摘要
1引言
在形态发生过程中,组织生长并自组织成复杂的功能单位,如器官。该过程通过信号和机械相互作用进行严格控制。组织中的长程信号相互作用可以由称为形态原的扩散物质和长程细胞过程介导(Restrepo公司等。, 2014). 扩散因子的动力学通常可以用连续反应-对流-扩散偏微分方程(PDEs)系统很好地描述。适当的组织表现取决于相关的时间尺度。对于均质各向同性胚胎组织,实验表明,该组织在长时间尺度上很好地近似于粘性流体(30分钟到数小时后平衡),在短时间尺度上(秒到分钟)近似于弹性材料(福加斯等。, 1998). 然而,生物控制通常发生在较短的时间范围内,许多细胞过程,如细胞迁移和粘附、细胞极性、定向分裂、单层结构和分化,不能以直接的方式形成连续的配方。为了研究这些过程,已经开发了许多不同规模和不同详细程度的基于细胞的模拟技术;在这里,我们讨论每个类别的主要代表。
这个Cellular Potts模型,由介绍格拉纳和格拉泽(1992),在一个格子上求解,每个格子点都有一个表示细胞单位的广义自旋值。与伊辛模型类似,哈密顿能量函数也使用Metropolis算法进行了公式化和最小化。它已经应用于许多问题,并在软件中实现CompuCell3D(重拍等。, 2012). 然而,生物问题与哈密顿量、温度和时间步长之间的对应并不总是简单的。
这个亚细胞元素模型将细胞划分为亚细胞元素,亚细胞元素由计算粒子表示。这些元素通过相互作用的势相互作用,这些势需要建模。元件的运动受过阻尼Langevin动力学控制,因此该方法是无网格的。该框架最初由引入纽曼(2005)后来由申请桑德修斯等。(2011年a),b条). 这种方法允许在2D和3D中进行详细的生物物理建模。
这个球体模型由开发德拉兹多等。(2007)假设非结构细胞群中的细胞类似于胶体颗粒。细胞被建模为点粒子,具有相互作用势。它们的运动包括随机运动和定向运动。相邻细胞形成粘着键,使用从接触力学借用的模型表示,例如约翰逊-肯德尔-罗伯茨模型(楚等。, 2005). 许多细胞过程,如细胞形状改变、分裂、死亡、溶解、细胞与细胞的相互作用和迁移,都已成功地转化为球体模型(德拉兹多等。, 2007). 尚未引入和实施细胞内外扩散。球体模型有效地扩展到3D,并已在开源框架中实现CellSys公司(Hoehme和Drasdo,2010年).
这个顶点模型使用多边形(或3D中的多面体)表示密集组织中的细胞,例如果蝇属翼盘上皮(法哈迪法等人。, 2007). 对于每个顶点,力要么通过势计算,要么直接计算。随后根据过阻尼运动方程或通过蒙特卡罗算法移动顶点。该模型在开源软件中实现贞洁(皮特·法西斯等。, 2009).
这个粘弹性胞模型(也称为IBCell模型)雷尼亚克等。(2004)和雷尼亚克(2007)使用浸没边界(IB)方法(Peskin,2003年)将单个可变形细胞表示为浸没的弹性体。细胞质、细胞外基质和液体由粘性的不可压缩流体表示。在这个框架中,实现了细胞生长、细胞分裂、凋亡和极化等大量生物过程。该模型用于研究肿瘤和上皮动力学。由于细节层次较高,粘弹性单元模型的计算成本较高,尚未在3D中实现。
软件框架虚拟叶已在默克斯等。(2011)虽然单元表示类似于顶点单元模型,但动力学是通过使用蒙特卡罗算法最小化哈密顿量来实现的。该模型假设细胞壁是刚性的,这对于植物的形态发生是合适的。
