摘要
研究表明,过量的一氧化氮(NO)会对大脑产生神经毒性影响。此外,抑制两种一氧化氮合成酶,即神经元型一氧化氮合酶(nNOS)和诱导型一氧化氮合酶类(iNOS),在帕金森病(PD)的MPTP模型中显示出神经保护作用。这些数据表明,NOS可能参与控制黑质多巴胺能神经元对环境损伤的抵抗。因此,我们调查了这些基因中存在的多态性是否会导致患帕金森病的风险。我们在法国农业联合会(MutuitéSociale Agricole)的受试者中进行了一项基于社区的病例对照研究,该组织是法国农业工人健康保险组织。本研究纳入了29名帕金森病患者和488名欧洲血统(主要是法国血统)的对照组,他们的年龄、性别和居住地区相匹配。观察到与iNOS第22外显子多态性相关(AA携带者OR=0.50,95%CI=0.29–0.86,P(P)=0.01)和nNOS外显子29(T等位基因携带者的OR=1.53、95%CI=1.08–2.16,P(P)=0.02); 未观察到与nNOS第18外显子多态性相关(T等位基因携带者OR=1.20,95%CI=0.85–1.69,P(P)=0.30). 此外发现nNOS多态性与当前和曾经吸烟有显著的相互作用(nNOS 18,P(P)=0.05; nNOS 29、,P(P)=0.04). 总之,这些数据支持这两个基因作为新的修饰基因参与PD。
简介
帕金森病(PD)是继阿尔茨海默病之后最常见的神经退行性疾病。在过去的10年里,三个不同基因(α-突触核蛋白、帕金和泛素C末端水解酶)的突变鉴定以及至少五个其他基因座的定位强调了遗传因素在PD发展中的重要性(1–9)此外,多巴胺能神经毒素MPTP的作用与帕金森病相似,这一观察结果促使研究人员对参与多巴胺途径、线粒体功能、脂蛋白代谢或外源性解毒的候选基因的多态性进行关联研究(10).
一氧化氮(NO)是一种具有多种生理作用的生物信使分子。然而,NO也是一种自由基,可以与超氧阴离子结合形成过氧亚硝酸盐。因此,NO有助于氧化应激并诱导细胞发生多种变化,包括(1)脂质过氧化(2)蛋白质的功能改变(例如亚硝基化)(3)DNA损伤和(4)线粒体能量功能障碍(11). 一氧化氮合酶(NOS)通过精氨酸和氧气催化NO的形成。NOS有三种亚型:神经元型NOS(nNOS);内皮型一氧化氮合酶(eNOS);和诱导型一氧化氮合酶(iNOS)。在这三种亚型中,nNOS和iNOS在帕金森病的神经退行性变和胶质细胞反应中的潜在意义特别相关。事实上,nNOS是神经元和胶质细胞中组成性表达的主要亚型。此外,iNOS可在脑胶质细胞和侵袭巨噬细胞中诱导,以应对各种损伤(12,13).
在人类和动物模型中有多种证据支持NO和NOS在PD病因中的作用。特别是,使用药理物质或基因缺陷小鼠抑制nNOS或iNOS,可以防止PD MPTP模型中多巴胺能神经元的破坏(14–19).
因此,我们在一项基于社区的病例对照研究中探讨了nNOS和iNOS基因多态性与PD之间的关系。此外,我们研究了暴露于农药或吸烟是否改变了这些多态性与PD之间的关系。MPTP是农药百草枯的类似物,而暴露于农药与PD相关(20). 此外,MPTP调节nNOS和iNOS的表达(18,21). 相反,吸烟与PD呈负相关,并可能影响NO的生成(22). 我们的数据表明nNOS和iNOS基因都与PD的发生有关。
结果
292名受试者向农业互助协会(MSA)申请免费的PD健康保险。其中,184名患者由神经科医生直接检查:153名患者被发现患有帕金森病,29名患者患有其他原因的帕金森症,两名患者不符合帕金森症标准。在剩下的108名患者中,要求免费医疗保险的神经科医生为86名患者提供了临床信息:他确认了72名PD患者的诊断,并诊断出14名帕金森综合征的另一原因。没有获得22名患者的详细临床信息。然而,根据MSA医生的临床检查,所有人都符合帕金森病的标准,并且没有人在帕金森病发作前使用过神经抑制剂;因此,我们保留了它们进行分析。247名患者(153+72+22)与676名对照组(194名四胞胎、41名三胞胎、12对)进行配对;89%的帕金森病患者和84%的匹配对照接受了血液取样进行DNA提取。此外,排除了五名患者和七名至少有一名父母在欧洲以外出生的对照组。最终样本包括209名患者和488名匹配的对照组(108名四胞胎、63名三胞胎、38对)。它们的一般特征如表所示1.
关于两个候选基因中三个双等位基因多态性的结果如表所示2.对照人群的所有三个多态性(iNOS)均处于Hardy–Weinberg平衡 22,P(P)=0.45; 无操作系统 18,P(P)=0.47; 无操作系统 29,P(P)=0.56).
iNOS 22多态性的AA纯合子在对照组中比患者中更常见(OR=0.56,95%CI=0.32–1.01;P(P)=0.05),而两组之间的杂合子频率没有显著差异。假设a等位基因的隐性效应,AA携带者的OR为0.50(95%CI=0.29–0.86;P(P)=0.01)与其他两种基因型相比。这种反向关联在男性中更强(OR=0.40,95%CI=0.17–0.92;P(P)=0.03),高于女性(OR=0.61,95%CI=0.30–1.23;P(P)=0.16); 然而,性别与iNOS 22之间的交互作用并不显著(P(P)=0.49). 当仅考虑病例和匹配的无PD家族史的对照组时,也获得了类似的结果(AA携带者的OR=0.48,95%CI=0.27–0.86;P(P)=0.01)或父母均在法国出生(AA携带者or=0.53,95%CI=0.30–0.93;P(P)=0.03),年轻发病PD患者(≤45岁,n个=4和10个匹配的对照组)被排除在分析之外(AA携带者的OR=0.47,95%CI=0.27–0.82;P(P)=0.01). 帕金森病发病时的年龄并没有改变这种联系。(数据未显示。)
患者和对照组之间nNOS 18多态性的基因型分布没有统计学上的显著差异(表2).
相反,患者和对照组之间nNOS 29多态性的基因型分布不同(P(P)=0.01). 杂合子和TT纯合子都有较高的PD风险。OR随着T等位基因的数量增加而增加(P(P)趋势=0.004)。至少有一个T等位基因携带者的OR为1.53(95%CI=1.08–2.16;P(P)=0.02). 根据共显性(484.281)和显性(486.868)模型的Akaike信息准则,共显性模型是最佳拟合模型。我们再次注意到,多态性与帕金森病之间的相关性在男性中更强(OR=1.84,95%CI=1.15-2.94;P(P)=0.01),高于女性(OR=1.21,95%CI=0.72-2.04;P(P)=0.47),尽管性别与nNOS 29之间的相互作用在统计上不显著(P(P)=0.23)。无PD家族史的病例和匹配对照组也获得了类似结果(T携带者OR=1.56,95%CI=1.08–2.26;P(P)=0.02),或父母均在法国出生(T携带者or=1.58,95%CI=1.10–2.26;P(P)=0.01),当年轻型PD患者被排除在分析之外时(T携带者OR=1.52,95%CI=1.07–2.15;P(P)=0.02)。帕金森病发病时的年龄并未改变这种联系(数据未显示)。
构建了两个nNOS多态性的单倍型(表3). 在患者中,有过多的单倍型,包括nNOS 29的T等位基因;这种过剩似乎与nNOS 18等位基因无关(nNOS 18-nNOS 29单倍型C-T和T-T与所有其他单倍型的病例对照比较:OR=1.39,95%CI=1.07–1.79,P(P)=0.009).
