摘要
这个从头开始通过细胞色素P450侧链切割(CYP11A1)酶活性将胆固醇转化为孕烯醇酮,从而在线粒体中开始产生类固醇和神经类固醇。转运体蛋白(TSPO)的C末端氨基酸结构域已被证明能结合胆固醇,从而决定其线粒体易位。本研究的目的是研究位于TSPO区域的Ala147Thr单核苷酸多态性对健康志愿者孕烯醇酮生成的影响。在以循环淋巴单核细胞为代表的外周细胞模型中评估孕烯醇酮的生成。首先,证实了CYP11A1在mRNA和蛋白质水平上的表达。孕烯醇酮的产量在基因型组之间存在差异。孕烯醇酮平均值的比较表明,与Ala147纯合个体相比,Thr147纯合子或杂合子个体的孕烯醇激素水平显著降低。这些发现表明,次要等位基因变异体Thr147在产生类固醇生成途径的第一个代谢物方面具有主导作用。有趣的是,与杂合个体相比,Ala147纯合个体的循环富含胆固醇的低密度脂蛋白水平显著升高。总之,我们的结果表明,TSPO内的Ala147Thr自发氨基酸取代能够影响孕烯醇酮的生成;这将鼓励进一步研究其在多基因血脂异常中的潜在作用。
在合成类固醇的细胞中从头开始类固醇生成始于线粒体内膜细胞色素P450侧链裂解酶(CYP11A1)将胆固醇转化为孕烯醇酮。类固醇的形成速度取决于胆固醇从细胞内储存物运输到线粒体内膜的速度(1–三). 调节胆固醇向线粒体转运的关键因素是普遍存在的线粒体转运体蛋白(TSPO),以前称为外周型苯二氮卓受体(2–6). 已提出TSPO的一种假定的五跨膜结构,其中C末端暴露在细胞质中,其中发现了最后一个跨膜结构域,其中包含胆固醇识别氨基酸一致序列[L/V-X(1-5)-Y-X(1-5R)](7–9). 特别是,TSPO C末端结构域(L144-S159)中的胆固醇识别氨基酸一致序列可以适合胆固醇分子,并可以定义胆固醇移位的入口门(7,8,10). C末端结构域在物种间具有很好的保守性。然而,在TSPO区域发现了两种单核苷酸多态性(SNP)(rs6971,rs6972),分别将丙氨酸147转变为苏氨酸(Ala147Thr)和精氨酸162转变为组氨酸(Arg162His)(http://www.ncbi.nlm.nih.gov/sites/entrez). 这些SNP与精神疾病相关(11,12)因为类固醇和神经甾醇在调节几个中枢神经系统功能中起着重要作用(13–15). 在本研究中,我们研究了Ala147Thr单核苷酸多态性是否会影响健康志愿者的孕烯醇酮生成。由于缺乏可评估的典型类固醇生成组织,我们在以循环淋巴单核细胞为代表的外周细胞模型中进行了研究。我们选择了一种原始的淋巴细胞/制剂,因为已经充分证明这种细胞表达TSPO(16–20)进一步的细胞纯化步骤需要从参与研究的受试者中采集大量血液。
为此,我们首先评估了人类细胞模型中CYP11A1酶在mRNA和蛋白质水平上的表达以及酶活性。
材料和方法
学科
共有232名健康志愿者参与了这项研究。所有受试者(意大利裔白种人)捐献唾液用于基因组DNA提取和Ala147Thr SNP基因分型。采集33名受试者的血样(隔夜禁食后0900至1030小时之间采集),以进行孕烯醇酮测定,并采集71名受试人员的血样,以评估以下循环胆固醇参数:总胆固醇(TC)、甘油三酯(TG)、高密度脂蛋白(HDL)和低密度脂蛋白。本研究的排除标准为妊娠、高血压、肥胖、糖尿病、酗酒、类固醇或神经活性药物治疗。
该研究设计得到比萨大学伦理委员会的批准。所有受试者均被告知研究程序的性质,并在参与前提供书面知情同意书。
DNA基因分型