对于许多由生物分子信号和组织物理之间紧密相互作用产生的形态发生现象,需要明确的细胞形状计算表示。在这里,我们提出了一个基于IBCell模型的灵活软件框架,作为一种新颖技术,它允许将生物分子信号模型紧密耦合到细胞解析的物理组织模型。第二节描述了该模型的核心组件和一般方法。第三部分详细介绍了该软件及其主要功能。第四章给出了应用示例,以演示框架的功能。
2进近
我们的方法允许对组织和信号动力学进行耦合模拟。为了描述组织动力学,粘弹性细胞模型需要表示细胞结构及其弹性特性,以及细胞质和细胞周围细胞外空间的粘性行为。因此,该模型基于三个核心部分:细胞的表示、流体和流体-结构相互作用的表示以及组织部分与信号模型的耦合。为了描述粘性流体和浸没在流体中的弹性结构之间的相互作用,我们使用浸没边界法(Peskin,2003年)如之前在粘弹性细胞模型中实现的,也称为IBcell模型(雷尼亚克等人。, 2004;Rejniak,2007年). 为了解决粘性流体的行为,我们使用了格子Boltzmann方法,这是一种有效的介观数值格式,最初是为了解决流体动力学问题而开发的(Chen和Doolen,1998年). 该方法以前已成功应用于反应扩散方程,如图灵系统(蓬斯·道森等。, 1993)以及耦合标量场,如温度(郭等。, 2002). 该方法首次与浸没边界法相结合(Peskin,2003年)由Feng和Michaelides(2004)后来被用于研究流动中的红细胞张等。(2007)在下文中,我们概述了实现的方法;第3节给出了实现细节。
2.1单元表示
细胞被表示为无质量的纯弹性结构,由形成多边形的几何点集来描述。几何点通过力连接。在一级近似中,可以确定弹性结构来代表弹性细胞膜。然而,可以在细胞内和细胞外体积中添加更多弹性结构,以模拟细胞骨架或细胞外基质的粘弹性特性。用户可以实现作用于细胞表征的生物机制。例如,当两个相邻单元边界之间的距离低于阈值距离时,可能会创建到相邻单元的新连接。同样,过度拉伸时可能会删除连接。
2.2流体与流体结构相互作用
粘弹性细胞模型将细胞(细胞质)以及细胞外空间(间质流体和细胞外基质)的含量表示为粘性牛顿流体。内部和外部流体与弹性膜相互作用,即流体对膜施加力,而膜对流体施加力。此外,由力引起的流体速度场会使弹性结构发生移动和变形。这种被称为流体-结构相互作用的相互作用是组织模型的核心。作用在这些点上的力(例如膜张力或细胞-细胞力)通过将力分布到周围流体而施加在流体上。由于局部压力,流体移动。在此步骤中,膜点被流体被动平流。因此,需要重新评估各点的部队。通过重复强制平流步骤,交互作用以迭代方式实现。
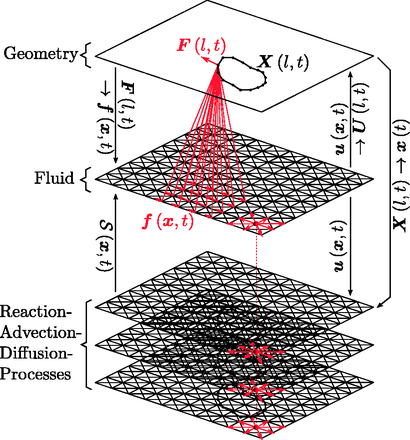
由于这个迭代过程,(弹性)结构与(粘性)流体耦合。根据参数化,该模型可以描述弹性、粘性或粘弹性材料行为。的上部图1说明了浸没边界交互作用。实现的浸入式边界核函数具有有界支持,即每个几何点影响并且仅受其直接邻域的影响。这里,内核函数的维度是四乘四(参见。图1). 流体方程用格子Boltzmann方法求解(Chen和Doolen,1998年),详见补充材料(第6节)。雷诺数通常为;因此,该状态由斯托克斯流描述(雷诺数为,使用U型作为特征速度,L(左)特征长度刻度和ν运动粘度。假设和,然后可以估计(福加斯等。, 1998)).