我们探讨了nNOS 29和iNOS 22多态性之间相互作用的假设(表4). 我们的数据表明,iNOS 22对nNOS 29 TT或CT携带者的保护作用弱于CC纯合子,换句话说,nNOS 22携带者对AA iNOS 29的保护作用强于GA或GG携带者(观察到的联合效应or=0.97,而预期or为0.29 × 1.41=0.41). 然而,交互测试并不显著(P(P)=0.16). 在一个包括多态性作为主要影响但没有交互作用的多变量模型中,PD和nNOS 29之间的关联(T携带者的OR=1.53,95%CI=1.08–2.17;P(P)=0.02)和iNOS 22(AA携带者OR=0.48,95%CI=0.28–0.83;P(P)=0.01)仍然显著。
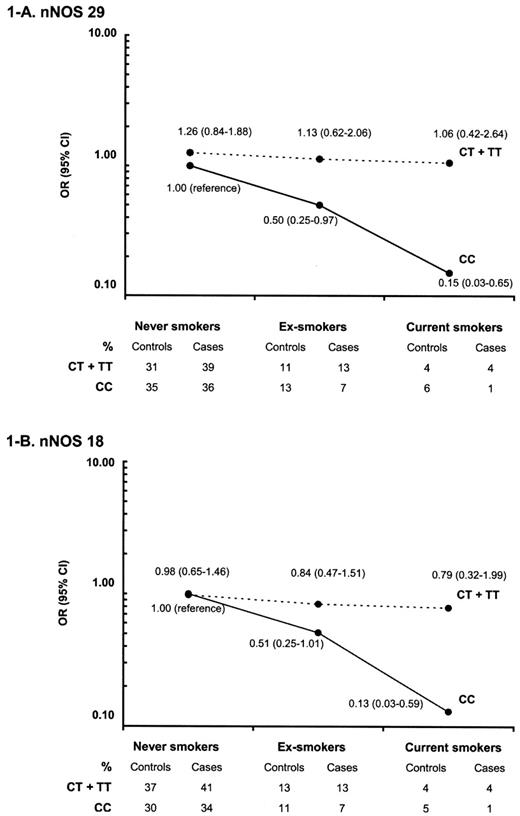
由于吸烟与该人群的PD呈负相关(表1),我们测试了nNOS基因的两个多态性与吸烟之间的相互作用。观察到相互作用,如图所示1对于这两种nNOS多态性,在多态性T等位基因的携带者中,PD和吸烟之间的反向关联丢失,而在C等位基因纯合子中存在这种反向关联(当前吸烟的影响大于戒烟)。多态性与当前吸烟之间的相互作用具有统计学意义(nNOS 18,P(P)=0.04; nNOS 29、,P(P)=0.04),而多态性与吸烟之间的相互作用在统计学上不显著(nNOS 18,P(P)=0.22; nNOS 29、,P(P)=0.17). 如果将以前吸烟和现在吸烟归为一组(曾经吸烟),那么曾经吸烟和nNOS 18之间的相互作用(P(P)=0.05)和nNOS 29(P(P)=0.04)多态性仍然显著。
最后,由于农药暴露与PD呈正相关,我们调查了农药暴露与多态性的联合效应,发现农药暴露与无多态性之间没有显著交互作用(数据未显示)。
讨论
在一项主要由散发患者组成的大型社区PD病例对照研究中,我们对两个NOS基因(nNOS和iNOS)的三个多态性进行了基因分型。我们发现,与对照组相比,帕金森病患者中至少有一个nNOS 29 T等位基因携带者的显著过度表达和AA iNOS 22纯合子的低表达,这是最新报道的多态性。在帕金森病和nNOS 18多态性之间没有观察到相关性。此外,nNOS 18和nNOS 29可能调节吸烟与PD的关系。
这些结果表明,nNOS和iNOS可能在散发性PD的易感性中起作用。这与动物或人类的一些研究一致。在动物中,nNOS抑制剂对MPTP具有保护作用,并且缺乏nNOS基因的突变小鼠比野生型小鼠对MPTP更有抵抗力(14–17,23). iNOS的类似实验表明,这种酶也在多巴胺能神经元的敏感性中发挥作用(18,19,24).
也有证据表明,PD患者存在NO过度生成以及nNOS和iNOS的影响。与对照组相比,经治疗和未经治疗的PD患者的脑脊液中亚硝酸盐浓度增加(25),并且与对照组相比,帕金森病患者的中性粒细胞中nNOS过表达(26). 使用光谱在黑质帕金森病患者与对照组的比较(27),并且在黑质尸检PD样本,但年龄匹配的对照样本中没有(28). PD患者路易小体中观察到NO形成标记物(例如硝基酪氨酸免疫反应)(29)和MPTP诱导的帕金森综合征患者。最后,用于治疗PD的溴隐亭是大鼠nNOS的强抑制剂,在较小程度上是小鼠iNOS的抑制剂(30).
我们没有发现nNOS 29和iNOS 22多态性之间有统计学意义的相互作用。然而,iNOS 22对nNOS 29 TT或CT携带者的保护作用弱于对CC纯合子的保护作用,我们的研究可能没有足够的能力检测小尺寸的相互作用。当其中一个多态性的影响被调整为另一个的影响时,每个多态性都与PD相关。这些发现表明这两个多态性具有独立的影响。
nNOS基因中的两个多态性似乎调节了吸烟的保护作用。据报道,在产后胎盘中,吸烟仅与eNOS多态性罕见等位基因携带者中eNOS表达降低相关(31). 尼古丁注射或暴露于香烟烟雾中会导致大鼠和兔子大脑中NO和/或NO代谢物的增加,这可能是通过NOS的作用介导的(32,33). 在本研究中,我们观察到,对于nNOS 29和nNOS 18,在多态性T等位基因携带者中,吸烟与PD之间的反向关系消失。这一观察结果与吸烟对nNOS产生NO的影响受nNOS多态性调控的假设一致,但这一假设需要得到证实。
MPTP可增加nNOS和iNOS的表达(18,19,21). 然而,我们没有发现有证据支持农药暴露与本研究中测试的多态性之间的相互作用。因此,如果杀虫剂的作用是通过NO生成的增加(如MPTP所观察到的那样)介导的,那么这种增加可能与这些多态性无关。
由于这些多态性的功能尚不清楚,仍需确定iNOS 22和nNOS 29多态性与PD的关联是否是导致PD易感性的始发事件。另一种可能性是,它们与另一个功能多态性或附近的另一个基因存在连锁不平衡。iNOS 22是一个外显子多态性,但不改变氨基酸序列。位于iNOS基因启动子的另一个重复多态性与路易体痴呆有关(34)这是一种以脑内存在路易体为特征的疾病。nNOS和iNOS基因都与NO代谢有关,并且在本研究中发现与PD相关,这一事实与另一个基因的含义相矛盾。然而,已观察到与PD呈正相关的Tau基因位于15以下 染色体17q上iNOS基因的cM(35–37). 晚发性帕金森病基因组筛查的最有力证据位于7 来自iNOS基因的cM(38). 因此,我们调查了Tau和iNOS 22之间的连锁不平衡,并调整了我们对Tau基因型PD和iNOS22之间关系的分析。Tau和iNOS 22在病例中没有连锁不平衡(D类′=0.05)和对照(D类′=0.006),调整后OR保持不变(未显示数据)。因此,PD和iNOS之间的关联独立于Tau基因。
nNOS 29位于5′非翻译区(UTR)的外显子。该区域通常与转录物和mRNA稳定性的调节有关。这种多态性或另一种位于基因表达调控区的多态性可能介导这种效应。这个问题很难解决,因为nNOS表达的调节非常复杂,有超过12个转录物(通过选择性剪接或启动子使用获得),并且可能被抗ens RNA转录物抑制表达。最近有报道称PD与nNOS基因启动子的另一个重复多态性有关。64名中国患者和对照组的等位基因大小分布存在显著差异(39).