使用JETQUICK血液和细胞培养DNA自旋试剂盒(德国罗纳GENOMED GmbH)从唾液细胞中分离出所有受试者的基因组DNA。Ala147Thr SNP(rs6971)的基因分型是使用先前描述的PCR限制性片段长度多态性方法的修改进行的(21). PCR混合物的总体积为30μl,由0.5 ng/μl基因组DNA、200μl米每个dNTP的100 n米正向和反向引物(正向引物:5′-TGGACAGGCACTTGGGTGAAC-3′;反向引物:5′-AAGCGTGGACGGCCACATCA-3′),1.25米米氯化镁2和1 U AmpliTaq Gold(加利福尼亚州福斯特市应用生物系统公司)放在配备的缓冲区中(一次)。进行热循环时,95℃下的初始变性为5分钟,然后在94℃下进行30次循环,每次1分钟,66℃下进行30s,72℃下进行1min,最后在72℃下延长7分钟。根据制造商的建议,用5 U Nru I消化PCR产物,并在2.5%超纯琼脂糖三硼酸EDTA凝胶(马里兰州盖瑟斯堡生命技术公司)上通过电泳分离。为了保证质量,5%的样品被盲目重复,每次消化都使用已知基因型的额外样品进行。
孕烯醇酮测量
循环电池样品的制备
血样被抽入含有肝素锂的试管中,然后按照博尤姆的方法进行处理以制备单核细胞(22). 将最终的细胞颗粒重新悬浮在完整的RPMI 1690培养基中,补充15%的胎牛血清,2 m百万升-谷氨酰胺、100 U/ml青霉素和100 mg/ml链霉素。细胞以约300000个细胞/孔的密度接种在96个平底板(Sarstedt,Newton,NC)中,并保持在5%CO的湿润大气中2-95%空气在37℃下放置24小时。为了评估细胞群,使用随机细胞样本(n=7),使用标准技术进行流式细胞术分析。淋巴细胞和单核细胞根据其物理散射特性进行门控。通过流式细胞术前向散射检测(FSC)分析细胞大小与侧面散射(SSC)点图。细胞在488nm处激发,FSC和SSC信号放大模式设置为线性范围。在Becton Dickinson FACScan流式细胞仪(BD Becton Dickinson Italia,Buccinasco,Italia)上进行分析,获得至少10000个总事件,并在筛选活细胞后使用Cell Quest软件。通过台盼蓝排除法和流式细胞术分析评估,存活率大于90%。
孕烯醇酮合成酶表达的表征
在随机细胞样本(n=7)中,分别使用RT-PCR和Western-blot分析在mRNA和蛋白质水平上评估CYP11A1的表达。
对于RT-PCR分析,使用总体积为14μl的quantiTect转录试剂盒逆转录1μg总RNA。两微升逆转录混合物用于PCR。用AmpliTaq Gold DNA聚合酶(Applied Biosystems)在95℃下扩增33个周期,持续1分钟,57℃下扩增30秒,72℃下扩增1分钟。PCR引物如下:sense,5′-GAGATGCCACGCAACCTGA-3′;反义,5′-CCCCACATCGCTGAGGTGT-3′。对两微升扩增产物进行半巢式PCR。用AmpliTaq Gold DNA聚合酶(Applied Biosystems)在95℃下进行33个周期的半巢式PCR扩增,扩增时间为1分钟,57℃下为30秒,72℃下为1分钟;反义,5′-CCCCACATCGCTGAGGTGT-3′。PCR产物在2%NuSieve-GTG琼脂糖凝胶(Lonza Group Ltd.,Basel,Switzerland)上通过琼脂糖电泳分离,在紫外线下通过溴化乙锭染色进行可视化,然后根据制造商的说明使用QIAquick凝胶提取系统进行纯化(Qiagen s.p.a.,Milan,Italy)。纯化的PCR产物由Primm测序服务机构(意大利米兰Primm srl)使用ABI 3730自动DNA测序仪(Applied Biosystems)进行测序。在两个方向上进行测序以确认发现。使用FinchTV软件1.4版(Goespiza Inc.,西雅图,华盛顿州)分析序列,并与人类CYP11A1 mRNA的序列CYP11A1[人类CYP111A1 mRNA国家生物技术信息中心参考序列:NM_000781(Bethesda,MD)]对齐。作为阳性对照,类固醇生成的人类细胞系ADF[从世界卫生组织IV级人类多形性胶质母细胞瘤获得(23,24)]已使用。
