食物消化是一个依赖pH的过程,因此,除了小肠中的几种酶水解外,花青素还会发生转化[12,78]. 摄入的花青素的一部分到达大肠,在那里代谢为低分子量的分解代谢产物,可在2-4小时(最多8小时)内排泄到粪便中或再次被吸收。通过胃或肠细胞屏障的活性转运蛋白在肝脏、肾脏、大脑或除胃外的其他器官和组织内的转运和吸收中起着重要作用[13,79]. 在最近一篇关于动物组织生物利用度的综述中,Sandoval-Ramírez等人[80]得出的结论是TAC吸收量为2.17×105小鼠肾脏pmol/g,1.73×105肝脏pmol/g,3.6×10三pmol/g心脏,1.16×105肺pmol/g;和6.08×10三猪脑pmol/g。在肠壁和肝脏中,花青素及其分解代谢产物经历第2阶段的酶代谢,也转化为葡萄糖醛酸化、硫酸化和甲基化形式[10,12,13,14,78,81,82,83]. 体内许多部位都存在微生物分解代谢产物,其浓度高于天然形式,这表明花青素的部分生物活性与其结肠分解代谢产物的协同作用有关[13,84]. 花青素代谢产物和转化产物已由几位作者进行了表征和量化[85,86,87,88,89]. Ferrars等人[88]在绝经后妇女服用接骨木12周后,发现了多种花青素酚类代谢产物,包括11种新的代谢产物,其浓度高于其天然花青素形式。影响花青素及其代谢物在生物体内命运的关键因素有很多:跨膜能力、pH值、消化酶、微生物群、胆汁酸或食物基质(14C) 或稳定同位素标记(13C) 示踪研究提供了有关花色苷代谢为酚酸衍生物的程度的有用信息。在这个意义上,Czank等人[90]研究了8名男性参与者摄入13C-氰化物-3-哦-葡萄糖苷(500 mg)。相对平均生物利用度为12.38%(5.37%通过尿液排泄,6.91%通过呼吸排泄)。作者发现13C-标记代谢物比其各自的天然化合物13C-氰胺-3-葡萄糖苷。检测到多达49种代谢物,其中包括:氰胺-3-葡萄糖苷和氰胺的II相结合物(氰胺-葡萄糖甙、甲基氰胺-葡萄糖甙和甲基氰胺-3-葡萄糖苷-葡萄糖甙);降解产物(原儿茶酸、间苯二甲醛和间苯三甲醛);原儿茶酸、苯乙酸、苯丙烯酸和马尿酸的II相共轭物。 花青素发挥其生物活性的机制尚未完全了解,因为尚不清楚其活性是否与天然形式、其衍生物或两者有关。区分它们不同的生物作用是一项非常具有挑战性的任务。对花青素代谢产物的抗氧化活性进行了一些比较研究[91]. 最近,Kim等人[92]提供了体外消化过程中氰苷类化合物化学变化的基本信息。氰化物-3-哦-半乳糖苷被降解为咖啡酰奎宁酸,在体外消化矢车菊素-3后未发现-哦-葡萄糖苷。在肠道部分,花青素代谢产物的生物活性(DPPH)降低。然而,在模拟结肠消化后,生物活性增加,可能是因为新形成的结肠代谢物。此外,与桑树提取物相比,草莓提取物中的花青素代谢产物表现出更高的DPPH自由基活性。在另一项研究中,共轭弹性蛋白酶-3的α-葡萄糖苷酶抑制活性和活性氧清除活性-哦-胃肠道消化后,糖苷样品可能增加[93]. 在人类消化过程的三个主要阶段观察到的物理化学反应的方案可以在图3[11]. 生物转化反应通过唾液淀粉酶在口腔中开始(pH值5.6–7.9)。一旦在pH值为1.5–3.5的胃中,花青素以多种离子形式存在,主要以红色黄杨阳离子和醌蓝种类存在。最后,在肠道步骤中(pH 6.7–7.4),花青素以无色甲醇的形式存在(吸收有限)并发生生物转化为低分子量分子,如酚酸或邻苯二酚(没食子酸、香草酸、原儿茶酸、4-羟基苯甲酸和丁香酸已被确定为飞燕草素-3的主要降解产物-哦-糖苷、芍药苷-3-哦-葡萄糖苷、氰化物-3-哦-葡糖苷,天竺葵苷-3-哦-葡萄糖苷和malvidin-3-哦-分别为葡萄糖苷)[13]. 为了更好地了解花青素的生物利用度,已经提出了不同的体内和体外模型来模拟消化[11]. Gowd等人[94]评估黑莓花青素提取物的酚类成分,然后在不同时间间隔(0-48h)进行人体肠道微生物发酵。作者揭示了肠道代谢物的形成增强了高糖加棕榈酸诱导的活性氧、线粒体膜崩塌和HepG2细胞中谷胱甘肽的耗竭。