如果患者和对照组之间的种族背景存在差异,以及根据种族背景的等位基因频率变化,则关联研究可能存在人群分层偏见。在这项研究中,只有父母两人出生在欧洲的受试者被包括在内。大多数参与者的父母都在法国出生,仅限于该亚组的分析得出了与总体结果类似的结果。此外,对于nNOS 29和nNOS 18多态性,我们发现其频率与美国高加索人群中报告的频率相似(40). 因此,人口分层不太可能对我们的研究结果产生偏见。
总之,我们的研究结果表明nNOS和iNOS基因是PD的易感基因,nNOS多态性改变了吸烟与PD的关系。为了在其他人群中证实这种关联,并破译所研究多态性的功能,还需要进行进一步的研究。
材料和方法
参与者
这项研究是在法国社会保障组织MSA的受试者中进行的,该组织负责向与农业有关的工人(即农民和农场工人,但也包括筒仓、农业合作社和种子店的工人,以及第三产业专业人员)报销与健康相关的费用研究方案已在其他地方详细描述(41).
简言之,18至75岁的个人在18个月内提交了PD完全医疗保险福利申请,他们被邀请参加。作为研究方案的一部分,他们由具有运动障碍经验的神经学家进行检查。当无法直接检查患者时,会联系患者的治疗神经科医生,并要求其填写临床问卷。帕金森综合征被定义为出现四个主要症状中的两个或多个:静止性震颤、运动迟缓、僵硬和姿势反射受损(42,43). 在排除以下标准后,PD被定义为帕金森综合征的存在:未另行解释的更广泛神经系统受累的显著或早期迹象;核上凝视麻痹;反复卒中伴帕金森综合征逐步进展;发病前接受抗精神病药物治疗;对左旋多巴无反应(42,43). 已经受益于痴呆症或要求免费医疗保险的患者或卧床不起的患者不包括在内。
在所有MSA附属机构中招募对照组,这些附属机构要求报销医疗费用(例如康复、眼镜、牙科护理、足部护理、水疗、护理、呼吸功能测试、动态心电图监测)。研究时,每个病例最多有三个对照组,按年龄(±2岁)、性别和居住地区匹配。如果参与者已经受益于PD或痴呆的免费医疗保险,接受了PD治疗,符合帕金森综合征标准,或卧床不起,则不将其纳入对照组。
获得了有关人口统计学数据、合并症、PD家族史以及父母双方出生国家的信息。一份面对面的问卷收集了有关吸烟的详细信息,包括每天平均吸烟次数以及开始吸烟和停止吸烟的年龄。此外,如果参与者一生中每天吸烟的次数不同,则要求他们将吸烟史划分为最多三个时间段。根据这一信息,参与者根据其在PD发病时的吸烟状态(患者中)或指数年龄(对照组中;定义为匹配病例中的PD发病年龄)被分为当前吸烟者、前吸烟者或从未吸烟者。直到帕金森病发作(患者)或指数年龄(对照组)为止的累计吸烟次数为每天吸烟次数(每天吸烟次数除以20)乘以吸烟年数;我们的计算考虑了根据上述时间段每天吸烟数量的变化。
使用两阶段逐案接触评估程序评估农药接触情况,该程序包括使用自填问卷进行专业接触筛查,对于根据该筛查进行专业接触的个人,在家中与MSA的职业健康医生进行详细访谈(41,44,45). 医生使用特定的问卷,填写职业史,并确认参与者是否在专业环境中亲自喷洒或配制过杀虫剂;根据与医生的访谈,在自填问卷中宣布为专业杀虫剂使用者的人中,有20%不是专业使用者(11%是专门用于园艺的使用者,9%从未使用过)。最后,调查问卷由一个不了解病例或对照状态的专家小组进行审查,我们能够将参与者分为三组,即从未接触杀虫剂的人、专门从事园艺的人和专业使用者。我们进行了一项小型验证研究,通过电话采访从那些被归类为从未使用过的人中随机选择的受试者(n个=15)或仅用于园艺的用户(n个=15); 自填问卷和电话采访总是一致的,这30名受试者中没有一人曾在专业环境中使用过杀虫剂。
克里姆林宫医院伦理委员会(巴黎)批准了该研究方案,所有受试者都签署了知情同意书。
基因分型
对参与者进行血液取样,并使用标准技术从外周血白细胞中提取基因组DNA。
对于iNOS基因(NOS2A),我们研究了外显子22(iNOS 22)中37498位的G/a替换,该替换不会导致氨基酸变化(PubMed,加入NT_024889),但会产生Btg公司I限制酶位点。引物为5′-CTGGCTGTTCCTGCTTTCC-3′和5′-CTCGGTGTAGGTGACC-3′。DNA最初变性10 在95°C下最小,然后接地程序:在95°C:12次循环 最小值,72–66°C,持续30 s、 72°C,持续30 s和23个循环,在95°C下进行1次 最低温度,66°C,持续30 s、 72°C,持续30 s,最后是10 72°C时最小值。在3%琼脂糖凝胶电泳后,可以看到限制性片段:79、24和23的三个片段 bp定义了G等位基因的存在,以及103和23的两个片段 bp A等位基因的存在。
我们还对由Grasemann鉴定的nNOS基因的两个T/C多态性进行了基因分型等。外显子18(nNOS 18)和外显子29(nNOS29)(40). 先前已经描述了用于外显子18的引物和PCR条件。外显子18的多态性废除了a德拉III限制场所。C等位基因由124和55的两个片段定义 bp和T等位基因的片段179 bp。我们为第29外显子的多态性设计了以下一组引物:5′-TTGGTTTTCCTGCTGATGT-3′和5′-GCTGTGCTAGTCCTGCA-3′。PCR条件与用于nNOS 18的条件相同。突变产生了Nla公司III限制性位点。C等位基因由102、69和16的三个片段定义 bp和T等位基因的四个片段94、69、16和8的存在 bp。
统计方法
为了最大限度地降低人口分层偏见的风险,仅纳入父母双方均在欧洲出生的病例和对照。与匹配设计一致,进行匹配分析,并使用比值比(OR)估计相对风险。对于每个研究变量,我们计算了OR、95%置信区间(CI)和P(P)-值,对具有可变数量对照的匹配集使用条件逻辑回归。条件logistic回归模型也用于调查相关变量的独立效应,调整潜在的混杂变量,并测试乘法交互作用。
为了比较病例和对照组的基因型分布,分别计算了杂合子和多态性等位基因纯合子的ORs,并通过在模型中包括虚拟变量和将野生等位基因的纯合子作为参考组来进行一般关联检验。此外,我们计算了P(P)-趋势值,包括对应于多态性等位基因数量的序数变量(共显性模型)。还进行了假设显性模型(nNOS 29和nNOS 18)或隐性模型(iNOS 22)的分析。为了比较nNOS 29多态性的显性和共显性模型,我们使用了Akaike信息标准,因为这两个模型没有嵌套。
在Hardy–Weinberg平衡假设下,使用E-M算法从基因型数据估计两个nNOS多态性的单倍型频率。计算总体似然比统计来比较病例和对照组之间nNOS单倍型的总体分布,并使用排列测试单个单倍型频率的差异(n个=10,000) (46). 连锁不平衡系数(D类′)使用标准公式计算两个nNOS多态性(47).