对于Western blot分析,将细胞蛋白样品(50μg)置于12.5%十二烷基硫酸钠-聚丙烯酰胺凝胶上,并转移到硝化纤维素膜上。将膜在含有0.05%吐温20的脱脂奶粉的5%印迹级阻断剂中阻断数小时,并在室温下用1:1000稀释的一级抗体孵育2小时[Millipore Corp.,Vimodrone,Italy;CYP11A1抗体:针对大鼠CYP11A1的421-441氨基酸序列开发,目录号AB1244;甘油醛-3-磷酸脱氢酶(GAPDH)抗体:在5%脱脂奶粉的印迹颗粒阻滞剂中针对人类GAPDH开发:目录号AB9132]。在Tris缓冲盐水中清洗三次10分钟后[10 m米Tris-HCl(pH 8)和150m米含有0.05%吐温20的NaCl],将膜与抗兔(针对CYP11A1抗体)或抗鸡(针对GAPDH抗体)辣根过氧化物酶结合二级抗体(1:5000)在室温下孵育1小时。在含有0.05%吐温20的Tris缓冲盐水中清洗膜三次。使用ECL Plus Western印迹剂(Amersham Pharmacia Biotech,Piscataway,NJ)将膜培养1分钟,并暴露于x射线胶片(Amersham Pharmaca Biotech)。使用Bio-Rad分析系统(Amersham Pharmacia Biotech,Piscataway,NJ)分析蛋白质浓度。类固醇原性胶质瘤细胞系ADF(23,24)作为阳性对照。
孕烯醇酮评估
33名受试者的淋巴细胞在无血清盐培养基[140 m米氯化钠,5米米KCl,10米米葡萄糖,1.8米米氯化钙2,1米米硫酸镁4,10米米HEPES(pH 7.4)和0.1%BSA]含有抑制剂三戊烷(25μ米)和SU10603(10μ米)37℃,在5%CO的增湿大气中2-95%空气。SU10603和trilostone分别是诺华农场(意大利瓦雷斯)和D Zister博士(爱尔兰都柏林大学)赠送的礼物。抑制剂用于确保细胞内不会发生孕烯醇酮的进一步代谢。如前所述,用酶免疫分析法(ELISA)测定细胞培养基中分泌的孕烯醇酮(25). 简单地说,保留细胞盐培养基并在1500×克用于在供应商建议的条件下定量测定孕烯醇酮。每种样品的ELISA均为一式三份。与其他类固醇的交叉反应通常小于1%,与孕酮的交叉反应为6%(德国汉堡IBL)。检测的灵敏度为0.05 ng/ml。使用单点竞争模型将未知样品与同时运行的孕烯醇酮标准品进行比较。
胆固醇参数测定
采用标准酶法评估71名受试者的TC、TG和HDL血浆浓度(德国诺伊斯Wako Chemicals GmbH)。使用Friedewald公式计算LDL(26).
统计分析
使用Hardy-Weinberg Simulator软件测试基因型偏离Hardy-Weinberg平衡(HWE)的程度(http://krunch.med.yale.edu/hsim/) (27). 对于具有两个等位基因的基因座,当等位基因频率和基因型比例之间的关系遵循等式p时,该基因座位于HWE中2+2个pq+q2=1,其中p和q分别是主要和次要等位基因的频率。皮尔逊χ2测试是为了测试性别频率的差异t吨进行测试以比较年龄的平均差异。孕烯醇酮含量和循环胆固醇参数表示为平均值±扫描电镜(东南方测量值)。采用单因素方差分析和Bonferroni检验比较基因型组间孕烯醇酮水平或循环胆固醇参数。P(P)<0.05被认为具有统计学意义。
结果
TSPO Ala147Thr单核苷酸多态性的基因分型
对232名白种人(31.9%男性,平均年龄36.5±13.6岁;68.1%女性,平均年龄37.3±10.9岁)进行了TSPO编码基因(SNP识别号rs6971)内Ala147Thr SNP的研究。基因型和等位基因分布总结于表1Ala147Thr SNP基因型分布符合HWE预期。在表1HapMap中报告的白种人中Ala147Thr SNP基因型和等位基因分布(http://www.ncbi.nlm.nih.gov/sites/entrez; 还显示了参考SNP编号rs6971)。研究样本中的Ala147Thr SNP基因型分布在女性和男性中分层,在性别或平均年龄之间没有显著差异(χ2= 4.59,P(P)= 0.10; 学生t吨试验t=0.48,P(P)= 0.629).
