该领域的狮子们在我们的期刊上发表了开创性的研究成果,来自Bridges,穆勒,麦克林托克和布伦纳,霍维茨,和哈特维尔。作者可以像卢里亚和德尔布鲁克那样在同一杂志上发表文章,和西沃尔·赖特,和罗纳德·费舍尔,克劳和木村(以及许多其他名人)。虽然遗传学提供了一个专业和科学的线索,可以追溯到我们领域的创始人,它也指向我们的未来.
约翰斯顿2014年,第10页
长时间遗传学读者一定会学习(参见Ganetzky和Hawley 2016该杂志的第一篇文章,第1卷,第1页,报道了一项基础研究,其结果继续在该领域占据中心位置:卡尔文·布里奇斯关于“非双连接作为遗传染色体理论的证据”的著名工作(桥梁1916).
100年来的世界纪录中还隐藏着什么宝藏遗传学,他们与现场正在进行的活动有什么联系?遗传学作者们可以高兴地发现,在早期的一卷中,对一个想法起源的探索不时会发现一种意想不到的联系。这篇文章讲述了一个这样的探索,它导致了第二H.S.Jennings在该杂志第一期第1卷第53页发表的关于“不同繁殖系统的数值结果”的文章(詹宁斯1916).
遗传外加剂
Amy Goldberg和我一直在研究基因混合,这是一个当前感兴趣的话题,经常出现在最近几期的遗传学(例如,洛赫穆勒等。2011;Verdu和Rosenberg 2011年;砾石2012;帕特森等。2012;杜兴等。2013;卢等。2013;梁和尼尔森2014;Lohse和Frantz 2014;桑德森等。2015;塞奇法尔等。2015). 在遗传混合种群中,由两个或多个长期分离的群体遗传混合而成的种群,就像经典的F种群一样2交叉,单个基因组由具有不同祖先的区域组成。种群遗传数据的激增,基因组片段起源种群推断的统计进展,对古代混合种群的兴趣,以及在自然实验形成的混合种群中绘制性状位点的前景,都有助于在遗传混合种群的研究中开展大量活动。
我们的重点是性别偏见混合,在混合人口的一个或多个祖先来源中,女性和男性的贡献水平存在差异。在人类中,性别偏见的混血儿很常见,通常在入侵人口中偏向男性,在本地群体中偏向女性。由于X染色体和常染色体具有不同的遗传模式,X染色体记录的创始雌性和雄性的特征与常染色体不同。已经建立了性别偏见混合物对常染色体标记影响的一般数学模型(戈德伯格等。2014),我们的目标是对X染色体执行相同的操作。
外加剂模型
根据我们之前的模型,我们考虑了两个来源人口(“1”和“2”),其中一个或两个来源对混合人口的贡献存在性别偏见。在一个具有离散世代的公式中,我们检查了特定来源群体中随机选择的女性和特定世代混合群体中随机选取的男性的预期X染色体混合——换句话说,就是模型预测的随机选择混合女性或男性的混合水平。特别令人感兴趣的是“混合隔离”,即在最初生成混合后,源群体的进一步贡献不会进入混合群体(Long 1991年).
离散时间模型通常是通过获得递归来研究的,递归可以根据前一时间步中的状态来描述系统的状态。Amy使用递归方法,发现作为时间的函数,可以使用迭代定义的序列计算预期的X染色体混合J型n个,初始条件后J型0=0和J型1=1遵循公式J型n个=J型n个−1+2个J型n个−2.如果后代的第一代n个=1在混合人口中以比例开始秒如果它的母亲和秒米其父亲来自源种群1,然后在混合隔离下,随机变量H(H)n个,f,来源1中随机女性X染色体祖先在世代混合群体中的比例n个≥1,以及H(H)n个,米,随机混合男性的相应外加剂,具有预期快速检查整数序列在线百科全书(斯隆2015)揭示了我们的序列J型n个,以0、1、1、3、5、11、21、43开头,被称为雅各布斯塔尔数。序列连续项的近似加倍产生了女性和男性贡献的近似2:1权重秒如果和秒米在方程式1和2中,这是一个合理的结果,因为在男女人数相等的种群中,女性体内的X染色体数量是男性相应数量的两倍。 然而,与此同时,序列问题的出现J型n个很特别。它的生成公式,J型n个=J型n个−1+2个J型n个−2,让人想起著名的斐波那契数列递归,F类n个=F类n个−1+F类n个−2我们知道,根据单倍体遗传(例如X染色体上的遗传),一个人的家谱祖先数量符合斐波那契序列(例如,盆地1963). 另一个X染色体问题中出现的相关序列不可能是巧合。但这些由简单线性递归构成的数字序列是从哪里进入群体遗传学的呢?
