")
")
")
")
")
")
主要功能和细节
兔HMGB1多克隆 适用人群:ICC/IF、WB 敲除已验证 反应对象:小鼠、大鼠、人类 同位素:IgG
选择批间可重复性更高的重组抗体


研究可靠 —— 各批次间结果一致且可重复 长期批量供应 —— 采用重组技术,可实现快速生产 首次实验即可成功 —— 经过大量验证确认了特异性 符合伦理标准 —— 产品不含动物成分
概述
-
产品名称 -
描述 兔多克隆抗体 至HMGB1 -
宿主 兔子 -
经测试应用 -
种属反应性 与反应: 老鼠、老鼠、人 预测可用于: 兔子、奶牛 
-
免疫原 -
阳性对照 WB:HAP1全细胞裂解物、NIH/3T3、MEF1、PC-12、HeLa、Jurkat、A431、HEK-293细胞。 ICC:HeLa细胞。
-
常规说明 多年来,生命科学行业一直处于再现性危机之中。 Abcam正在通过我们的一系列重组单克隆抗体和用于金标准验证的敲除编辑细胞系引领解决这一问题。 购买前请检查此产品是否满足您的需求。 如果您有任何问题、特殊要求或疑虑,请在购买之前向我们发送查询和/或联系我们的支持团队。 以下是本产品的推荐替代品,以及出版物、客户评论和问答
性能
-
形式 液体 -
存放说明 在4°C下装运。 短期储存在+4°C下(1-2周)。 交付时,等分。 储存温度为-20°C或-80°C。 避免冷冻/解冻循环。 -
存储溶液 pH值:7.40 防腐剂:0.02%叠氮化钠 成分:PBS 浓度<1mg/ml的该产品批次可添加BSA作为稳定剂。 如果您想了解特定批次的配方信息,请联系我们的科学支持团队,他们会很乐意提供帮助。 -

浓度信息加载。。。 -
纯度 免疫原亲和力纯化 -
克隆 多克隆 -
同种型 IgG抗体 -
研究领域
相关产品
-
ChIP相关产品 -
兼容的辅助设备 -
免疫肽(阻断) -
同位素控制 -
重组蛋白 -
相关产品
应用
靶标
-
功能 多功能氧化还原敏感蛋白,在不同的细胞隔室中具有不同的作用。 细胞核中是主要的染色质相关非组蛋白之一,作为DNA伴侣参与复制、转录、染色质重塑、V(D)J重组、DNA修复和基因组稳定性。 建议成为一种通用的核酸生物传感器。 促进宿主对无菌和感染信号的炎症反应,并参与先天性和适应性免疫反应的协调和整合。 细胞质中作为免疫原核酸的传感器和/或伴侣,涉及TLR9介导的免疫反应的激活,并介导自噬。 作为危险相关分子模式(DAMP)分子,在组织损伤期间增强免疫反应。 释放到细胞外环境可以结合DNA、核小体、IL-1β、CXCL12、AGER亚型2/sRAGE、脂多糖(LPS)和脂磷壁酸(LTA),并通过多种表面受体的参与激活细胞。 在细胞外室中,完全减少的HMGB1(由坏死释放)作为趋化因子,二硫键HMGB1作为细胞因子(积极分泌),磺酰HMGB1则(由凋亡细胞释放)促进免疫耐受(PubMed:23519706,PubMed:33446148,PubMet:23994764,PubMer:25048472)。 具有促血管生成活性(根据相似性)。 可能参与血小板活化(根据相似性)。 与磷脂酰丝氨酸和磷脂酰乙醇酰胺结合(通过相似性)。 与RAGE结合介导神经元生长信号(通过相似性)。 可能在扩大聚谷氨酰胺(polyQ)蛋白质的积累中发挥作用,例如亨廷顿蛋白(HTT)或TBP(PubMed:23303669,PubMed:25549101)。 核功能归因于HGMB1的完全降低。 与染色质结合并结合DNA,优先选择非标准DNA结构,如单链DNA、含DNA的十字形或弯曲结构、超螺旋DNA和ZDNA。 