表1。当前研究和HapMap中TSPO Ala147Thr单核苷酸多态性的基因型和等位基因分布
| 高加索人. | n个. | 基因型n(%). | HWEχb条,P(P). | 等位基因n(%). |
|---|
| 阿拉147一. | 阿拉147Thrb条. | Thr147型c(c). | 阿拉147. | Thr147型. |
|---|
| 当前研究 | | | | | | | |
| 总计 | 232 | 113 (48.7) | 98 (42.2) | 21 (9.1) | 0.001, 0.97 | 324(69.8) | 140 (30.2) |
| 雌性 | 158 | 74(46.8) | 73 (46.2) | 11 (7.0) | | 221(69.9) | 92 (30.1) |
| 男性 | 74 | 39 (52.7) | 25 (33.8) | 10 (13.5) | | 103 (69.6) | 45 (30.4) |
| HapMap-CEU公司d日 | 60 | 28 (46.7) | 23 (38.3) | 9 (15.0) | | 79 (65.8) | 41 (34.2) |
| CEU_发电机_面板d日 | 59 | 28 (47.5) | 22 (37.3) | 9 (15.3) | | 78 (66.1) | 40 (33.9) |
| 高加索人. | n个. | 基因型n(%). | HWEχb条,P(P). | 等位基因n(%). |
|---|
| 阿拉147一. | 阿拉147Thrb条. | Thr147型c(c). | 阿拉147. | Thr147型. |
|---|
| 当前研究 | | | | | | | |
| 总计 | 232 | 113 (48.7) | 98 (42.2) | 21(9.1) | 0.001, 0.97 | 324 (69.8) | 140(30.2) |
| 雌性 | 158 | 74 (46.8) | 73(46.2) | 11 (7.0) | | 221 (69.9) | 92 (30.1) |
| 男性 | 74 | 39 (52.7) | 25 (33.8) | 10 (13.5) | | 103 (69.6) | 45 (30.4) |
| HapMap-CEU公司d日 | 60 | 28 (46.7) | 23 (38.3) | 9 (15.0) | | 79 (65.8) | 41 (34.2) |
| CEU_发电机_面板d日 | 59 | 28 (47.5) | 22 (37.3) | 9 (15.3) | | 78 (66.1) | 40 (33.9) |
表1。当前研究和HapMap中TSPO Ala147Thr单核苷酸多态性的基因型和等位基因分布
| 高加索人. | n个. | 基因型n(%). | HWEχb条,P(P). | 等位基因n(%). |
|---|
| 阿拉147一. | 丙氨酸147Thrb条. | Thr147型c(c). | 阿拉147. | 苏氨酸147. |
|---|
| 当前研究 | | | | | | | |
| 总计 | 232 | 113 (48.7) | 98 (42.2) | 21 (9.1) | 0.001, 0.97 | 324 (69.8) | 140 (30.2) |
| 雌性 | 158 | 74 (46.8) | 73 (46.2) | 11 (7.0) | | 221 (69.9) | 92 (30.1) |