斐波那契数
搜索斐波那契数和群体遗传学很快就把我们引向了一个历史悠久的插曲(赖特1969第171页;普罗文1971第136页;克劳1987)包括计算一个常染色体二倍体基因座的杂合概率n个通过同胞交配的近交世代,从AA公司和aa公司父母。纯合性和杂合性概率的计算对于理解动植物育种方案对所产生品系特性的影响非常重要。不同的育种系统以其特有的杂交序列产生了多大程度的近交?
同胞关系问题很难解决,第一份解决方案的报告是珍珠(1913),是错误的。在对错误主张的一系列更正中詹宁斯(1914)使用离散时间递归表明n个第h代,包括as代n个=1从最初的杂交中出生的同胞,是F类n个/2n个−1他评论道:“这个公式是2的连续幂与斐波那契级数的连续项的组合,斐波那奇级数以一种奇怪的方式出现在各种自然现象中”(詹宁斯1914第693页)。
我们追踪了斐波那契数的来源。然而,它们是在常染色体计算中出现的。肯定有人看过X。
詹宁斯1914年的报告很简短,重点是纠正早先的错误。詹宁斯(1916) 遗传学接下来是文章。在这篇长达37页的论文中,詹宁斯不仅考虑了与珀尔交换的同胞交配和常染色体基因座的情况,还考虑了其他几个繁殖系统:例如随机交配、自交、与同一亲本交配的亲子后代在多代后代中繁殖,以及父母与自己的后代交配的后代。各种病例具有特征性的离散时间递归,描述了纯合子或杂合子在连续世代中的比例。
詹宁斯的文章令人印象深刻。它的问题是具有挑战性的,案例的数量远远超过了现代作家在一份手稿中的投入。与此同时,这很难理解。数字仅限于一些示意图,文章几乎全是数学,没有上下文。人们想了解更多关于亲子交配计划时的兴趣,在这种计划中,每个个体被用于两代,确切地说是两代。
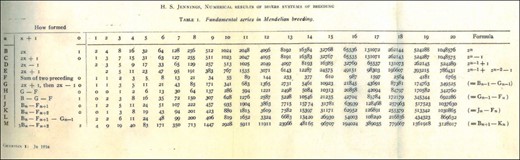
也许这篇文章中出现了X染色体?这篇文章有一个目录性别相关因素作为它的一部分。然而,一开始,这一部分是没有启发性的。但在获得每个繁殖系统的递归时,詹宁斯计算了前几个项,方便地将数值收集到一个综合表中。在表格中,斐波那契数列的正下方,用“G”简单注释的一行是雅各布斯数列——我们刚刚推导出的数字!
外加剂与育种系统之间的联系
我们回到了性别相关因素发现里面有神秘的G(图1)Jacobsthal序列是否解锁了该部分。詹宁斯(1916)检查了一个涉及X染色体的病例AA公司给一个男性一带有唯一X染色体的Y一.后代之间随机交配,持续n个几代人。等位基因的频率是多少A类在女性和男性中n个新形成的第几代生产线?
我们简化了詹宁斯方程,改变了他第58段中的指数,这样,就像我们的混合模型一样,其中第1代是混合种群存在的第一代n个第h代包括初始交叉的结果,后跟n个-1代随机交配。使用我们的符号,Jennings得到了频率第页n个,f等位基因的A类女性和相应频率第页n个,米男性詹宁斯的等位基因频率完全符合方程式1和2中的预期混合比例,在所有女性都来自源1的特殊情况下(秒如果=1),所有男性均来自来源2(秒米= 0). 换句话说,在一代人n个,X染色体等位基因的频率A类在繁殖系统中,雌性和雄性AA公司被交给一个男性一Y、 随机交配持续n个−1代,正好等于混合模型中源1雌性和雄性的预期X染色体混合水平,在混合模型中,雌性从源1进入,雄性从源2进入(表1).