图1。
算法概述。该算法由三个耦合层组成。几何图形(顶部,在图2)用于计算力作用于每个几何体节点。这些不一定与点阵点重合的力使用IB方法核函数分散到流体点阵(中间部分),将流体解算器向前推进一个时间步长后,使用相同的核函数将速度插值到几何节点位置,。几何体节点根据其速度移动,并重新启动迭代。速度流体晶格的位置也被复制到反应-对流-扩散解算器(PDE)中几何图形的。用于建模信号的反应-平流-扩散解算器的状态可用于计算质量源对于流体解算器
2.3信号
信号网络表示为反应-对流-扩散过程系统。弹性膜可分别作为仅存在于胞外或胞内体积的化合物的无流边界。反应-对流-扩散求解器可以配备潜在的耦合反应项,以模拟扩散因子的信号传递交互作用。根据模型,信号可能会影响组织动力学。例如,可以通过使流体的质量源依赖于反应-对流-扩散解算器的值来实现,从而使组织局部膨胀(参见。图1). 此外,扩散化合物可以单独配置为在整个区域内自由扩散,或仅在细胞内外扩散(例如,使用细胞膜的无通量边界条件)。
3软件
3.1单元表示
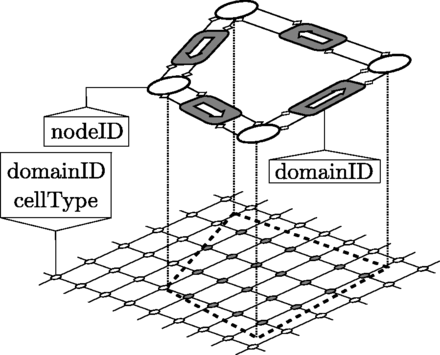
单元几何结构由两个元素组成几何节点作为IB点,以及连接,连接成对几何节点。简化的单元格显示在图2. The连接被归因于域ID标志,它是包围域的标识符(遵循逆时针方向性约定)。另一侧(右侧)的域标识符按照惯例为零,表示间隙空间。这个域ID的连接被复制到流体和反应-平流-扩散解算器。此外域ID与单元格类型标志关联,单元格类型。通过应用自定义差异规则单元格类型可以根据自定义标准更改单个单元格的;否则,所有单元格默认为单元格类型 = 1(带有单元格类型 = 0再次成为间隙空间)。这样,反应项和质量源可能取决于特定的细胞或特定的细胞类型。
图2。
几何图形表示的元素。单元是闭合多边形,由几何体节点(在顶部讨论)和每个两个几何体节点之间的连接(顶部的阴影框)组成。每个连接存储对其前面和后续几何体节点的两个引用,以及反之亦然每个几何体节点存储对其之前和后续连接的两个引用(由顶部的聚合箭头显示)。按照惯例,多边形的方向是逆时针的。每个几何体节点都有一个唯一的、不可变的nodeID属性,该属性在创建新几何体节点时在内部分配。每个连接都有一个domainID属性,该属性在左侧表示域的域标识符。右侧的域标识符定义为零,代表细胞外空间。使用连接的domainID,将自动设置晶格节点的domainID(下部)。此外,每个domainID都与一个cellType关联。用户可以根据domainID和/或cellType属性设置MassSolverXX、BioSolverXX和CDESolverXX的行为
3.2用户提供的解算器
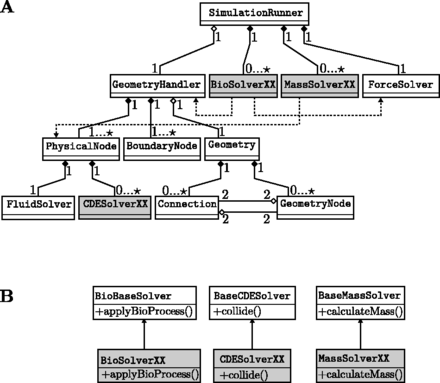
用户可以添加以下例程:MassSolverXX、CDESolverXX和生物解决方案XX(XX是一个待选的名字)。这个质量求解器XX-如前所述,从流体解算器添加或减去质量。这个CDE解决方案XX用于实现信号模型的反应项。最后生物解决方案XX可用于对几何体和力执行生物激励操作。这种操作可能是细胞分裂,详见第3.5.4节。图3A总结了最重要的类及其交互。需要自定义的类被着色。为了添加新的自定义例程(例如,质量修改解算器MassSolverXX,反应-对流-扩散求解器CDE解决方案XX或是一个有生物动机的解决者生物溶剂XX),用户需要从他们各自的虚拟基类继承(参见。图3B) ●●●●。图4可视化例程,这些例程由SimulationRunner仿真跑步者(参见。图3A) ●●●●。
图3。
重要类的简化UML图。必须由用户提供的类被着色。XX表示任意解算器名称。(A)这个SimulationRunner仿真跑步者控制模拟的执行。这个GeometryHandler公司有一个集合物理节点,表示晶格,一组边界节点它们被编织在格子里几何图形对象。后者包含单元格的几何信息,即几何节点和连接。这个几何节点和连接每一个都有前面和后面元素的两个引用,如中所述图2.生物解决方案XX从GeometryHandler公司和ForceSolver来相应地更改状态。类似地MassSolverXX公司获得晶格的参考并向流体添加质量源。(B)要实现新的自定义例程,用户必须从提供的基类(从生物基础解算器对于出于生物学动机的日常活动BaseCDE溶剂对于反应-对流-扩散过程基本质量求解器用于体量修改例程)