可以弯曲DNA并通过环化增强DNA的灵活性,从而通过增强转录因子结合和/或使远程调控序列接近,提供一种机制来促进各种基因启动子的活性(PubMed:20123072)。 可能在核苷酸切除修复(NER)中起增强作用(通过相似性)。 然而,使用体外系统在NER中的效果却有矛盾的报道(PubMed:19446504,PubMed:19360789)。 可能参与错配修复(MMR)和基底切除修复(BER)通路(PubMed:15014079,PubMed:16143102,PubMet:17803946)。 可能参与双绞线断裂修复,如非同源端接(NHEJ)(通过相似性)。 通过充当RAG复合物的辅因子参与V(D)J重组:通过刺激切割和RAG蛋白在保守重组信号序列(RSS)的23bp间隔区结合起作用(通过相似性)。 体外可以从高度弯曲的DNA中置换组蛋白H1(根据相似性)。 可以重组标准核小体,导致转录因子结合的结构约束放松(通过相似性)。 增强固醇调节元件结合蛋白(SREBPs)如SREBF1与其同源DNA序列的结合,并增加其转录活性(通过相似性)。 促进TP53与DNA的结合(PubMed:23063560)。 建议以与HSPB1相关的转录依赖方式参与线粒体质量控制和自噬; 然而,这一功能受到了质疑(通过相似性)。 可以调节端粒酶复合体的活性,并可能参与端粒的维持。 在细胞质中,建议通过与BECN1的竞争性相互作用分离BECN1:BCL2复合物,导致自噬激活(PubMed:20819940)。 参与氧化应激介导的自噬(PubMed:21395369)。 可以保护BECN1和ATG5免受钙蛋白酶介导的分裂,从而控制其促自噬和促凋亡功能,并调节炎症相关细胞损伤的程度和严重程度(通过相似性)。 在髓系细胞中,通过促进自噬对内毒素血症和细菌感染具有保护作用(通过相似性)。 参与巨噬细胞对CpG-DNA的反应中TLR9的内体移位和激活。 细胞外室(主动分泌或被动释放后)参与炎症反应的调节。 完全降低的HGMB1(释放后会被氧化)与CXCL12相关,在组织损伤的初始阶段介导炎症细胞的募集; CXCL12:HMGB1复合物触发CXCR4同二聚化(PubMed:22370717)。 诱导单核细胞衍生的未成熟树突状细胞的迁移,并似乎调节与AGER/RAGE和ITGAM相关的中性粒细胞的粘附和迁移功能(通过相似性)。 可结合各种类型的DNA和RNA,包括微生物非甲基化CpG-DNA,以增强对核酸的先天免疫反应。 建议在混杂的DNA/RNA传感中发挥作用,该传感与特定模式识别受体随后的区分传感相配合(通过相似性)。 促进与AGER/RAGE相关的细胞外DNA诱导的AIM2炎症小体激活(PubMed:24971542)。 二硫HMGB1与跨膜受体结合,如AGER/RAGE、TLR2、TLR4和可能的TREM1,从而激活其信号转导途径。 调节细胞因子/趋化因子的释放,如TNF、IL-1、IL-6、IL-8、CCL2、CCL3、CCL4和CXCL10(PubMed:12765338,PubMed:18354232,PubMet:19264983,PubMed:20547845,PubMed:24474694)。 促进巨噬细胞刺激的自然杀伤(NK)细胞与其他细胞因子如IL-2或IL-12协同分泌干扰素-γ(PubMed:15607795)。 TLR4被认为是促进巨噬细胞活化的主要受体,通过TLR4进行信号传递似乎与LY96/MD-2有关(PubMed:20547845)。 