SAS软件版本6.11用于统计分析(SAS institute,Cary,NC,USA)。
致谢
我们感谢所有与参与者面谈的MSA(MutualéSociale Agricole)医生以及帮助协调研究的MSA员工。David Gervais博士和Lamia Benslamia博士参与了PD患者的检查。Chantal Cordelier女士和Carline Amiel博士、Frédérique Brunner博士和Jean-Philippe Galanaud博士帮助评估了农药接触情况。Safia Zenagui博士参与了该研究的协调工作。我们感谢Xavier Hermant和Vincent Mouroux的技术援助。本研究得到了INSERM(国家圣母研究所)、里尔巴斯德研究所、MSA和法国环境部的支持。CL是技术部的接受者。
图1。nNOS 29之间的相互作用(A类)和nNOS 18(B类)nNOS基因多态性与吸烟。表格显示了每个接触类别的频率,数字显示了吸烟的个体和联合影响以及nNOS基因中nNOS 29(A)和nNOS 18(B)多态性的比值比(95%置信区间)。实线对应CC载波,虚线对应CT和TT载波。(A)P(P)-nNOS 29与当前吸烟之间的相互作用值,0.04;不包括吸烟,0.17。(B)P(P)-nNOS 18与当前吸烟量之间的相互作用值为0.04;不包括吸烟,0.22。
| 一般特征. | 控件(n个=488). | 案例(n个=209). | OR(95%置信区间)一. | P(P) 一. |
|---|
| PD发病年龄,中位数(范围) | — | 65 (35–75) | — | — |
| 研究时的年龄,中位数(范围) | 68 (36–79) | 68 (37–75) | — | 0.89 |
| 男性,%(n个) | 59 (289) | 57 (119) | — | — |
| 父母出生国,%(n个) | | | | |
| 父母均出生于法国 | 93 (443) | 92 (190) | 1.00(参考) | — |
| 至少有一位父母在另一个欧洲国家出生 | 7 (33) | 8 (16) | 1.24 (0.63–2.43) | 0.54 |
| 未知 | (12) | (3) | | |
| PD家族史,%(n个) | | | | |
| 否 | 95 (451) | 91 (189) | 1.00(参考) | — |
| 是的 | 5(25) | 9 (18) | 1.75 (0.92–3.22) | 0.09 |
| 未知 | (12) | (2) | | |
| 吸烟,%(n个) | | | | |
| 从未 | 67(323) | 75 (155) | 1.00(参考) | — |
| 戒烟者 | 24 (115) | 20 (42) | 0.71 (0.46–1.11) | 0.13 |
| 当前吸烟者 | 9 (45) | 5 (10) | 0.42 (0.20–0.89) | 0.02 |
| 曾经吸烟者(现任和前任吸烟者) | 33 (160) | 25 (52) | 0.63 (0.42–0.96) | 0.03 |
| ≤18包年b条 | 15 (75) | 15 (31) | 0.82 (0.49–1.38) | 0.46 |
| >18包年b条 | 18 (85) | 10 (20) | 0.42 (0.23–0.75) | 0.003c(c) |
| 未知 | (5) | (2) | | |
| 农药使用,%(n个) | | | | |
| 否 | 32 (143) | 29 (58) | 1.00(参考) | — |
| 用于园艺 | 21 (93) | 22 (43) | 1.52(0.88–2.61) | 0.13 |
| 专业 | 47 (213) | 49 (98) | 1.64 (0.94–2.84) | 0.08 |
| 用于园艺或专业 | 68 (306) | 71 (141) | 1.57 (0.98–2.53) | 0.06 |
| 未知 | (39) | (10) | | |
| 一般特征. | 控件(n个=488). | 案例(n个=209). | OR(95%置信区间)一. | P(P) 一. |
|---|
| PD发病年龄,中位数(范围) | — | 65 (35–75) | — | — |
| 研究时的年龄,中位数(范围) | 68 (36–79) | 68 (37–75) | — | 0.89 |
| 男性,%(n个) | 59 (289) | 57 (119) | — | — |
| 父母出生国,%(n个) | | | | |
| 父母均出生于法国 | 93 (443) | 92 (190) | 1.00(参考) | — |
| 至少有一位父母在另一个欧洲国家出生 | 7 (33) | 8 (16) | 1.24 (0.63–2.43) | 0.54 |
| 未知 | (12) | (3) | | |
| PD家族史,%(n个) | | | | |
| 否 | 95 (451) | 91 (189) | 1.00(参考) | — |
| 是的 | 5 (25) | 9 (18) | 1.75(0.92–3.22) | 0.09 |
| 未知 | (12) | (2) | | |
| 吸烟,%(n个) | | | | |
| 从未 | 67 (323) | 75 (155) | 1.00(参考) | — |
| 戒烟者 | 24 (115) | 20 (42) | 0.71 (0.46–1.11) | 0.13 |
| 当前吸烟者 | 9 (45) | 5 (10) | 0.42 (0.20–0.89) | 0.02 |
| 曾经吸烟者(现任和前任吸烟者) | 33 (160) | 25 (52) | 0.63 (0.42–0.96) | 0.03 |
| ≤18包年b条 | 15 (75) | 15 (31) | 0.82 (0.49–1.38) | 0.46 |
| >18包年b条 | 18 (85) | 10 (20) | 0.42 (0.23–0.75) | 0.003c(c) |
| 未知 | (5) | (2) | | |
| 农药使用,%(n个) | | | | |
| 否 | 32 (143) | 29 (58) | 1.00(参考) | — |
| 用于园艺 | 21 (93) | 22(43) | 1.52 (0.88–2.61) | 0.13 |
| 专业 | 47 (213) | 49 (98) | 1.64(0.94–2.84) | 0.08 |
| 用于园艺或专业 | 68 (306) | 71 (141) | 1.57 (0.98–2.53) | 0.06 |
| 未知 | (39) | (10) | | |
| 一般特征. | 控件(n个=488). | 案例(n个=209). | OR(95%置信区间)一. | P(P) 一. |
|---|
| PD发病年龄,中位数(范围) | — | 65 (35–75) | — | — |
| 研究时的年龄,中位数(范围) | 68 (36–79) | 68 (37–75) | — | 0.89 |
| 男性,%(n个) | 59 (289) | 57 (119) | — | — |
| 父母出生国,%(n个) | | | | |
| 父母均出生于法国 | 93 (443) | 92 (190) | 1.00(参考) | — |
| 至少有一位父母在另一个欧洲国家出生 | 7 (33) | 8 (16) | 1.24 (0.63–2.43) | 0.54 |
| 未知 | (12) | (3) | | |
| PD家族史,%(n个) | | | | |
| 否 | 95(451) | 91 (189) | 1.00(参考) | — |