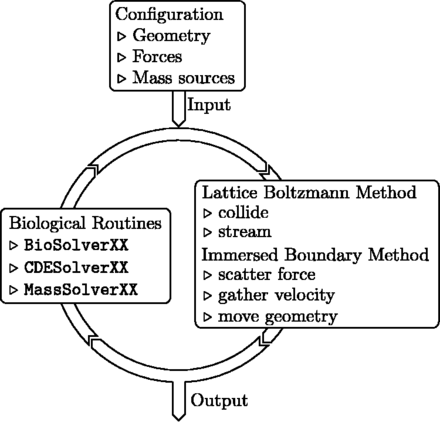
图4。
求解器中的迭代处理。启动时,库加载用户提供的配置文件(包含全局仿真参数、初始几何体、初始力)。在每次迭代期间,库的类SimulationRunner仿真跑步者(参见。图3A) 先后调用物理例程(用于解决流体和反应-平流-扩散过程的格子-玻尔兹曼方法,以及用于解决流体-结构相互作用的浸没边界方法)和生物例程(几何的生物动机重新排列、力的修改等)。当前配置和可选的整个解算器状态可以以选定的频率保存
3.3输入和输出
通过加载和转储配置文件来实现与用户的通信。通用配置文件包含全局模拟参数,例如反应-对流-扩散解算器的模拟时间、域大小、流体粘度和扩散系数。几何点和相应的几何连接存储在几何文件中。第三个文件包含力,包括一对几何点之间的力、自由定义的力或空间锚定点。流体和反应对流扩散解算器状态可以写入.txt文件或.vtk格式,并可以使用第三方软件(例如Matlab或ParaView)进行后处理。(可选)可以以可加载格式保存解算器状态以恢复模拟。
3.4物理过程
3.4.1粘性和弹性行为
粘性行为是使用不可压缩流体的表示来实现的(使用格子Boltzmann方法求解,参见补充材料),该表示在流体动力极限下收敛到Navier–Stokes方程。流体在规则笛卡尔和欧拉网格上求解。膜由一组点表示,这些点连接形成闭合多边形。各种力可能作用于膜节点,例如膜张力(参见第3.4.3节)。流体和弹性结构之间的相互作用采用浸没边界法(参见补充材料)。膜点根据局部流体速度场以拉格朗日方式移动。
3.4.2生化化合物的反应扩散
生化信号可以用一组耦合的反应扩散PDE来描述。与流体方程类似,这些方程在规则笛卡尔和欧拉网格上求解(使用晶格-玻尔兹曼方法求解,参见补充材料)。化合物的浓度可由其他溶剂获得,例如使其他过程(如细胞分裂)依赖于信号因子。细胞边界可以选择为扩散化合物看不到的边界,也可以选择为无通量边界。为了解释平流,流体速度场直接从流体解算器传递,因为流体和反应扩散晶格在空间上重合。求解器的耦合在中可视化图1.
3.4.3作用力
力是模拟环境中不可或缺的一部分。力总是与膜点相连。任何类型的保守力(可以从势导出)都可以很容易地实现。目前,实施了以下类型的部队:
两个几何节点之间的弹簧力
几何节点和空间锚点之间的弹簧力
作用在几何节点上的自由力
水平或垂直滑动力(因此仅强制年或x个坐标)
两个几何节点之间的恒力
应用示例包括两个几何节点之间的恒定力,该力可用于模拟恒定膜张力,从而使细胞周长最小化(见第3.5.2节)。此外,几何点可以动态地探索其局部邻域,并向另一个几何点施加力来自另一个单元格因此,模拟细胞-细胞连接(在第3.5.3节中讨论)。
3.5生物过程
生物溶剂(BioSolver)适应与生物过程相关的功能。这些过程可能主要与力和几何形状的修改有关。BioSolver可以完全访问化合物浓度。此外,它还知道这些单元,它们的几何图形是单独存储的。这使BioSolver能够计算细胞面积和平均或综合化合物浓度。因为所有细胞都是单独标记的,所以细胞行为可以依赖于细胞身份。除了单元标识之外,单元还带有单元类型标记,可以根据运行时条件进行更改。后一种功能可用于模拟细胞分化。
以细胞分裂事件为例。这里,必须选择一个分割平面。其位置和方向的选择取决于用户的模型:细胞分裂平面可以设置为垂直于细胞伸长最大的轴。接下来,必须对单元格进行分割,这需要删除相应的几何连接,并插入新的几何节点和连接以闭合分割的单元格。
注意,化合物的浓度场以及流体解算器的速度场和压力场在生物模块中没有直接改变。
3.5.1电池面积控制
根据用户的生物模型,必须控制细胞面积。通过假设一个单元可能会改变其在三维空间的范围,该区域可能会因其相邻单元施加的力而收缩或膨胀,这可以有效地建模为“区域弹性”。在极限情况下,电池抵抗外力,保持其面积,仅对静水压力的变化作出反应。通常,为了控制单元格的面积,需要调整每个单元格的参考面积。参考面积用作简单比例控制器的设定点,即局部质量源在牢房里k个与当前单元格区域之间的面积差成正比和设定值区域:哪里α是一个比例常数。可以容易地实现更先进的控制方法,例如比例积分控制方法。 这种控制细胞面积的方法也可用于使细胞以受控的方式生长或收缩,即分化为肥大细胞类型的细胞可能会体积增大。实施这一过程只需将新的目标区域设置为设定点区域即可。区域控制器将使单元格靠近其新区域。