| 男性 | 74 | 39 (52.7) | 25 (33.8) | 10 (13.5) | | 103 (69.6) | 45 (30.4) |
| HapMap-CEU公司d日 | 60 | 28 (46.7) | 23 (38.3) | 9 (15.0) | | 79 (65.8) | 41 (34.2) |
| CEU_发电机_面板d日 | 59 | 28 (47.5) | 22 (37.3) | 9 (15.3) | | 78 (66.1) | 40(33.9) |
| 高加索人. | n个. | 基因型n(%). | HWEχb条,P(P). | 等位基因(%). |
|---|
| 阿拉147一. | 阿拉147Thrb条. | 苏氨酸147c(c). | 阿拉147. | Thr147型. |
|---|
| 当前研究 | | | | | | | |
| 总计 | 232 | 113 (48.7) | 98 (42.2) | 21 (9.1) | 0.001, 0.97 | 324 (69.8) | 140 (30.2) |
| 雌性 | 158 | 74 (46.8) | 73 (46.2) | 11 (7.0) | | 221 (69.9) | 92 (30.1) |
| 男性 | 74 | 39 (52.7) | 25 (33.8) | 10 (13.5) | | 103 (69.6) | 45 (30.4) |
| HapMap-CEU公司d日 | 60 | 28 (46.7) | 23 (38.3) | 9 (15.0) | | 79 (65.8) | 41 (34.2) |
| CEU_发电机_面板d日 | 59 | 28 (47.5) | 22 (37.3) | 9(15.3) | | 78 (66.1) | 40 (33.9) |
孕烯醇酮测量
细胞群评估
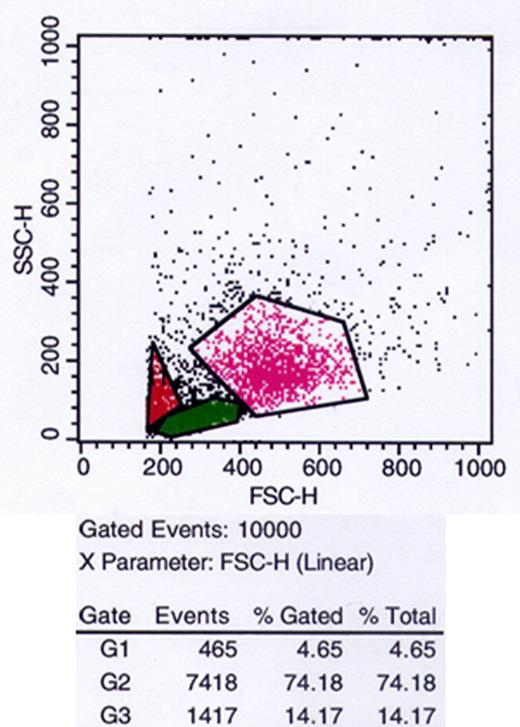
流式细胞术用于评估通过差速离心从健康个体血液样本(n=7)中分离的细胞群。使用正向和侧向散射特征来识别淋巴细胞和单核细胞。如散射细胞图所示图1,两个细胞群清晰可见。电子选择淋巴细胞和单核细胞(绿色和粉红色区域)并分析细胞数量。定量分析表明,样本中至少有75%的淋巴细胞和14%的单核细胞。24小时后死亡细胞的百分比约为5%。
F类免疫球蛋白. 1.
散射细胞图分析。散射细胞图分析显示按大小和粒度进行的细胞分离,通常用于识别不同的白细胞类型(绿色区域G2,淋巴细胞;粉红色区域G3,单核细胞;红色区域G1,死亡细胞)。流式细胞仪检测(FSC-H)得出的光散射参数与.SSC-H)以点图形式报告。在x轴和y轴上,报告了流式细胞仪检测器检测到的散射光单位。FSC和SSC信号放大模式设置为线性比例。给出了一个具有代表性的实验。
孕烯醇酮合成酶表达的表征
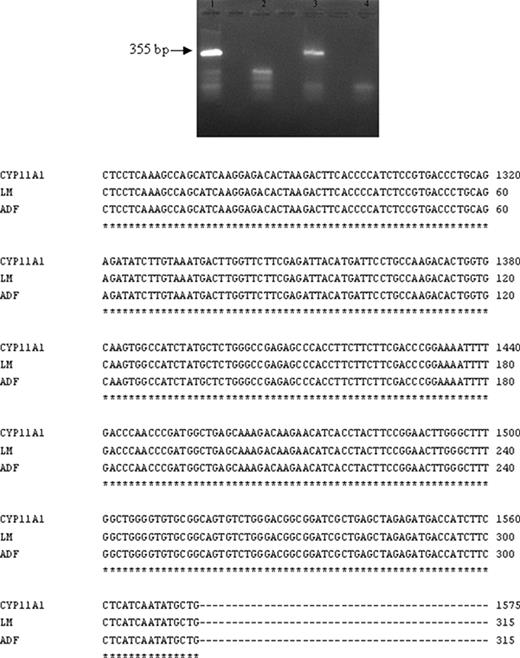
为了研究CYP11A1在淋巴单核细胞中的表达,进行了RT-PCR和蛋白质印迹分析(n=7)。作为阳性对照,人类类固醇生成胶质细胞系ADF(23,24)已使用。使用设计用于扩增人CYP11A1基因的第六和第九外显子的特异性引物,在总RNA中鉴定CYP11A1 mRNA的存在。在琼脂糖凝胶上一致检测到预期大小(355 bp)的片段,测序显示与GenBank(NM_000781)中保存的相应序列完全一致(图2).
F类免疫球蛋白. 2.