几项研究也报告说,在花青素结肠发酵后,有益细菌增加(双歧杆菌物种。,放线菌门,拟杆菌纲,乳酸杆菌/肠球菌物种。,阿克曼菌属) [95,96,97,98,99,100]. 肠道微生物群具有β-葡萄糖苷酶活性,可以从苷元中释放葡萄糖,并提供能量支持细菌生长。Zhou等人最近进行的一项研究[95]提示食用蓝莓及其提取物可以发挥益生元活性,并对人体肠道微生物群的组成和丰度产生调节作用。花青素可以通过调节肠道微生物来增强人体健康,而肠道微生物通常与各种疾病有关[95,101]. 然而,值得注意的是,花青素衍生物还可以减少一些有害细菌,例如溶组织梭菌结肠发酵后[101,102]. 花青素结肠代谢代谢产物和结肠微生物群变化的总结如所示图4[11]. 无论如何,对于所涉及的机制以及哪些因素对生物利用度有关键影响,仍有许多疑问[7]. 花青素对一个人有效,但对另一个人的效果可能不同[12]所得结果仍存在较大的变异性。根据文献和最近的评论[11,14]这种可变性是由于在三个层面上引入的缺乏同质性:(i)食品基质和食品加工;(ii)酶水平(受遗传因素和饮食、年龄和性别的影响);以及(iii)微生物群功能。考虑到诱导间或诱导内变异性的报告数据非常稀少,生物利用度方法也没有标准化,因此很难得出确切的结论。一方面,体外方法(基于细胞的分析)没有考虑到人体中单个微生物群的作用;而另一方面,体内试验(人体试验和动物研究)每个受试者都有自己的微生物群[14]. 值得注意的是,由于花青素的高度不稳定性和易降解性,将其纳入食品和医疗产品中是一项具有挑战性的任务。从这个意义上说,在体外模拟胃肠消化和结肠发酵中,使用纳米/微胶囊与天然聚合物是提高敏感物质稳定性的最佳策略之一[103,104]. 根据最近关于这个主题的评论[104]已测试了不同的技术来封装花青素,包括喷雾干燥>冷冻干燥>凝胶化>基于脂质的颗粒>电动水动力过程。第一种方法是最经济、最简单、最实用的方法(80-90%)[105,106]. 由于花青素的亲水性,其他技术的使用仍然缺乏探索,因此是未来研究的一个有希望的领域[104,107,108,109]. β-环糊精包裹的黑莓花色苷[101,110]或阿拉伯树胶[111]有助于延缓体外模拟胃肠消化过程中花青素的释放。花色苷的稳定性也会受到壁材类型的影响。最近,Wu等人[103]评价了四种不同壁材在体外模拟消化和结肠发酵过程中的作用。包封技术显著提高了结肠的通畅性,延缓了花青素的释放,尤其是大豆蛋白。通过肠道微生物群的作用,在结肠发酵过程中产生的花青素降解产物,如丁香酸,表明其对宿主健康有益。 5.花青素的治疗作用
眼睛健康:自1966年第一份关于花青素对人类视力的积极影响的报告以来,富含花青素的提取物已被世界各地广泛用作眼部健康的补充物[161,162]. 口服黑醋栗花青素可能是开角型青光眼患者的一种有希望的补充,对抗青光眼药物也有效,而富含花青素的越桔提取物在视网膜炎症期间对视力有保护作用[115]. 已经证实,氰化物有助于大鼠视紫红质的再生和平滑肌松弛[116]. 结果还表明,越桔提取物能够抑制不规则吡啶A2E的光氧化作用,这是一种随着年龄增长而积聚在视网膜上皮细胞中的自荧光色素,可对细胞造成光损伤。在一项比较研究中,与安慰剂组相比,近视受试者的夜间视觉功能显著改善,对比敏感度水平提高[114]. 花青素还可抑制短暂性近视,减轻眼睛疲劳或增加青光眼视网膜血流量[118,121,161,163]. 心血管疾病:花青素在预防心肌梗死和与死亡率相关的心血管疾病方面的作用尤为重要。花青素提取物已被用于抑制血栓初期的血小板聚集;在治疗毛细血管脆性引起的微循环不良问题方面;并防止LDL氧化[122,164,165,166]. 在一项针对血脂异常患者(40-65岁)的安慰剂对照试验中,摄入浆果衍生花青素通过抑制胆固醇酯转移蛋白改善了脂蛋白谱[123]. 作者观察到,与安慰剂组相比,花青素组的高密度脂蛋白(HDL)胆固醇水平和细胞胆固醇流出到血清中的量增加,LDL胆固醇水平降低。Álvarez-Suárez等人也报告了类似的结果[127]在一项体内研究中,健康志愿者补充了草莓(500克)。