| 模型特征. | 外加剂型号. | 育种系统模型. |
|---|
| 首字母“十字架” | 混合种群由源种群1的雌性和源种群2的雄性组成 | 女性之间发生交叉AA公司和一只雄性一Y(Y) |
|
| 初次交配后的交配类型 | 混合种群中发生随机交配 | 后代之间随机交配 |
|
| 首次杂交后交配的世代数 | n个− 1 | n个− 1 |
|
| 关注人群描述 | 混合隔离下的混合种群 | 自交系 |
|
| 为人口计算的数量 | 世代混合群体中源群体1的预期X染色体混合比例n个,雌性(第页n个,f)和男性(第页n个,米) | X染色体等位基因频率A类在自交系世代中n个,雌性(第页n个,f)和男性(第页n个,米) |
|
| 数学结果一 | 第页n个,f=J型n个+1个/2n个 | 第页n个,f=J型n个+1个/2n个 |
|
| 第页n个,米=J型n个/2n个−1 | 第页n个,米=J型n个/2n个−1 |
|
| 来源 | Goldberg和Rosenberg(2015年,方程式12和13) | 詹宁斯(1916年,第58段) |
| 模型特征. | 外加剂型号. | 育种系统模型. |
|---|
| 首字母“十字架” | 混合种群由源种群1的雌性和源种群2的雄性组成 | 女性之间发生交叉AA公司和一只雄性一Y(Y) |
|
| 初次交配后的交配类型 | 混合种群中发生随机交配 | 后代之间随机交配 |
|
| 首次杂交后交配的世代数 | n个− 1 | n个− 1 |
|
| 关注人群描述 | 混合隔离下的住院人群 | 近交系 |
|
| 人口计算数量 | 世代混合群体中源群体1的预期X染色体混合比例n个,雌性(第页n个,f)和男性(第页n个,米) | X染色体等位基因频率A类在自交系世代中n个,雌性(第页n个,f)和男性(第页n个,米) |
|
| 数学结果一 | 第页n个,f=J型n个+1个/2n个 | 第页n个,f=J型n个+1个/2n个 |
|
| 第页n个,米=J型n个/2n个−1 | 第页n个,米=J型n个/2n个−1 |
|
| 来源 | Goldberg和Rosenberg(2015年,方程式12和13) | 詹宁斯(1916年,第58段) |
| 模型特征. | 外加剂型号. | 育种系统模型. |
|---|
| 首字母“十字架” | 混合种群由源种群1的雌性和源种群2的雄性组成 | 女性之间发生交叉AA公司和一只雄性一Y(Y) |
|
| 初次交配后的交配类型 | 混合种群中发生随机交配 | 后代之间随机交配 |
|
| 首次杂交后交配的世代数 | n个− 1 | n个− 1 |
|
| 关注人群描述 | 混合隔离下的住院人群 | 近交系 |
|
| 人口计算数量 | 世代混合群体中源群体1的预期X染色体混合比例n个,雌性(第页n个,f)和男性(第页n个,米) | X染色体等位基因频率A类在自交系世代中n个,雌性(第页n个,f)和男性(第页n个,米) |
|
| 数学结果一 | 第页n个,f=J型n个+1个/2n个 | 第页n个,f=J型n个+1个/2n个 |
|
| 第页n个,米=J型n个/2n个−1 | 第页n个,米=J型n个/2n个−1 |
|
| 来源 | Goldberg和Rosenberg(2015年,公式12和13) | 詹宁斯(1916年,第58段) |
| 模型特征. | 外加剂型号. | 育种系统模型. |
|---|
| 首字母“十字架” | 混合种群由源种群1的雌性和源种群2的雄性组成 | 女性之间发生交叉AA公司和一只雄性一Y(Y) |
|
| 初次交配后的交配类型 | 混合种群中发生随机交配 | 后代之间随机交配 |
|
| 首次杂交后交配的世代数 | n个− 1 | n个− 1 |
|