| 是的 | 5 (25) | 9(18) | 1.75 (0.92–3.22) | 0.09 |
| 未知 | (12) | (2) | | |
| 吸烟,%(n个) | | | | |
| 从未 | 67 (323) | 75 (155) | 1.00(参考) | — |
| 戒烟者 | 24 (115) | 20 (42) | 0.71 (0.46–1.11) | 0.13 |
| 当前吸烟者 | 9 (45) | 5 (10) | 0.42 (0.20–0.89) | 0.02 |
| 曾经吸烟者(现任和前任吸烟者) | 33 (160) | 25 (52) | 0.63 (0.42–0.96) | 0.03 |
| ≤18包年b条 | 15 (75) | 15 (31) | 0.82 (0.49–1.38) | 0.46 |
| >18包年b条 | 18 (85) | 10 (20) | 0.42 (0.23–0.75) | 0.003c(c) |
| 未知 | (5) | (2) | | |
| 农药使用,%(n个) | | | | |
| 否 | 32(143) | 29 (58) | 1.00(参考) | — |
| 用于园艺 | 21(93) | 22 (43) | 1.52 (0.88–2.61) | 0.13 |
| 专业 | 47 (213) | 49 (98) | 1.64 (0.94–2.84) | 0.08 |
| 用于园艺或专业 | 68 (306) | 71 (141) | 1.57 (0.98–2.53) | 0.06 |
| 未知 | (39) | (10) | | |
| 一般特征. | 控件(n个=488). | 案例(n个=209). | OR(95%置信区间)一. | P(P) 一. |
|---|
| PD发病年龄,中位数(范围) | — | 65 (35–75) | — | — |
| 研究时的年龄,中位数(范围) | 68 (36–79) | 68 (37–75) | — | 0.89 |
| 男性,%(n个) | 59 (289) | 57 (119) | — | — |
| 父母出生国,%(n个) | | | | |
| 父母均出生于法国 | 93 (443) | 92 (190) | 1.00(参考) | — |
| 至少有一位父母在另一个欧洲国家出生 | 7 (33) | 8 (16) | 1.24(0.63–2.43) | 0.54 |
| 未知 | (12) | (3) | | |
| 帕金森氏症家族史,%(n个) | | | | |
| 否 | 95 (451) | 91 (189) | 1.00(参考) | — |
| 是的 | 5(25) | 9 (18) | 1.75 (0.92–3.22) | 0.09 |
| 未知 | (12) | (2) | | |
| 吸烟,%(n个) | | | | |
| 从未 | 67 (323) | 75 (155) | 1.00(参考) | — |
| 戒烟者 | 24 (115) | 20 (42) | 0.71 (0.46–1.11) | 0.13 |
| 当前吸烟者 | 9 (45) | 5 (10) | 0.42 (0.20–0.89) | 0.02 |
| 曾经吸烟者(现任和前任吸烟者) | 33 (160) | 25 (52) | 0.63 (0.42–0.96) | 0.03 |
| ≤18包年b条 | 15 (75) | 15 (31) | 0.82 (0.49–1.38) | 0.46 |
| >18包年b条 | 18 (85) | 10 (20) | 0.42 (0.23–0.75) | 0.003c(c) |
| 未知 | (5) | (2) | | |
| 农药使用,%(n个) | | | | |
| 否 | 32 (143) | 29 (58) | 1.00(参考) | — |
| 用于园艺 | 21 (93) | 22 (43) | 1.52 (0.88–2.61) | 0.13 |
| 专业 | 47 (213) | 49 (98) | 1.64 (0.94–2.84) | 0.08 |
| 用于园艺或专业 | 68 (306) | 71 (141) | 1.57 (0.98–2.53) | 0.06 |
| 未知 | (39) | (10) | | |
| 多态性. | 控制(%). | 案例(%). | OR(95%置信区间)一. | P(P) 一. |
|---|
| iNOS 22号 | n个=482 | n个=207 | | |
| GG公司 | 36 | 34 | 1.00(参考) | — |
| 通用航空公司 | 47 | 57 | 1.21 (0.85–1.73) | 0.28 |
| AA公司 | 17 | 9 | 0.56 (0.32–1.01) | 0.05 |
| AA与 GG+GA | | | 0.50 (0.29–0.86) | 0.01 |
| 一般测试 | | | | 0.02 |
| 趋势测试 | | | | 0.27 |
| nNOS 18号机组 | n个=488 | n个=209 | | |
| 科科斯群岛 | 46 | 42 | 1.00(参考) | — |
| 计算机断层扫描 | 43 | 44 | 1.15(0.80–1.64) | 0.46 |
| TT公司 | 11 | 14 | 1.45 (0.85–2.48) | 0.17 |
| CT+TT 与CC对比 | | | 1.20(0.85–1.69) | 0.30 |
| 一般测试 | | | | 0.21 |
| 趋势测试 | | | | 0.18 |
| nNOS 29号 | n个=488 | n个=209 | | |
| 科科斯群岛 | 54 | 45 | 1.00(参考) | — |
| 计算机断层扫描 | 38 | 43 | 1.41 (0.99–2.03) | 0.06 |
| TT公司 | 8 | 12 | 2.33 (1.27–4.26) | 0.01 |
| CT+TT 与CC对比 | | | 1.53 (1.08–2.16) | 0.02 |
| 一般测试 | | | | 0.01 |
| 趋势测试 | | | | 0.004 |
| 多态性. | 控制(%). | 案例(%). | OR(95%置信区间)一. | P(P) 一. |
|---|
| iNOS 22号 | n个=482 | n个=207 | | |
| GG公司 | 36 | 34 | 1.00(参考) | — |
| 通用航空公司 | 47 | 57 | 1.21 (0.85–1.73) | 0.28 |
| AA公司 | 17 | 9 | 0.56 (0.32–1.01) | 0.05 |
| AA与 GG+GA | | | 0.50 (0.29–0.86) | 0.01 |
| 一般测试 | | | | 0.02 |
| 趋势测试 | | | | 0.27 |
| nNOS 18号机组 | n个=488 | n个=209 | | |
| 科科斯群岛 | 46 | 42 | 1.00(参考) | — |
| 计算机断层扫描 | 43 | 44 | 1.15 (0.80–1.64) | 0.46 |
| TT公司 | 11 | 14 | 1.45(0.85–2.48) | 0.17 |
| CT+TT 与CC对比 | | | 1.20 (0.85–1.69) | 0.30 |
| 一般测试 | | | | 0.21 |
| 趋势测试 | | | | 0.18 |
| nNOS 29号 | n个=488 | n个=209 | | |
| 科科斯群岛 | 54 | 45 | 1.00(参考) | — |
| 计算机断层扫描 | 38 | 43 | 1.41 (0.99–2.03) | 0.06 |