在细菌LPS或LTA介导的炎症反应中,炎症反应与内毒素结合,并将其转移到CD14,以向相应的TLR4:LY96和TLR2复合物发出信号(PubMed:18354232,PubMed:21660935,PubMet:25660311)。 通过与ACER/RAGE的关联促进肿瘤增殖(通过相似性)。 可通过IL1R1:IL1RAP受体复合物结合IL1-beta和信号(PubMed:18250463)。 与A类CpG结合可激活与TLR9、MYD88和AGER/RAGE相关的浆细胞样树突状细胞中的细胞因子生成,并可激活自身反应性B细胞。 通过含HMGB1的染色质免疫复合物,还可以通过B细胞受体(BCR)依赖性和ACER/RAGE依赖性机制(通过相似性)促进B细胞对内源性TLR9配体的反应。 抑制巨噬细胞吞噬凋亡细胞; 该功能依赖于多聚腺苷二磷酸核糖化,并与凋亡细胞细胞表面的磷脂酰丝氨酸结合(根据相似性)。 适应性免疫可能通过激活效应T细胞和抑制调节性T(TReg)细胞来增强免疫(PubMed:15944249,PubMed:22473704)。 相反,在不涉及效应或调节性T细胞的情况下,表达淋巴毒素LTA:LTB异源三聚体的T细胞需要肿瘤浸润和激活,从而促进肿瘤恶性进展(通过相似性)。 也有报道限制T细胞增殖(根据相似性)。 释放HMGB1:凋亡过程中形成的核小体复合体可以通过TLR2发出信号,诱导细胞因子产生(PubMed:19064698)。 参与凋亡细胞诱导免疫耐受; 当凋亡细胞释放其促炎活性时,活性氧物种(ROS)依赖的氧化作用(特别是Cys-106)可中和其促炎作用(PubMed:18631454)。 在巨噬细胞活化过程中,活化的淋巴细胞衍生的自凋亡DNA(ALD-DNA)促进ALD-DNA向内体的募集。 -
组织特异性 无处不在。 以血小板表达(PubMed:11154118)。 -
序列相似性 属于HMGB家族。 包含2个HMG盒DNA结合结构域。 -
结构域 HMG盒2介导促炎细胞因子刺激活性并与TLR4结合(PubMed:12765338,PubMed:20547845)。 然而,在凋亡诱导免疫耐受的背景下,不参与介导免疫原活性(PubMed:24474694)。 酸性C末端结构域形成柔性结构,其可以与HMG盒分子内可逆地相互作用,并调节与DNA和其他蛋白质的结合(PubMed:23063560)。 -
翻译后修饰 丝氨酸残基磷酸化。 NLS两个区域的磷酸化是细胞质易位和分泌所必需的(PubMed:17114460)。 LPS刺激后在多个部位乙酰化(PubMed:22801494)。 核定位信号(NLS1和NLS2)中赖氨酸残基的乙酰化导致细胞质定位和随后的分泌(通过相似性)。 Lys-3上的乙酰化导致优先结合DNA末端并削弱DNA弯曲活性。 半胱氨酸残基Cys-23、Cys-45和Cys-106的还原/氧化,以及可能的分子内二硫键,包括Cys-23和Cys-46,在不同的细胞隔室中产生具有特定功能活性的不同氧化还原形式:1-完全还原HMGB1(HMGB1C23hC45hC106h),2-二硫基HMGB1 和3-磺酰基HMGB1(HMGB1C23soC45soC106so)。 LPS刺激后分泌的PARP1可使多聚ADP-核糖化。 CASP1的体外裂解释放出一种含有HMG盒1的肽,该肽可能介导免疫原性活性; 该肽拮抗凋亡诱导的免疫耐受(PubMed:24474694)。 可被凝血酶水解:血栓调节蛋白复合物; 减少与肝素的结合和促炎活性。 -