扩增产物的RT-PCR和核苷酸序列测定。在图的顶部,RT-PCR分析显示了人类淋巴细胞(3区)和阳性对照(ADF类固醇生成细胞系,1区)中预期大小的片段(355 bp)。没有逆转录酶的样本(第2和第4道)。对预期大小的片段(355 bp)进行纯化和测序,以检查获得的淋巴单核细胞(LM)和ADF细胞cDNA核苷酸序列的特异性。为此,将cDNA序列与GenBank数据库(GenBank登录号NM_000781)中报告的人类CYP11A1 mRNA序列进行比较。在图的底部,对淋巴单核细胞(LM)和ADF细胞获得的核苷酸序列的比较表明,它们与已发表的人类CYP11A1 mRNA序列完全一致。
使用针对大鼠CYP11A1 421–441氨基酸序列开发的商业多克隆抗体验证CYP11A1-蛋白的存在。如所示图3,抗CYP11A1抗体识别淋巴细胞和阳性对照ADF细胞系中的56.1-kDa人CYP11A1-蛋白(GenBank登录号NP_000772)。淋巴细胞中也存在较小的人CYP11A1亚型(42.0 kDa;GenBank登录号NP_001093243)。抗GAPDH抗体识别淋巴单核细胞和阳性对照ADF细胞系中的36.0-kDa人GAPDH蛋白(GenBank登录号NP_002037)。
F类免疫球蛋白. 3.
人CYP11A1酶的Western blot分析。抗CYP11A1抗体识别淋巴单核细胞(LM)或阳性对照ADF细胞系中的56.1-kDa人CYP11A11(GenBank登录号NP_000772)。淋巴细胞中也证实了较小的人CYP11A1亚型(42.0 kDa;GenBank登录号NP_001093243)。抗GAPDH抗体可识别淋巴单核细胞或ADF细胞系中36.0-kDa人GAPDH(GenBank登录号NP_002037)。
孕烯醇酮评估
通过测量细胞释放并在盐培养基中积累的孕烯醇酮,测定了33名健康人(36.4%男性,平均年龄30.6±7.9岁;63.6%女性,平均年龄30±9.7岁)淋巴细胞中CYP11A1的功能活性。约3×105细胞产生0.1165±0.04375 ng/ml孕烯醇酮。为了确定孕烯醇酮来自CYP11A1的催化活性,一些这样的淋巴单核细胞样本(n=10)用特异性CYP11A1抑制剂氨基谷酰胺(20μ米). 氨鲁米特治疗后未检测到孕烯醇酮水平。
基因型组孕烯醇酮产量的比较
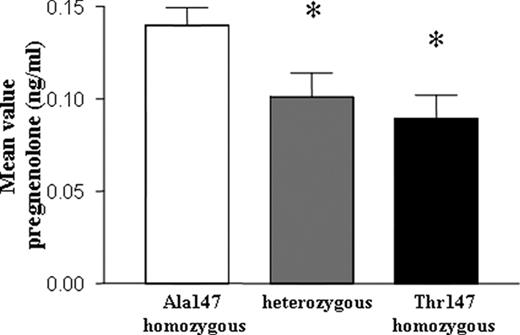
人类淋巴单核细胞产生的孕烯醇酮水平的平均值被分为TSPO Ala147Thr SNP的三个基因型组:Ala147纯合子个体(n=15)、杂合子个体(n=12)和Thr147纯合子个体(n=6)。孕烯醇酮水平的平均值为:Ala147纯合基因型=0.1397±0.0370 ng/ml;杂合基因型=0.1010±0.0449 ng/ml;Thr147纯合基因型=0.0895±0.0304 ng/ml(图4).
F类免疫球蛋白. 4.
基因型组孕烯醇酮产量的比较。孕烯醇酮平均值的比较显示,基因型组之间的总体差异具有统计学意义(ANOVA分析,P(P)= 0.013). Bonferroni的测试证明杂合子(t=2.554,P(P)<0.05)或Thr147纯合基因型(t=2.659,P(P)<0.05)显示与Ala147纯合基因型相比,孕烯醇酮水平显著降低。*,具有统计意义的结果。
孕烯醇酮平均值的比较显示,基因型组之间的总体差异具有统计学意义(ANOVA分析,P(P)= 0.013). 特别是,Bonferroni的测试显示杂合子(t=2.554,P(P)<0.05)和Thr147纯合基因型(t=2.659,P(P)与Ala147纯合基因型相比,孕烯醇酮水平显著降低(图4).
基因型组血浆胆固醇参数的比较
循环胆固醇参数的平均值被分层为TSPO Ala147Thr SNP的三个基因型组:Ala147纯合子个体(n=34)、杂合子个人(n=30)和Thr147纯合个体(n=7)。TC、LDL、HDL和TG的平均值以及基因型组间比较的统计分析报告于表2.