| TT公司 | 8 | 12 | 2.33 (1.27–4.26) | 0.01 |
| CT+TT 与CC对比 | | | 1.53 (1.08–2.16) | 0.02 |
| 一般测试 | | | | 0.01 |
| 趋势测试 | | | | 0.004 |
| 多态性. | 控制(%). | 案例(%). | OR(95%置信区间)一. | P(P) 一. |
|---|
| iNOS 22号 | n个=482 | n个=207 | | |
| GG公司 | 36 | 34 | 1.00(参考) | — |
| 通用航空公司 | 47 | 57 | 1.21 (0.85–1.73) | 0.28 |
| AA公司 | 17 | 9 | 0.56 (0.32–1.01) | 0.05 |
| AA与 GG+GA | | | 0.50 (0.29–0.86) | 0.01 |
| 一般测试 | | | | 0.02 |
| 趋势测试 | | | | 0.27 |
| nNOS 18号机组 | n个=488 | n个=209 | | |
| 科科斯群岛 | 46 | 42 | 1.00(参考) | — |
| 计算机断层扫描 | 43 | 44 | 1.15 (0.80–1.64) | 0.46 |
| TT公司 | 11 | 14 | 1.45 (0.85–2.48) | 0.17 |
| CT和TT 与CC对比 | | | 1.20 (0.85–1.69) | 0.30 |
| 一般测试 | | | | 0.21 |
| 趋势测试 | | | | 0.18 |
| nNOS 29号 | n个=488 | n个=209 | | |
| 科科斯群岛 | 54 | 45 | 1.00(参考) | — |
| 计算机断层扫描 | 38 | 43 | 1.41 (0.99–2.03) | 0.06 |
| TT公司 | 8 | 12 | 2.33 (1.27–4.26) | 0.01 |
| CT+TT 与CC对比 | | | 1.53 (1.08–2.16) | 0.02 |
| 一般测试 | | | | 0.01 |
| 趋势测试 | | | | 0.004 |
| 多态性. | 控制(%). | 案例(%). | OR(95%置信区间)一. | P(P) 一. |
|---|
| iNOS 22号 | n个=482 | n个=207 | | |
| GG公司 | 36 | 34 | 1.00(参考) | — |
| 通用航空公司 | 47 | 57 | 1.21 (0.85–1.73) | 0.28 |
| AA公司 | 17 | 9 | 0.56 (0.32–1.01) | 0.05 |
| AA与 GG+GA | | | 0.50 (0.29–0.86) | 0.01 |
| 一般测试 | | | | 0.02 |
| 趋势测试 | | | | 0.27 |
| nNOS 18型 | n个=488 | n个=209 | | |
| 科科斯群岛 | 46 | 42 | 1.00(参考) | — |
| 计算机断层扫描 | 43 | 44 | 1.15 (0.80–1.64) | 0.46 |
| TT公司 | 11 | 14 | 1.45 (0.85–2.48) | 0.17 |
| CT+TT 相对于CC | | | 1.20 (0.85–1.69) | 0.30 |
| 一般测试 | | | | 0.21 |
| 趋势测试 | | | | 0.18 |
| nNOS 29号 | n个=488 | n个=209 | | |
| 科科斯群岛 | 54 | 45 | 1.00(参考) | — |
| 计算机断层扫描 | 38 | 43 | 1.41 (0.99–2.03) | 0.06 |
| TT公司 | 8 | 12 | 2.33 (1.27–4.26) | 0.01 |
| CT+TT 与CC对比 | | | 1.53 (1.08–2.16) | 0.02 |
| 一般测试 | | | | 0.01 |
| 趋势测试 | | | | 0.004 |
| nNOS 18–nNOS 29单倍型(%). | 控件(n个=976条染色体). | 案例(n个=418条染色体). | 或一. | P(P) 一. |
|---|
| C-C公司 | 60 | 54 | 0.47 | 0.02 |
| C-T公司 | 7 | 10 | 1.41 | 0.10 |
| T-C公司 | 13 | 12 | 0.96 | 0.84 |
| T-T型 | 20 | 24 | 1.28 | 0.05 |
| nNOS 18–nNOS 29单倍型(%). | 控件(n个=976条染色体). | 案例(n个=418条染色体). | 或一. | P(P) 一. |
|---|
| C-C公司 | 60 | 54 | 0.47 | 0.02 |
| C-T型 | 7 | 10 | 1.41 | 0.10 |
| T-C公司 | 13 | 12 | 0.96 | 0.84 |
| T型 | 20 | 24 | 1.28 | 0.05 |
| nNOS 18–nNOS 29单倍型(%). | 控件(n个=976条染色体). | 案例(n个=418条染色体). | 或一. | P(P) 一. |
|---|
| C-C公司 | 60 | 54 | 0.47 | 0.02 |
| C-T公司 | 7 | 10 | 1.41 | 0.10 |
| T-C公司 | 13 | 12 | 0.96 | 0.84 |
| T-T型 | 20 | 24 | 1.28 | 0.05 |
| nNOS 18–nNOS 29单倍型(%). | 控件(n个=976条染色体). | 案例(n个=418条染色体). | 或一. | P(P) 一. |
|---|
| C-C公司 | 60 | 54 | 0.47 | 0.02 |
| C-T公司 | 7 | 10 | 1.41 | 0.10 |
| T-C公司 | 13 | 12 | 0.96 | 0.84 |
| T-T型 | 20 | 24 | 1.28 | 0.05 |
表4。nNOS 29和iNOS 22多态性对帕金森病风险的个体和联合影响
| 多态性. | . | 控制(%). | 案例(%). | OR(95%置信区间)一. | P(P). |
|---|
| nNOS 29型. | iNOS 22号. | n个=482. | n个=207. | . | . |
|---|
| 科科斯群岛 | GG+GA | 45 | 43 | 1.00(参考) | — |
| 科科斯群岛 | AA公司 | 9 | 3 | 0.29 (0.11–0.76) | 0.01 |
| CT和TT | GG+GA | 38 | 48 | 1.41 (0.98–2.04) | 0.07 |
| CT+TT | AA公司 | 8 | 6 | 0.97 (0.49–1.93) | 0.93 |
| | | | 互动 | 0.16 |
| 多态性. | . | 控制(%). | 案例(%). | OR(95%置信区间)一. | P(P). |
|---|
| nNOS 29号. | iNOS 22号. | n个=482. | n个=207. | . | . |
|---|
| 科科斯群岛 | GG+GA | 45 | 43 | 1.00(参考) | — |
| 科科斯群岛 | AA公司 | 9 | 3 | 0.29 (0.11–0.76) | 0.01 |
| CT+TT | GG+GA | 38 | 48 | 1.41 (0.98–2.04) | 0.07 |
| CT+TT | AA公司 | 8 | 6 | 0.97 (0.49–1.93) | 0.93 |
| | | | 互动 | 0.16 |
表4。nNOS 29和iNOS 22多态性对帕金森病风险的个体和联合影响