3.5.2膜张力
作用于两对膜点之间的力的定义允许模拟细胞的膜张力。默认情况下,恒定的收缩力具有量级应用于每对相邻的膜点。因此,膜点上的合力我由指向其前膜点的力组成我−1,以及指向其连续膜点的力我+1: 这种方法可以被解释为一种主动重塑的膜:当拉伸时,为了不增加较长时间尺度(小时)的膜张力,合成新膜。另一方面,过多的膜被降解,以承受膜张力。因此,膜张力使细胞的周长最小化。由于细胞内液体(以及细胞区域)在没有相邻细胞和活性机制的情况下保持不变(参见第3.5.1节),细胞呈圆形。在短时间尺度(秒)上,可以使用胡克弹簧势对被动(非重塑)弹性膜进行建模。然后,膜张力与静止膜周长的偏差成正比。在这两种情况下,薄膜都是柔性的(即没有弯曲刚度);如果用户需要弯曲刚度,可以在自定义BioSolver中轻松实现。
膜张力的实施需要考虑几何重网格。每当插入一个新的膜点时,它都需要立即与相邻的膜点相连,因为在没有膜张力的情况下,细胞会过度拉伸。膜点的力需要在移除时移除。从算法上讲,这是通过在每个时间步长去除和重建所有膜力来实现的。此时,膜张力的大小取决于信号因素。
生物溶剂膜张力是一个班级通过立即重塑来管理膜张力的例子,以及BioSolverHookean膜张力实现了简单的胡克弹簧。
3.5.3细胞连接
细胞可以与相邻细胞建立细胞连接。在最简单的情况下,每个膜点我使用函数获取半径最小且避免距离最近的几何节点得到最近的膜点j个在预定义的截止半径内的另一个单元格我最大值如果没有这样的膜点,则为零。一旦候选膜点满足标准,一个新的虎克力具有弹簧常数和静止长度我0创建时间: 细胞连接力定期(可能不是在每个时间点)删除和更新,其中细胞连接更新的频率可能反映细胞连接合成速率。
功能获取半径为且避免的几何节点返回另一个细胞的所有膜点,这些点位于预定义的截止半径内;返回的列表可能为空。这为通过从候选列表中随机选择膜点引入随机性提供了可能性。形成交叉的概率可能取决于交叉长度:越短,形成新交叉的概率越高。此外,膜点的移除可能是随机的,并且取决于连接长度的概率,即过度拉伸的连接被移除的概率更高。甚至连连接处需要更新的膜点也可以随机选择。同样,每次更新的膜节点数反映了细胞有限的细胞连接合成活动。
膜点内部存储在快速邻域列表数据结构中,非常适合空间范围查询。BioSolverCellJunction公司是负责细胞连接的类的示例。
3.5.4细胞分裂
细胞分裂功能需要几个步骤。首先,必须确定划分哪些单元的标准。标准可能是最大细胞面积、最大空间扩张或生化信号。一旦一个单元格提交进行分割,就必须选择单元格分割平面。同样,如何选择飞机取决于生物模型。一个常用的规则是使用由随机方向向量和单元重心定义的平面。然而,可以很容易地实施不同的规则,例如从非均匀概率分布中提取的随机方向(反过来,可以通过信令因子梯度等进行控制)或垂直于最长轴的分割平面(Minc公司等。, 2011). 在下一步中,确定与分割平面相交的两个膜段;这在中实现获取两个连接随机方向或获取两个连接最长轴。这两个膜段随后被移除,穿过细胞的两个新膜段被引入,导致母体细胞被切割。最后,必须为其中一个子单元指定一个新的域标识号;另一个子单元从母单元继承域标识号。新的域标识号设置为最大域标识号加一,并自动复制到物理网格。默认情况下,两个子单元都从母单元继承单元类型标志,该标志也自动复制到了物理网格。
基本的单元划分功能在类中实现BioSolverCellDivision公司。
3.5.5区别
分化根据用户定义的、生物激发的规则更改细胞的细胞类型标志。这些规则可能基于细胞面积,或者基于信号因子浓度,可能整合在细胞面积上。一旦被承诺分化,细胞就会根据规则改变其细胞类型标志。新的细胞类型标志将自动复制到物理网格。细胞类型标志可用于进行信号动力学,也可用于其他依赖于细胞类型的生物激发过程。
域标识符标志和单元类型标志之间的关联存储在单元格类型跟踪器映射_,它是GeometryHandler公司。这确保了生物解决方案XX类可以轻松访问此信息。微分控制的基本实现可以在中找到生物解决方案差异化.