每日摄入可改善血脂状况,降低总胆固醇、LDL胆固醇和甘油三酯水平,而HDL胆固醇保持不变。这增加了受试者的抗溶血防御和血小板功能。在另一项针对24年以上男性的前瞻性队列研究中,水果花青素摄入量增加与非致命性心肌梗死(14%)和缺血性中风风险降低相关[124]. 对45项随机对照试验的荟萃分析表明,食用浆果和纯化花青素(2.2−1230 mg花青素/天)显著增加HDL胆固醇,降低LDL胆固醇、甘油三酯、收缩压和舒张压,以及炎症标记物CRP和TNFα[167]. 分析还表明,一些人更容易受到花青素摄入的保护作用:(i)超重;(ii)超过50年;以及(iii)心血管疾病风险增加的人群。另一项对99项随机对照试验的荟萃分析表明,无论参与者的健康状况如何,食用富含花青素的产品都能显著降低收缩压和舒张压[168]. 在体外实验中,花青素也显示出对猪胰腺弹性蛋白酶的抑制作用[169],一种酶,通过攻击纤维和胶原蛋白,在动脉硬化、肺气肿或类风湿性关节炎等疾病中发挥重要作用。此外,富含花青素的提取物加速了愈合过程,显示出对大鼠胃十二指肠溃疡的预防和治疗作用[7]. 它们对粘多糖生物合成的影响可以证明可以提高胃粘膜层的功效,增加结缔组织和毛细血管的基础物质[170]. 抗肥胖和抗糖尿病作用:花青素通过多种机制显示出抗肥胖作用,如抑制脂质吸收、调节脂质代谢、增加能量消耗、抑制食物摄入和调节肠道微生物群,这表明花青素是抗肥胖治疗的潜在候选药物[171]. Kwon等人[128]观察到,添加黑大豆花青素的饮食降低了大鼠的体重增加,与不添加黑大豆的高脂饮食相比,添加高脂肪饮食加黑大豆花青素的大鼠体重增加显著降低。花青素还改善了脂质状况,抑制了高脂肪饮食诱导的肝脏重量增加,并降低了附睾和肾周脂肪垫的重量。 此外,2型糖尿病与肥胖密切相关[66]. 花青素可以通过抑制肠道葡萄糖吸收、诱导胰腺胰岛素分泌、上调4型葡萄糖转运蛋白和抑制肝脏糖异生来缓解2型糖尿病的并发症[172]. Heyman等人在13周内用不同的浆果为小鼠补充高脂肪饮食后[173]观察到,与对照组相比,这些补充的小鼠体重增加较少,空腹胰岛素水平较低,并且对葡萄糖稳态有介导的积极影响。Jankowski等人[130]描述了用葡萄喂养的大鼠注射链脲佐菌素后,尿液和血清中的糖浓度显著降低。作者提出的花青素的机制是胶原蛋白、脂蛋白和糖蛋白的生物合成减少,弹性蛋白酶和腺苷脱氨酶的活性降低(糖尿病患者的活性都很高)。用樱桃治疗大鼠可显著降低血糖和尿微量白蛋白,并增加尿素中的肌酸酐分泌水平[174]. “红辣椒”(阿根廷的一种红色水果)的果肉、种子和果皮具有降血糖作用,可增加葡萄糖的吸收,降低葡萄糖的扩散速度,并促进葡萄糖在细胞膜上的转运[175]在体外模拟胃十二指肠消化。人类食用蓝莓和苹果/梨也与降低2型糖尿病风险相关[176]. 抗菌作用:花青素对多种微生物的抗菌活性也有很好的证明。破坏细胞壁、细胞膜和细胞间基质导致细胞损伤的可能机制[66,140,177]. 黑莓提取物具有抗菌活性,对嗜水气单胞菌和innocua李斯特菌[141]. 蔓越莓提取物对屎肠球菌对万古霉素耐药,铜绿假单胞菌,金黄色葡萄球菌、和大肠杆菌[142]. 不同类型的浆果提取物抑制革兰氏阴性菌,但不抑制革兰阳性菌[143]可能是因为革兰氏阴性菌对疏水性化合物起着预防屏障的作用,但对亲水性化合物没有作用[178]. 抗癌活性:许多作者已经描述了花青素抗癌活性的可能机制:抗突变活性;抑制DNA氧化损伤和致癌物激活;诱导II相酶解毒;细胞周期阻滞;抑制环氧合酶-2酶;以及诱导细胞凋亡和抗血管生成[179,180,181,182,183,184]. 在乳腺癌中,花青素通过直接作用于DNA片段化和促进MCF-7癌细胞死亡,抑制促进其进展和发育的关键调节剂[185,186]. 此外,研究表明花青素具有广泛的体外抗侵袭和体内抗转移活性。例如,飞燕草素可以作为一种潜在的抗转移剂,通过特异性抑制NF-κB依赖的MMP-9基因表达来抑制PMA诱导的癌细胞侵袭[187,188]. 