| 多态性. | . | 控制(%). | 案例(%). | OR(95%置信区间)一. | P(P). |
|---|
| nNOS 29号. | iNOS 22号. | n个=482. | n个=207. | . | . |
|---|
| 科科斯群岛 | GG+GA | 45 | 43 | 1.00(参考) | — |
| 科科斯群岛 | AA公司 | 9 | 3 | 0.29 (0.11–0.76) | 0.01 |
| CT+TT | GG+GA | 38 | 48 | 1.41 (0.98–2.04) | 0.07 |
| CT+TT | AA公司 | 8 | 6 | 0.97 (0.49–1.93) | 0.93 |
| | | | 互动 | 0.16 |
| 多态性. | . | 控制(%). | 案例(%). | OR(95%置信区间)一. | P(P). |
|---|
| nNOS 29号. | iNOS 22号. | n个=482. | n个=207. | . | . |
|---|
| 科科斯群岛 | GG+GA | 45 | 43 | 1.00(参考) | — |
| 科科斯群岛 | AA公司 | 9 | 3 | 0.29 (0.11–0.76) | 0.01 |
| CT+TT | GG+GA | 38 | 48 | 1.41 (0.98–2.04) | 0.07 |
| CT+TT | AA公司 | 8 | 6 | 0.97 (0.49–1.93) | 0.93 |
| | | | 互动 | 0.16 |
工具书类
1Polymeropoulos,M.H.,Lavedan,C.,Leroy,E.,Ide,S.E.,Dehejia,A.,Dutra,A.,Pike,B.,Root,H.,Rubenstein,J.,Boyer,R。等。(
1997
)帕金森氏病家族中确定的α-同核蛋白基因突变。科学类
,276
,2045
–2047. 2Leroy,E.、Boyer,R.、Auburger,G.、Leube,B.、Ulm,G.和Mezey,E.,Harta,G.,Brownstein,M.J.、Jonnalagada,S.、Chernova,T。等。(
1998
)帕金森病的泛素途径自然
,395
,451
–452. 3北田,T.,朝川,S.,服部,N.,松美,H.,山村,Y.,广岛,S(
1998
)parkin基因突变导致常染色体隐性遗传青少年帕金森综合征。自然
,392
,605
–608. 4Gasser,T.、Muller-Myhsok,B.、Wszolek,Z.K.、Oehlmann,R.、Calne,D.B.、Bonifati,V.、Bereznai,B.、Fabrizio,E.、Vieregge,P.和Horstmann,R.D(
1998
)帕金森氏病的易感性基因定位于染色体2p13。自然遗传学。
,18
,262
–265. 5Farrer,M.、Gwinn Hardy,K.、Muenter,M.、DeVrieze,F.W.、Crook,R.、Perez Tur,J.、Lincoln,S.、Maraganore,D.、Adler,C.、Newman,S。等。(
1999
)与帕金森病和姿势性震颤分离的染色体4p单倍型。嗯,分子遗传学。
,8
,81
–85. 6Valente,E.M.、Bentivoglio,A.R.、Dixon,P.H.、Ferraris,A.、Ialongo,T.、Frontali,M.、Albanese,A.和Wood,N.W(
2001
)常染色体隐性遗传早发性帕金森综合征新基因座PARK6在人类染色体1p35~p36上的定位。Am.J.Hum.遗传学。
,68
,895
–900. 7van Duijn,C.M.,Dekker,M.C.,Bonifati,V.,Galjaard,R.J.,Houwing-Disitermaat,J.J.,Snijders,P.J.,Testers,L.,Breedveld,G.J.,霍斯廷克,M.,Sandkuijl,L.A。等。(
2001
)Park7,一个常染色体隐性早发性帕金森病的新基因座,位于染色体1p36。Am.J.Hum.遗传学。
,69
,629
–634. 8Funayama,M.、Hasegawa,K.、Kowa,H.、Saito,M.,Tsuji,S.和Obata,F(
2002
)帕金森病(Parkinson’s disease,PARK8)的一个新基因座定位于染色体12p11.2-q13.1。安。神经。
,51
,296
–301. 9Li,Y.J.,Scott,W.K.,Hedges,D.J.,Zhang,F.,Gaskell,P.C.,Nance,M.A.,Watts,R.L.,Hubble,J.P.,Koller,W.C.,Pahwa,R。等。(
2002
)两种常见神经退行性疾病的发病年龄受基因控制。Am.J.Hum.遗传学。
,70
,985
–993. 10Tan,E.K.、Khajavi,M.、Thornby,J.I.、Nagamitu,S.、Jankovic,J.和Ashizawa,T(
2000
)帕金森病多态性关联研究的变异性和有效性。神经病学
,55
,533
–538. 11Law,A.、Gauthier,S.和Quirion,R(
2001
)对阿尔茨海默病说“不”:一氧化氮与阿尔茨海默氏型痴呆症之间的假定联系。大脑研究评论。
,35
,73
–96. 12Togashi,H.、Sasaki,M.、Frohman,E.、Taira,E.、Ratan,R.R.、Dawson,T.M.和Dawson(V.L.)(
1997
)神经(I型)一氧化氮合酶调节核因子κB活性和免疫(II型)一氧化氮合成酶的表达。程序。美国国家科学院。科学。美国
,94
,2676
–2680. 13W.K.阿尔德顿、C.E.库珀和R.G.诺尔斯(
2001
)一氧化氮合酶:结构、功能和抑制。生物化学。J。
,357
,593
–615. 14Klivenyi,P.,Andreassen,O.A.,Ferrante,R.J.,Lancelot,E.,Reif,D.和Beal,M.F(
2000
)抑制神经元型一氧化氮合酶可防止MPTP毒性。神经报告
,11
,1265
–1268. 15Hantraye P.、Brouillet E.、Ferrante R.、Palfi S.、Dolan R.、Matthews R.T.和Beal M.F(
1996
)抑制神经元一氧化氮合酶可预防MPTP诱导的狒狒帕金森综合征。自然医学。
,2
,1017
–1021. 16Przedborski,S.、Jackson-Lewis,V.、Yokoyama,R.、Shibata,T.、Dawson,V.L.和Dawson(T.M.)(
1996
)神经元一氧化氮在1-甲基-4-苯基-1,2,3,6-四氢吡啶(MPTP)诱导的多巴胺能神经毒性中的作用。程序。国家。阿卡德。科学。美国
,93
,4565
–4571. 17Schulz,J.B.、Matthews,R.T.、Muqit,M.M.、Browne,S.E.和Beal,M.F(
1995
)7-硝基吲唑对神经元一氧化氮合酶的抑制可保护小鼠免受MPTP诱导的神经毒性。神经化学杂志。
,64
,936
–939. 18利伯拉托雷,G.T.,杰克逊·莱维斯,V.,武科萨维奇,S.,曼迪尔,A.S.,维拉,M.,麦考利夫,W.G.,道森,V.L.,道生,T.M.和普雷德博斯基,S(
1999
)诱导型一氧化氮合酶刺激帕金森病MPTP模型中的多巴胺能神经变性。自然医学。
,5
,1403
–1409. 19Dehmer,T.、Lindenau,J.、Haid,S.、Dichgans,J.和Schulz,J.B(
2000
)诱导型一氧化氮合酶缺乏可防止MPTP毒性体内.神经化学杂志。