3.6准确度和性能
格子Boltzmann格式具有二阶精度,显式浸没边界法在空间和时间上具有一阶精度。内部数据结构使用快速邻居列表(单元格列表)实现来优化范围查询(例如搜索本地邻居中的其他单元格),其搜索复杂性为,使用N个代表细胞的膜点数。许多迭代计算(Lattice Boltzmann和浸没边界例程,如粒子流和碰撞、速度聚集和力散射)使用共享内存范式进行并行化。然而,一些计算步骤无法并行化。这通常是在共享数据结构上发生写操作的情况,例如存储几何体节点和力结构的数据结构(例如ForceSolver::deleteForceType()和GeometryHandler::computeAreas())。此外,几何重网格(细化和粗化)函数以及数据I/O没有并行化,但假设发生的频率远低于实际的流体和反应-对流-扩散解算器。因此,由于序列代码的比例不容忽视,因此软件最好在快速多核处理器上运行。
3.7工具、依赖性和文档
需要一个支持C++0x的编译器(如GCC 4.7或更高版本)。该软件依赖于Boost(http://www.boost.org;1.54.0或更高版本)、OpenMP、CMake(http://www.cmake.org)和vtk(网址:http://www.vtk.org/;5.8或更高)。源代码使用Doxygen进行了大量文档记录(http://www.stack.nl网站/dimitri/doxygen)。吉特(网址:http://git-scm.com)用于版本控制。该软件仅在linux操作系统上进行了测试。
3.8可用性
文档和源代码可在http://tanakas.bitbucket.org/lbibcell/index.html.
4应用示例
4.1细胞分裂、分化和信号传递
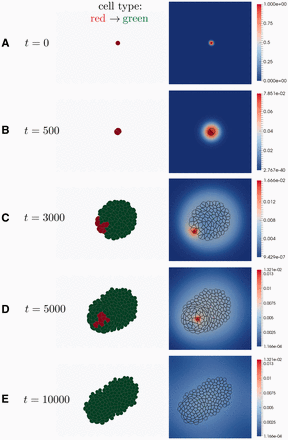
为了演示该软件的功能,我们首先考虑一个具有细胞类型特异性细胞分裂和信号依赖性分化的组织模型(图5). 开始时,半径为R(右) = 10被放置在一个二次400x400域的中间(图5A) ●●●●。等压边界条件设置在域边界。最初的细胞是红细胞型,正在高速增殖。当考虑单层上皮时,假设细胞生长和最终增殖建模所需的大量摄取是从顶腔通过顶膜发生的。此外,最初的细胞分泌一种信号因子抑制红细胞向绿细胞分化。一旦单元格面积增加一倍,单元格将按随机方向划分(参见。图5B) ●●●●。子细胞继承细胞类型,但只有母细胞继续表达信号分子.所有红色细胞都整合了覆盖他们的区域。对于低信号水平,红细胞类型分为绿细胞类型。绿色细胞类型不会生长,只有在外力拉伸细胞时才会分裂。在图5C、 细胞分裂后,子细胞的信号水平下降,发生分化。经过几轮细胞分裂后,组织开始形成(参见。图5D) ●●●●。靠近分泌的初始细胞的细胞仍然不受分化的保护,而距离较远的细胞则不可逆地分化。由于随机选择的细胞分裂轴,可能会发生增殖的红细胞被捕获的情况(参见。图5E) ●●●●。的表达式在时间关闭t吨 = 5000,因此很快就会导致完全分化(参见。图5F) ●●●●。增殖停止后,细胞会缓慢重排,因为如果过度拉伸,细胞-细胞连接就会断裂,并形成新的连接[根据方程式(3)]. 在组织的边界,细胞试图达到球形,而在中间,主要是特征性的五角形和六角形出现(参见。图5F和补充文件S6.4)。
图5。
细胞分裂、分化和信号传递。(A类)最初的结构由单个圆形红细胞组成。红细胞类型以高速增殖。最初的细胞被标记并表达一个信号分子抑制分化。(B)第一次细胞分裂发生。分割轴是随机选择的。子细胞继承母细胞的细胞类型,但只有母细胞继续表达信号分子. (C)在远离原始细胞的细胞中,信号水平(信号分子的空间整合浓度)下降,并发生向绿色细胞类型的分化。绿色细胞类型不会在内部生长,只有在受到外力过度拉伸时才会分裂。(D)由于随机选择的细胞分裂轴,高度增殖的红细胞被困在形成组织中。在t吨 = 5000,分化抑制分子的表达被关闭,导致剩余红细胞分化。(E)在没有高增殖的情况下,细胞会重新排列以最大化周长/面积比。出现了特征性的五角和六角细胞形状(参见补充文件S6.4)。靠近边界的单元格尝试形成圆形