表2。TSPO Ala147Thr SNP基因型组胆固醇参数的平均值和统计分析
| . | 基因型组. | 方差分析F,P(P)价值. |
|---|
| 147Ala纯合子. | 杂合的. | 147Thr纯合子. |
|---|
| 总费用 | 195 ± 35 | 180 ± 24 | 179 ± 28 | 2.25, 0.11 |
| 低密度脂蛋白 | 116±31 | 100 ± 19 | 96 ± 22 | 3.82,0.03一 |
| 高密度脂蛋白 | 61 ± 12 | 63 ± 12 | 60±11 | 1.43, 0.74 |
| TG公司 | 77 ± 23 | 76 ± 29 | 97 ± 60 | 0.31, 0.25 |
| . | 基因型组. | 方差分析F,P(P)价值. |
|---|
| 147Ala纯合子. | 杂合的. | 147Thr纯合子. |
|---|
| 总费用 | 195 ± 35 | 180 ± 24 | 179 ± 28 | 2.25, 0.11 |
| 低密度脂蛋白 | 116 ± 31 | 100 ± 19 | 96 ± 22 | 3.82, 0.03一 |
| 高密度脂蛋白 | 61 ± 12 | 63 ± 12 | 60 ± 11 | 1.43, 0.74 |
| TG公司 | 77 ± 23 | 76 ± 29 | 97 ± 60 | 0.31, 0.25 |
表2。TSPO Ala147Thr SNP基因型组胆固醇参数的平均值和统计分析
| . | 基因型组. | 方差分析F,P(P)价值. |
|---|
| 147Ala纯合子. | 杂合的. | 147Thr纯合子. |
|---|
| 总费用 | 195 ± 35 | 180 ± 24 | 179±28 | 2.25, 0.11 |
| 低密度脂蛋白 | 116 ± 31 | 100 ± 19 | 96 ± 22 | 3.82, 0.03一 |
| 高密度脂蛋白 | 61 ± 12 | 63 ± 12 | 60 ± 11 | 1.43, 0.74 |
| TG公司 | 77 ± 23 | 76 ± 29 | 97 ± 60 | 0.31, 0.25 |
| . | 基因型组. | 方差分析F,P(P)价值. |
|---|
| 147Ala纯合子. | 杂合的. | 147Thr纯合子. |
|---|
| 总费用 | 195 ± 35 | 180 ± 24 | 179 ± 28 | 2.25, 0.11 |
| 低密度脂蛋白 | 116 ± 31 | 100 ± 19 | 96 ± 22 | 3.82, 0.03一 |
| 高密度脂蛋白 | 61 ± 12 | 63 ± 12 | 60 ± 11 | 1.43, 0.74 |
| TG公司 | 77 ± 23 | 76±29 | 97 ± 60 | 0.31, 0.25 |
基因型组间唯一表现出统计显著差异的胆固醇参数是LDL(方差分析,P(P)=0.03)。具体来说,Bonferroni的测试显示杂合子组(t=2.48,P(P)<0.05)与Ala147纯合基因型相比,LDL平均值显著降低(表2). 由于在Ala147纯合组和杂合组之间只能发现中等显著性差异,因此这种差异在生物学上确实非常显著。与Ala147纯合子组相比,Thr147纯合基因型没有显示出显著差异(t=1.87,P(P)> 0.05).
讨论
在本研究中,我们研究了TSPO基因中的Ala147Thr单核苷酸多态性是否会影响健康人淋巴细胞中孕烯醇酮的产生。
我们首先确定了样本中Ala147Thr单核苷酸多态性的等位基因频率,该样本由232名健康个体组成,并测试了这些基因型是否偏离HWE。HWE测试可以揭示种群中各种现象的发生,例如自然选择或随机交配。此外,它通过检测不适当的基因分型方案引入的系统偏差,提供了更高水平的质量保证。Ala147Thr SNP基因型分布符合HWE预期。TSPO单核苷酸多态性显示微小等位基因的频率为30.2%,编码氨基酸苏氨酸147。这个和我们以前的结果(12)有助于更好地估计高加索人群中Ala147Thr SNP的频率,因为与HapMap数据库中报告的样本相比,基因型分布是在更大的样本中获得的(http://www.ncbi.nlm.nih.gov/sites/entrez; 参考SNP ID rs6971)(参见表1). 据我们所知,该SNP的基因型分布评估仅在大样本日本个体中进行(11,21,28). 在之前的遗传学研究中,已经评估了Ala147Thr单核苷酸多态性是否会影响特定精神疾病的易感性。特别是,在白人人群中,已发现Ala147Thr单核苷酸多态性与焦虑的中间表型、成人分离焦虑障碍有关(12).