细胞定位 核心。 染色体。 细胞质。 保密。 细胞膜。 内体。 内质网-高尔基体中间隔室。 基态主要为核。 在细胞质和细胞核之间穿梭(PubMed:12231511,PubMed:17114460)。 在自噬刺激下从细胞核转移到细胞质(PubMed:20819940)。 巨噬细胞在细胞外环境中的释放需要激活NLRC4或NLRP3炎性体(根据相似性)。 通过扩散将坏死细胞被动释放至细胞外环境,包括完全还原的HGMB1,随后被氧化(PubMed:19811284)。 也由凋亡细胞释放(PubMed:16855214,PubMed:18631454)。 各种免疫细胞和非免疫细胞(如巨噬细胞、单核细胞、中性粒细胞、树突状细胞和自然杀伤细胞)对各种刺激(如脂多糖和细胞因子)的主动分泌涉及通过分泌溶酶体的非常规分泌过程(PubMed:12231511,PubMed:14532127,PubMet:15944249)。 血浆细胞对LPS的反应而分泌(通过相似性)。 发现于活化血小板表面(PubMed:11154118)。 -
UniProt提供的信息 -
数据库链接 Entrez基因: 282691 奶牛 Entrez基因: 3146 人类 Entrez基因: 100862258 鼠标 Entrez基因: 15289 鼠标 Entrez基因: 100328641 兔子 Entrez基因: 25459 老鼠 Omim公司: 163905 人类 瑞士保护银行: 第10103页 奶牛
查看所有内容 -
别名 两性激素抗体 染色体蛋白、非组蛋白、HMG1抗体 DKFZp686A04236抗体
查看所有内容
图片
-

免疫细胞化学/免疫荧光-抗-HMGB1抗体(ab18256) ab18256染色HeLa细胞HMGB1。 用4%多聚甲醛固定细胞(10分钟),用0.1%PBS-Triton X-100渗透5分钟,然后用1%牛血清/10%正常山羊血清/0.3M甘氨酸在0.1%PBS吐温中封闭1小时。 然后将细胞在4°C下以1µg/ml的ab18256培养过夜,并 约7291 ,小鼠单克隆[DM1A]对α-微管蛋白-负载控制。 然后将细胞与 约150081 ,山羊抗兔IgG多克隆第二抗体-H&L(Alexa Fluor ® 488),以1/1000稀释度预吸附(以绿色显示)和 ab150120型 小鼠IgG-H&L(Alexa Fluor)山羊多克隆二级抗体 ® 594),以1/1000稀释度预吸附(以伪品红色显示)。 用DAPI标记细胞核DNA(以蓝色显示)。 使用高含量分析仪(Operetta CLS、Perkin Elmer)采集图像,并显示共焦截面的最大强度投影。 -

Western blot-抗-HMGB1抗体(ab18256) 所有车道: 抗-HMGB1抗体(ab18256),浓度为1µg/ml 车道1: 野生型HAP1全细胞裂解物 车道2: HMGB1敲除HAP1全细胞裂解物 每车道20µg的裂解液/蛋白质。 预测的带宽大小: 25千Da 1-4车道: 合并信号(红色和绿色)。 绿色-在29 kDa下观察到ab18256。 红色-装载控制, 约9484 ,在37 kDa下观察到。 ab18256显示在野生型HAP1细胞中识别HMGB1,因为信号在HMGB1敲除细胞中的预期MW处丢失。 在野生型和敲除型细胞中观察到额外的交叉反应带。 对野生型和HMGB1基因敲除样品进行SDS-PAGE分析。 Ab18256和 约9484 (小鼠抗GAPDH负荷对照)分别以1μg/ml和1/2000稀释度在4°C下孵育过夜。 用山羊抗兔IgG H&L(IRDye ® 800CW)预吸收床 ab216773号 和山羊抗鼠IgG H&L(IRDye ® 680RD)预吸收床 约216776 二级抗体在成像前在室温下以1/20000稀释1小时。 -