4.2生长细胞域上的图灵模式
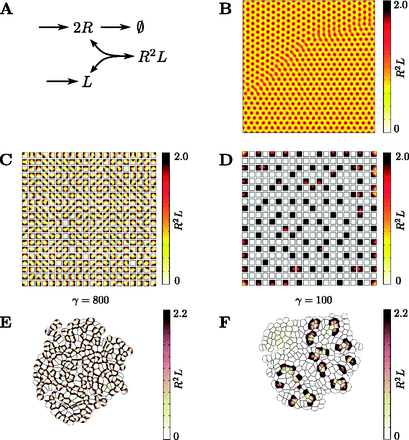
图6。
生长细胞域上的图灵模式。(A)图灵不稳定性可以通过Schnakenberg型反应实现,涉及缓慢扩散的化合物R(右),此处解释为受体和快速扩散化合物L(左),此处解释为自由扩散配体。一个配体分子与两个受体结合,形成复合物该复合物可以被解释为生物信号。(B)该模型在连续正方形晶格上求解(使用d日 = 1, γ = 800,一 = 0.1,b条 = 0.9),从而形成经典的常规斑点图案。生物信号如图所示。(C)与B中相同的系统在理想的静态细胞域上求解,即受体的扩散R(右)仅限于单元格。新兴生物信号现在分布不规则。(D)与C中相同的系统,但带有γ = 100,在理想的静态细胞域上求解。较少的细胞显示显著的信号水平没有发现有规律的图案(盐和胡椒图案)。(E类)与C中相同的系统在不断增长的细胞域中求解。细胞的增殖速度与其信号成正比产生的模式在更大范围内具有规律性,但局部模式与连续模式的行为有显著差异(B类)和静态蜂窝(C类)域。(F类)与D中相同的系统在不断增长的细胞域上求解。细胞的增殖率与信号的局部强度成正比.高水平的活性细胞簇浮现
根据域的类型,我们观察到不同的模式。在连续域上,我们得到了众所周知的正则点模式(图6B) ●●●●。在理想的静态细胞域上,具有不规则内部结构的整体规则图案(图6C) 可以观察到。减小模拟参数γ(它反向控制斑点之间的距离)会导致更意外的图案:对于γ = 100,局部规律性完全丧失(图6D) ●●●●。最后,在动态增长的细胞域上,局部增殖率与信号,我们得到不规则的图案(图6E) ●●●●。对于较低的值γ = 100,细胞簇高信号电平出现(图6F) ●●●●。总之,即使是相对简单的信号机制也可能导致显著不同的结果,这取决于组织的表现方式。
5讨论
我们开发了一个可扩展的、开源的基于细胞的模拟环境,用于研究形态发生问题。该新框架允许物理驱动的粘弹性细胞模型与调控信号模型的耦合模拟。实现了粘性耗散、弹性、平流、扩散、局部反应、局部质量源和汇、细胞分裂和细胞分化等过程。通过将我们的框架应用于图灵信号系统,我们表明信号系统在动态组织上的表现可能与在简单的连续组织表示上的表现不同。因此,我们主张在动态生长的细胞域上测试连续形态发生信号模型。
该框架允许研究各种机械调节机制。通过使细胞分裂方向依赖于信号线索,可以研究对宏观组织几何形状的影响。细胞迁移可以通过在特定细胞类型上引入梯度依赖力来建模。通过指定具有不同细胞-细胞连接强度的多种细胞类型,可以实现细胞分类。该框架专门用于研究信号和生物物理细胞特性的相互影响。
粘弹性细胞模型以高分辨率表示细胞形状,因此与顶点模型不同,它不局限于密集的组织。此外,水动力相互作用、膜张力和静水压力是模型的组成部分。速度场在整个区域都可用,这是解释信号成分平流的一个关键优势,因此可以对细胞内浓度进行空间描述。然而,该模型不容易扩展到三维。由于需要对表面进行网格划分,预计算法和计算复杂性将非常重要,并将取决于未来的工作。然而,所提出的框架对于研究本质上的2D形态发生问题是理想的,例如之前也通过法哈迪法等。(2007)和石原慎久(2012)二维。
基金
作者承认SNF Sinergia赠款“间充质干细胞软骨内骨化发育工程”和SNF SystemsX RTD NeurostemX的资助。
利益冲突:未声明。
工具书类
等. (
2012
)BMP受体相互作用导致肢体发育过程中的数字模式
.科学。代表。
,2
,991
.