,74
,2213
–2216. 20H.Checkoway和L.M.Nelson(
1999
)帕金森病病因学研究的流行病学方法。流行病学
,10
,327
–336. 21Grunewald,T.和Beal,M.F(
1999
)NOS敲除与神经保护。自然医学。
,5
,1354
–1355. 22波根·S、德米尔戈伦·S、塔斯克兰·D、卡尼特·L、伊尔马兹·O、科伊鲁·E·O、巴尔干·B和伦敦·E·D(
2000
)尼古丁调节大鼠大脑中的一氧化氮。欧洲神经心理药理学。
,10
,463
–472. 23Matthews,R.T.、Yang,L.和Beal,M.F(
1997
)S-甲基硫代瓜氨酸是一种神经型一氧化氮合酶抑制剂,可防止丙二酸盐和MPTP的神经毒性。实验神经学。
,143
,282
–286. 24伊玛目S.Z.、伊斯兰F.、伊扎克Y.、斯利克W.Jr.和阿里S.F(
2000
)通过靶向一氧化氮和过氧亚硝酸盐预防多巴胺能神经毒性:对预防甲基苯丙胺诱导的神经毒性损伤的影响。纽约学院安。科学。
,914
,157
–171. 25库雷希,G.A.、贝格,S.、贝德纳尔,I.、索德斯滕,P.、福斯伯格,G.和塞登,A(
1995
)帕金森病患者脑脊液中亚硝酸盐浓度升高。神经报告
,6
,1642
–1644. 26Gatto,E.M.、Riobo,N.A.、Carreras,M.C.、Chernavsky,A.、Rubio,A.、Satz,M.L.和Poderoso,J.J(
2000
)帕金森病中性粒细胞神经元一氧化氮合酶的过度表达。一氧化氮
,4
,534
–539. 27谢吉尔·J.K.、卡马克·R.、库珀·C.E.、库珀·J.M.、曼·V.M.和夏皮拉·A.H(
1996
)检测人类黑质中与帕金森病相关的亚硝基化合物。生物化学。生物物理学。Res.Commun公司。
,228
,298
–305。 28Hunot,S.、Boissiere,F.、Faucheux,B.、Brugg,B.、Mouatt-Prigent,A.、Agid,Y.和Hirsch,E.C(
1996
)一氧化氮合酶与帕金森病的神经元易损性。神经科学
,72
,355
–363。 29Good,P.F.,Hsu,A.,Werner,P.,Perl,D.P.和Olanow,C.W(
1998
)帕金森病中的蛋白质硝化。神经病理学杂志。实验神经学。
,57
,338
–342。 30Renodon,A.,Boucher,J.L.,Sari,M.A.,Delaforge,M.,Ouazzani,J.和Mansuy,D(
1997
)溴隐亭是脑一氧化氮合酶的一种强抑制剂:其治疗作用的可能后果。FEBS信函。
,406
,33
–36. 31Wang,X.L.,Sim,A.S.,Wang,M.X.,Murrell,G.A.,Trudinger,B.和Wang,J(
2000
)基因型依赖性和香烟特异性对内皮型一氧化氮合酶基因表达和酶活性的影响。FEBS信函。
,471
,45
–50. 32Weruaga,E.,Balkan,B.,Koylu,E.O.,Pogun,S.和Alonso,J.R(
2002
)长期服用尼古丁对大鼠脑内一氧化氮合酶表达和活性的影响。《神经科学杂志》。物件。
,67
,689
–697. 33Deliconstantinos,G.和Villiotou,V(
2000
)香烟烟雾的气相氧化剂增加了兔脑突触体的一氧化氮合酶和黄嘌呤氧化酶活性。神经化学。物件。
,25
,769
–774. 34Xu,W.、Liu,L.、Emson,P.、Harrington,C.R.、McKeith,I.G.、Perry,R.H.、Morris,C.M.和Charles,I.G(
2000
)NOS2A基因中的CCTTT多态性与路易体痴呆相关。神经报告
,11
,297
–299. 35Martin,E.R.,Scott,W.K.,Nance,M.A.,Watts,R.L.,Hubble,J.P.,Koller,W.C.,Lyons,K.,Pahwa,R.,Stern,M.B.,Colcher,A。等。(
2001
)tau基因单核苷酸多态性与晚发性帕金森病的相关性。日本汽车制造商协会
,286
,2245
–2250. 36Farrer,M.、Skipper,L.、Berg,M.和Bisceglio,G.、Hanson,M.,Hardy,J.、Adam,A.、Gwinn-Hardy,K.和Aasly,J(
2002
)在挪威人群中,tau H1单倍型与帕金森病相关。神经科学。莱特。
,322
,83
–86. 37Maraganore,D.M.、Hernandez,D.G.、Singleton,A.B.、Farrer,M.J.、McDonnell,S.K.、Hutton,M.L.、Hardy,J.A.和Rocca,W.A(
2001
)帕金森病扩展tau基因单倍型的病例对照研究。安。神经。
,50
,658
–661. 38Scott,W.K.,Nance,M.A.,Watts,R.L.,Hubble,J.P.,Koller,W.C.,Lyons,K.,Pahwa,R.,Stern,M.B.,Colcher,A.,Hiner,B.C。等。(
2001
)帕金森病的全基因组筛查:多基因证据。日本汽车制造商协会
,286
,2239
–2244. 39Lo,H.S.、Hogan,E.L.和Soong,B.W(
2002
)台湾帕金森病患者神经元型一氧化氮合酶基因5′-侧翼区多态性。神经学杂志。科学。
,194
,11
–13。 40Grasemann,H.、Yandava,C.N.和Drazen,J.M(
1999
)神经一氧化氮合酶(NOS1)是哮喘的主要候选基因。临床。实验性过敏
,29(补充4)
,39
–41. 41Elbaz,A.,Levecque,C.,Clavel,J.,Vidal,J.S.,Richard,F.,Corrèze,J.R.,Delemotte,B.,Amouyel,P.,Alpérovitch,A.,Chartier-Harlin,M.C.和Tzourio,C(
2002
)UCH-L1基因S18Y多态性与帕金森病:年龄依赖关系的证据。莫夫。迪索德。
,内政部10.1002/mds.10326。 42M.C.de Rijk、W.A.Rocca、D.W.Anderson、M.O.Melcon、M.M.Breteler和D.M.Maraganore(
1997
)帕金森病诊断标准的人群视角。神经病学
,48
,1277
–1281. 43Bower,J.H.、Maraganore,D.M.、McDonnell,S.K.和Rocca,W.A(
1999
)1976–1990年明尼苏达州奥姆斯特德县帕金森综合征的发病率和分布。神经病学
,52
,1214
–1220. 44Clavel,J.、Glass,D.C.、Cordier,S.和Hemon,D(
1993
)专家对基于人群的病例对照研究中有机溶剂职业接触回顾性评估的标准化。国际流行病学杂志。
,22(补充2)
,第121节
–126. 45W.F.斯图尔特和P.A.斯图尔特(
1994
)职业病例对照研究:I.收集工作史和工作相关暴露的信息。美国医学杂志。
,26
,297
–312. 46D.J.沙伊德、C.M.罗兰、D.E.蒂内斯、R.M.雅各布森和G.A.波兰(
2002
)连锁阶段不明确时,性状和单倍型之间关联的得分测试。Am.J.Hum.遗传学。
,70
,425
–434。 47Thompson,E.A.、Deeb,S.、Walker,D.和Motulsky,A.G(
1988
)紧密连锁标记之间连锁不平衡的检测:AI-CIII载脂蛋白基因的RFLP。Am.J.Hum.遗传学。
,42
,113
–124.


{kind=link}