在肺癌中,从桑树中分离出的氰-3-葡萄糖苷和氰-3-rutinoside可抑制A549细胞的迁移和侵袭,并降低MMP-2和uPA,增强TIMP-2和PAI。花青素还可以抑制致癌细胞的生长,从而引发结肠癌,诱导细胞凋亡,甚至可以在免疫反应中作为巨噬细胞的调节剂[180]. Forester等人[189]还报道了花青素代谢物降低结肠癌细胞活力、导致细胞周期阻滞和凋亡的积极作用。在口腔和宫颈癌中,通过牡丹苷3-葡萄糖苷和氰胺-3-葡萄糖苷的治疗,SCC-4细胞和HeLa细胞的侵袭性减弱[190]. 值得注意的是,花色苷的结构对其生物活性有很大的影响[191,192,193]. 从这个意义上讲,苷元、糖和酰化酸的类型,以及糖基化和酰化的位置和程度似乎是影响抗癌性能的主要因素[191]. Jing等人[192]利用人结肠癌HT29细胞株比较了富含花青素提取物的抗癌特性。作者报道了以下生长抑制率:紫玉米>荞麦和越橘>紫胡萝卜和葡萄>萝卜和接骨木。这些非酰化单糖基化花色苷比那些含有天竺葵苷、三糖苷和/或肉桂酸酰化的花色苷具有更强的抗癌性能。 神经退行性疾病:花青素也特别适合治疗神经退行性病,如阿尔茨海默氏症、帕金森氏症或肌萎缩性侧索硬化症。其主要机制包括抗氧化途径、钙稳态、炎症、蛋白质稳态以及促生存和促凋亡信号的平衡[194,195]. 在帕金森氏病的原代细胞模型中,从蓝莓、葡萄籽、芙蓉、黑醋栗和桑椹中提取的提取物可以抑制鱼藤酮引起的多巴胺能细胞死亡[154]. 此外,Strathearn等人[154]观察到,那些富含花青素和原花青素的提取物比富含其他多酚的提取物表现出更强的神经保护活性。 大鼠口服花青素(200mg/kg)能够调节胆碱能神经传递,恢复Na+,K+-ATP酶和钙2+-ATP酶活性,并防止东莨菪碱配伍引起的记忆障碍[156]. Rehman等人[157]显示了花青素的神经保护作用,该作用基于使用D-半乳糖诱导氧化应激和炎症反应的人工老化模型。其潜在作用机制包括:糖基化终产物受体表达降低、活性氧水平降低和脂质过氧化。Shih等人[155]观察到,喂食富含花青素的桑椹提取物的小鼠在回避反应测试中表现出淀粉样β蛋白显著减少,学习和记忆能力改善。与对照小鼠相比,喂食的小鼠在大脑和肝脏中也表现出更高的抗氧化酶活性和更少的脂质氧化。此外,经证明,富含花青素的桑椹提取物可以降低血清天冬氨酸转氨酶、丙氨酸转氨酶,甘油三酯和总胆固醇的水平,这些水平随着年龄的增长而升高。 此外,花青素的治疗效果可以通过封装来改善[158,159,160]. 例如,阿明等人(Amin et al[158]表明负载花青素的包封纳米颗粒被细胞迅速吸收,增强了其对淀粉样蛋白β毒性的神经保护作用,高于单独的花青素。在小鼠体内研究中也观察到类似的活性[158,160]. 6.结论
为了确定花青素的抗氧化活性以及花青素摄入对人体健康的影响,需要考虑许多因素。首先,需要从多参数角度评估抗氧化活性,因为总浓度、结构、样品性质、pH值和反应机理对其效果起着重要作用。应定期对每种机制进行一次以上的抗氧化剂分析。此外,强烈建议对花青素进行鉴定和量化,以确定样品抗氧化活性的准确值。其次,需要了解这些化合物摄入后的生物利用度。许多研究证明了富含花青素的提取物在预防疾病方面的益处。然而,值得注意的是,它们的功效取决于其生物利用度。在消化过程中,花青素被代谢成各种结合物,然后最终代谢成酚酸降解产物。累积的证据表明,所有可能的形式之间的协同效应可以解释它们所具有的健康促进特性。花青素吸收、代谢、分布和排泄的诱导间和诱导内变异也很明显。可能影响这种变异性的主要因素包括:食物基质和加工、酶水平和微生物群功能。应注意不同方面:(i)在评估生物利用度时,执行设计良好的标准化方法;(ii)考虑花青素代谢的诱导间或诱导内变异;(iii)评估花青素代谢产物在胃肠消化过程中的抗氧化活性;并且,由于花青素可能发挥不同的生物活性,(iv)绘制出更准确的表征曲线。









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}