等. (
1997
)声音刺猬(Shh)在小鼠胚胎肺生长和形态发生中的作用
.开发
,124
,53
–63
.
等人. (
2012
)模拟表明,一个简单的网络足以控制肺分支形态发生过程中的分支点选择、平滑肌和血管形成
.生物开放
,1
,775
–788
.
(
1998
)流体流动的格子Boltzmann方法
.每年。流体力学版次。
,30
,329
–364
.
等. (
2005
)约翰逊-肯德尔-罗伯茨理论在活细胞中的应用
.物理。修订版Lett。
,94
,028102
.
等. (
2007
)物理在多细胞系统生长和模式形成中的作用:我们可以从基于单个细胞的模型中学到什么?
J.Stat.物理。
,128
,287
–345
.
等. (
2007
)细胞力学、细胞间相互作用和增殖对上皮细胞堆积的影响
.货币。生物。
,17
,2095
–2104
.
(
2004
)求解流体-粒子相互作用问题的浸入边界格子Boltzmann方法
.J.计算。物理学。
,195
,602
–628
.
等. (
1998
)活胚胎组织粘弹性特性的定量研究
.生物物理学。J。
,74
,2227
–2234
.
(
1972
)生物模式形成理论
.凯伯内提克
,12
,30
–39
.
(
1992
)基于二维扩展Potts模型的生物细胞分选模拟
.物理。修订版Lett。
,69
,2013
–2016
.
等. (
2002
)Boussinesq方程的耦合格子BGK模型
.国际期刊数字。方法流体
,39
,325
–342
.
(
2010
)基于细胞的多细胞系统仿真软件
.生物信息学
,26
,2641
–2642
.
(
2012
)形态发生过程中力动力学的贝叶斯推断
.J.西奥。生物。
,313
,201
–211
.
(
2013
)基于配体受体的图灵机制控制下的肾分支形态发生
.物理。生物。
,10
,46003
.
等. (
2012
)肺发育早期的分支模式选择
.公共科学图书馆计算。生物。
,8
,e1002377号
.
等人. (
2014
)几何形状和信号的相互作用使肺分支形态发生强劲
.开发
,141
,4526
–4536
.
等. (
2011
)VirtualLeaf:基于细胞的植物组织生长和发育建模的开源框架
.植物生理学。
,155
,656
–666
.
等. (
2011
)单元几何形状对分区平面定位的影响
.单元格
,144
,414
–426
.
(
2005
)使用亚细胞元素建模多细胞系统
.数学。Biosci公司。工程师。
,2
,613
–624
.
(
2003
)浸没边界法
.数字学报
,11
,479
–517
.
等. (
2009
)Chaster:一种测试驱动的生物建模软件开发方法
.计算。物理。Commun公司。
,180
,2452
–2471
.
等. (
1993
)反应扩散方程的格子Boltzmann计算
.化学杂志。物理学。
,98
,1514
.
(
2007
)模拟单个细胞生长的浸入式边界框架:在肿瘤早期发展中的应用
.J.西奥。生物。
,247
,186
–204
.
等. (
2004
)绒毛滋养层双层生长力学的计算模型
.牛市。数学。生物。
,66
,199
–232
.
等. (
2014
)形态原DPP对图案形成和生长的协调作用
.货币。生物。
,24
,245兰特
–255兰特
.
等. (
2011年a
)鸡胚原始条纹形成过程中大尺度涡旋运动的“趋化偶极子”机制
.物理。生物。
,8
,045008
.
等. (
2011年b
)主动生物力学过程亚细胞模型中的涌现细胞和组织动力学
.物理。生物。
,8
,045007
.
(
1979
)具有极限环行为的简单化学反应体系
.J.西奥。生物。
,81
,389
–400
.
等. (
2012
)使用CompuCell3D对组织进行多尺度建模。方法细胞生物学。
,325
–366
.
(
2013
)长骨发育模型中相互依赖的组织生长和图灵模式
.物理。生物。
,10
,56009
.
(
1952
)形态发生的化学基础
.菲尔翻译。R.Soc.B生物学。科学。
,237
,37
–72
.
等. (
2007
).模拟可变形液囊的浸没边界格子Boltzmann方法及其在微观血流中的应用
.物理。生物。
,4
,285
–295
.
作者注释
©作者2015。牛津大学出版社出版。保留所有权利。有关权限,请发送电子邮件至:journals.permissions@oup.com








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}