Western blot-抗-HMGB1抗体(ab18256) 所有车道: 1µg/ml的抗HMGB1抗体(ab18256) 车道1: NIH/3T3全细胞裂解物( 约7179 ) 车道2: MEF1(小鼠胚胎成纤维细胞系)全细胞裂解液 3号车道: PC-12(大鼠肾上腺嗜铬细胞瘤细胞系)全细胞裂解液 每车道10µg的裂解液/蛋白质。 次要 1号和3号车道: IRDye 680结合山羊抗狂犬病IgG(H+L),1/10000稀释 车道2: IRDye 680结合山羊抗狂犬病IgG(H+L),1/10000稀释 在还原条件下进行。 预测的带宽大小: 25千Da 观察到的频带大小: 29千Da 为什么实际的频带大小与预测的不同? 其他波段位于: 59kDa。 我们不确定这些额外波段的身份。 -

Western blot-抗-HMGB1抗体(ab18256) 所有车道: 抗-HMGB1抗体(ab18256),浓度为1µg/ml 车道1: HeLa(宫颈腺癌人上皮细胞系)全细胞裂解物 车道2: Jurkat全细胞裂解物( ab7899型 ) 3号车道: A-431全细胞裂解物( 约7909 ) 车道4: HEK-293全细胞裂解物( 约7902 ) 每车道10µg的裂解液/蛋白质。 次要 所有车道: IRDye 680结合山羊抗狂犬病IgG(H+L),1/10000稀释 在还原条件下进行。 预测的带宽大小: 25千Da 观察到的频带大小: 29千Da 为什么实际的频带大小与预测的不同? -
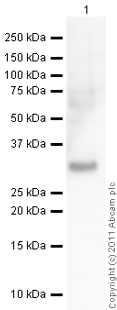
Western blot-抗-HMGB1抗体(ab18256) -

Western blot-抗-HMGB1抗体(ab18256) 这张图片是由Francesca Cerbai博士提交的Abreview提供的 所有车道: 稀释1/1000的抗-HMGB1抗体(ab18256) 车道1: 大鼠脑全组织裂解液-注入asf 1周 车道2: 大鼠脑全组织裂解液-注入LPS 1周 3号车道: 大鼠脑全组织裂解液-注入acsf 8周 车道4: 大鼠脑全组织裂解液-注入LPS 8周 车道5: 大鼠脑全组织裂解液-注入LPS 4周 车道6: 大鼠脑全组织裂解液-注入LPS 4周,在注入LPS 2周后,用海桑烷治疗2周 7号车道: 大鼠脑全组织裂解液-注入LPS 4周,在注入LPS 2周后用美金刚治疗2周。 每车道40µg的裂解液/蛋白质。 次要 所有车道: 生物素化山羊抗兔IgG 使用ECL技术开发。 在还原条件下进行。 预测的带宽大小: 25千Da 暴露时间: 30秒
数据表及文件
-
SDS下载 -
数据表下载
文献 (496)
印地语N 等。 铂基方案对成人晚期儿童型横纹肌肉瘤有效,并取决于HMGB1的表达。 国际分子科学杂志 24:不适用(2023年)。 公共医学:36614297 蔡B 等。 Gasdermin E通过小鼠视网膜中的全反式受体介导光感受器损伤。 生物化学杂志 298:101553 (2022). 公共医学:34973334 卡米亚M 等。 以肌纤维坏死为靶点可改善炎症性肌病。 国家公社 13:166 (2022). 公共医学:35013338 高Y 等。 miR-340-5p通过靶向抑制HMGB1抑制胰腺腺泡细胞炎症和凋亡。 实验治疗学 23:140 (2022). 公共医学:35069821 张XL 等。 HMGB1-促中性粒细胞外陷阱导致小鼠心脏舒张功能障碍。 J Am心脏协会 11:e023800(2022)。 公共医学:35156391