| 关注人群描述 | 混合隔离下的住院人群 | 近交系 |
|
| 人口计算数量 | 世代混合群体中源群体1的预期X染色体混合比例n个,雌性(第页n个,f)和男性(第页n个,米) | X染色体等位基因频率A类在自交系世代中n个,雌性(第页n个,f)和男性(第页n个,米) |
|
| 数学结果一 | 第页n个,f=J型n个+1个/2n个 | 第页n个,f=J型n个+1个/2n个 |
|
| 第页n个,米=J型n个/2n个−1 | 第页n个,米=J型n个/2n个−1 |
|
| 来源 | Goldberg和Rosenberg(2015年,方程式12和13) | 詹宁斯(1916年,第58段) |
从数学上讲,这些计算回答了同一个问题:虽然我们计算X染色体位点上随机女性或男性的源种群混合,但詹宁斯计算X染色体座位的“源种群”的“混合”,包括单个AA公司女性。根据詹宁斯的方案,等位基因的频率A类记录提供雌性的来源种群的混合比例。从某种意义上说,一个世纪前处于该领域前沿的纯合性和杂合性水平的育种系统计算是现在流行的现代混合计算的一个版本。
观点
同一数列的两个问题的出现J型n个是确定相隔99年的相似车型系列之间联系的关键。然而,问题仍然存在:混合模型和繁殖系统模型之间的其他联系在遗传学记录,他们能为现代混合物研究带来什么见解?混合作为种群间杂交的一种形式的类比是常见的。但在某些情况下,这种平行性不仅仅是一种类比,因为混合和繁殖系统模型可以使用完全相同的数学。
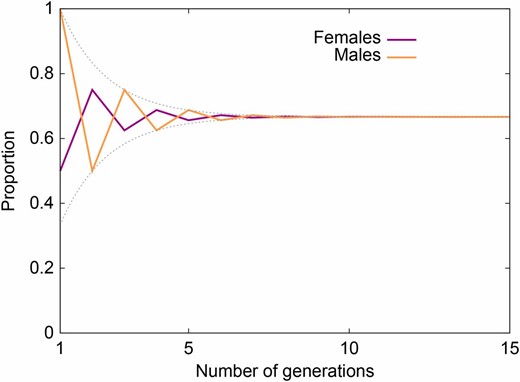
除了与混合模型的关系之外詹宁斯(1916)值得注意的还有其他一些原因,包括早期的分类交配模型和早期针对有害隐性选择的单基因座分析。这可能是人们最常记住的,因为它确定了X染色体上哈迪-温伯格平衡方法中的振荡。因为每个男性X染色体在过去的一代中都是女性的,所以男性的等位基因频率“追赶”女性的频率,滞后一代。与常染色体不同,平衡是随着时间的推移而达到的,而不是立即实现的。与此模式的对应关系也可以在Goldberg和Rosenberg(2015):特定源总体的预期混合遵循相同的振荡(图2). 事实上,等式3和4精确地给出了等位基因的女性和男性频率A类代内n个对于仅从第0代开始的种群,在接近Hardy–Weinberg平衡时≥1AA公司女性和一Y男性(Crow和Kimura 1970年第46页)。与Hardy–Weinberg设置平行,与常染色体位点不同(戈德伯格等。2014)在杂交分离条件下,随机雌雄X染色体上的期望混合并非随时间而恒定。
图2
X染色体模型中女性和男性的混合比例和等位基因频率。绘制的值可以等效地解释为:(1)源群体1中随机雌性和随机雄性混合体的X染色体比例n个在混合模型中,第1代中的后代个体的母亲都从源种群1进入,父亲都从源群体2进入(Goldberg和Rosenberg,2015年); (2) 生成中n个,X染色体等位基因在女性和男性中的频率A类在一个十字架之后AA公司女性和一个一Y男性,其次是n个−随机交配1代(詹宁斯1916); 和(3)代n个X染色体等位基因在女性和男性中的频率A类之后n个随机交配的第几代,从第0代开始AA公司女性和一Y男性(詹宁斯1916). 女性的数值由方程式3给出,男性的数值由公式4给出。注意Jacobsthal数的闭式表达式是J型n个= [2n个− (−1)n个]/第三代n个,绘制的两个量都受到下限(2)的约束n个− 1)/(3 × 2n个−1)和上界(2n个+ 1)/(3 × 2n个−1).
一些早期的教科书,包括Crow和Kimura(1970)和李(1976),讨论X染色体Hardy–Weinberg振荡。然而,在后来的书中,振荡模式及其与递归序列的连接有时会丢失,在某些情况下,参考詹宁斯(1916).
从H.S.Jennings的角度来看,Crow(1987)评论说詹宁斯(1916)在育种系统中,很快就被Sewall Wright对任意系谱产生的近亲繁殖系数的一般方法所取代。然而,这篇文章揭示了Hardy–Weinberg模型的X染色体行为,预测了现代混合研究中感兴趣的问题,并且有助于扩展斐波那契数的长长的生物学示例列表。
在遗传学档案中充满了惊喜。遗传学据报道,布里奇斯的手稿于1915年10月21日收到,詹宁斯的手稿于8月26日收到。人们很容易推测遗传学如果编辑们在安排杂志第一期的出版顺序时没有颠倒到达时间,情况可能会有所不同。
致谢
我感谢编辑、匿名审稿人和艾米·戈德伯格的评论,这些评论大大改进了手稿。
引用文献
盆地
S公司
,
1963
自然界中出现的斐波那契数列。
斐波那契季刊
1
:53
——56
.桥梁
C B类
,
1916
非双联作为遗传染色体理论的证据。
遗传学
1
:1
——52
.乌鸦
J F公司
,
1987
70年前遗传学:H.S.Jennings和近亲繁殖理论。
遗传学
115
:389
——391
.乌鸦
J F公司
,木村
M(M)
,
1970
群体遗传学理论导论
.伯吉斯
,明尼阿波利斯
.杜兴
P(P)
,伊夫科维奇
D类
,赫特
S公司
,斯蒂芬
W公司
,劳伦特
S公司
,
2013
人口统计推断揭示了北美地区非洲和欧洲的混合黑腹果蝇人口。
遗传学
193
:291
——301
.加内茨基
B和S
,霍利
俄罗斯
,
2016
百年诞辰遗传学:通向未来的桥梁。
遗传学
202
DOI:10.1534/genetics.115.180182。戈德伯格
A类
,罗森博格
不适用
,
2015
超过2/3和1/3:X染色体上性别偏见混合物的复杂特征。
遗传学
201
:263
——279
.戈德伯格
A类
,贝尔杜
P(P)
,罗森博格
不适用
,
2014
常染色体混合水平是混合人群性别偏见的信息来源。
遗传学
198
:1209
——1229
.砾石
S公司
,
2012
当地祖先的群体遗传学模型。
遗传学
191
:607
——619
.詹宁斯
H S公司
,
1914
近亲繁殖结果的公式。
美国国家。
48
:693
——696
.詹宁斯
H S公司
,
1916
不同繁殖系统的数值结果。
遗传学
1
:53
——89
.Johnston,M.,2014年 在GSA期刊上。GSA报告人2014年冬季,第10-11页。
锂
C C公司
,
1976
人口遗传学第一课程
.Boxwood出版社
,加利福尼亚州太平洋格罗夫
.梁
M(M)
,尼尔森
对
,
2014
混合区的长度。
遗传学
197
:953
——967
.卢
P-R公司
,利普森
M(M)
,帕特森
N个
,穆尔贾尼
P(P)
,皮克雷尔
J K公司
等,
2013
利用连锁不平衡推断人类混合历史。
遗传学
193
:1233
——1254
.洛赫穆勒
英国
,巴斯塔曼特
C、D
,克拉克
A G公司
,
2011
在非洲裔美国人中检测最近混合情况下的定向选择。
遗传学
187
:823
——835
.洛泽
K(K)
,弗兰茨
L A F公司
,
2014
三个基因组的最大似然分析证实了尼安德特人在欧亚大陆的混血。
遗传学
196
:1241
——1251
.长
J C公司
,
1991
混合种群的遗传结构。
遗传学
127
:417
——428
.帕特森
N个
,穆尔贾尼
P(P)
,罗
Y(Y)
,马利克
S公司
,罗兰
N个
等,
2012
人类历史中古老的混合体。
遗传学
192
:1065
——1093
.珍珠
对
,
1913
对近亲繁殖问题分析的贡献。
美国国家。
47
:577
——614
.普罗文
宽B
,
1971
理论群体遗传学的起源
.芝加哥大学出版社
,芝加哥
.桑德森
J型
,Sudoyo公司
H(H)
,卡拉费特
T月
,锤子
M F公司
,考克斯
百万富翁
,
2015
利用小波变换从局部基因组祖先重建过去的混合过程。
遗传学
200
:469
——481
.塞奇法尔
A类
,布兰德万
Y(Y)
,拉尔夫
P(P)
,笼子
G公司
,
2015
次级接触区基因组的空间混合。
遗传学
201
:243
——261
.贝尔杜
P(P)
,罗森博格
不适用
,
2011
杂交种群混合历史的一般力学模型。
遗传学
189
:1413
——1426
.赖特
S公司
,
1969
进化与种群遗传学第2卷:基因频率理论
芝加哥大学出版社
,芝加哥
.
©遗传学2016







{kind=link}
{kind=link}