Ala147Thr单核苷酸多态性在侵袭性和趋化性不同的人类乳腺癌细胞系中也有报道;然而,这种多态性与特定的侵袭性表型无关(29).
对于孕烯醇酮分析,我们首先验证了人类淋巴单核细胞中第一种类固醇生成酶CYP11A1的存在和功能活性,该酶催化胆固醇转化为孕烯醇醇酮,被选为外周循环细胞模型。通过RT-PCR和测序分析鉴定CYP11A1基因的特异转录物(图2). 在这些细胞中,鉴定出人类CYP11A1多肽的两种亚型,即亚型a(GenBank登录号NP_000772,分子量56.1 kDa(图3). 亚型a的表达也已在主要不具有合成类固醇激素功能的组织中得到鉴定(30–34). 关于亚型b的可用数据较少(http://www.ncbi.nlm.nih.gov/protein网站). CYP11A1酶活性通过测量细胞培养基中在没有和存在特异性CYP11A1-抑制剂氨基谷氨酰亚胺的情况下孕烯醇酮的生成来评估,以确定特异性酶活性。我们的结果与先前证明CYP11A1酶在其他物种免疫细胞群中存在和/或活性的其他报告一致(30,35). 人淋巴细胞合成孕烯醇酮的平均值为0.1165 ng/3×105如预期的那样,该值远低于主要用于合成类固醇激素的细胞(36–39) (即MA-10 Leydig和Y-1肾上腺皮质细胞系(~1×10)产生的孕烯醇酮为4.9±0.06 ng和21±0.9 ng5单元格)。
我们发现TSPO Ala147Thr氨基酸取代与健康个体淋巴单核细胞中孕烯醇酮产生的特定水平有关。与Ala147纯合个体相比,Thr147纯合子或杂合子个体的淋巴细胞产生的孕烯醇酮水平显著降低(图4). 这些结果表明Thr147等位基因变体具有显性效应。Thr147等位基因的一个拷贝的存在可能影响胆固醇易位的功效。有人认为TSPO的C末端结构域可以容纳胆固醇分子,这可能定义了胆固醇移位的入口门(7,8,10). 通过对大鼠TSPO序列的定点突变,发现C末端结构域内的氨基酸Y152、Y153和R156参与了TSPO与胆固醇的相互作用(7,8). 据我们所知,本研究首次调查了人类TSPO C末端自发突变在类固醇生成中的潜在作用。我们的发现为理解遗传变异如何促进和影响人类类固醇分子合成提供了一个起点。我们无法得出147Thr等位基因变异携带者孕烯醇酮生产受损可能影响哪种类固醇生成途径的结论。事实上,孕烯醇酮可以转化为不同的类固醇,这取决于所使用的特定酶途径。尽管存在这种局限性,但由于类固醇分子参与多种生物功能,这种TSPO多态性可能与这些疾病的易感性/保护性有关(40,41)这与类固醇的生成减少/增加有关。
有趣的是,健康人的Ala147Thr单核苷酸多态性与血浆低密度脂蛋白水平特别相关。与Ala147基因型个体相比,杂合子个体的低密度脂蛋白水平显著降低。Ala147Thr单核苷酸多态性与血脂异常之间的因果关系目前尚不能得出结论。然而,这些结果鼓励进一步研究Ala147Thr单核苷酸多态性对冠心病患者胆固醇代谢的潜在影响。可以推测,以产生孕烯醇酮能力下降为特征的Thr147携带者细胞可能上调受体以增加血浆胆固醇内流。事实上,已经证明,缺乏胆固醇线粒体易位中TSPO相关蛋白(类固醇生成性急性调节蛋白)的啮齿类动物肾上腺表达高水平的受体,这些受体内化循环中富含胆固醇的脂蛋白(42).
总之,我们发现Ala147Thr单核苷酸多态性与细胞产生类固醇生成途径第一代谢物孕烯醇酮的能力呈正相关。类固醇和神经甾醇在正常生理功能和疾病中的相关性鼓励在基础科学水平以及在人类临床研究背景下继续研究TSPO多态性。
这项工作得到了意大利大学和科学研究部的支持(PRIN 2005:协议2005069159)。
披露摘要:作者无需声明任何内容。
缩写:
CYP11A1、,
FSC、,
GAPDH、,
高密度脂蛋白,
硬件设备,
低密度脂蛋白,
SNP、,
SSC、,
TC、,
TG、,
TSPO、,
1斯托科
DM公司
2000
线粒体内胆固醇转移。
Biochim生物物理学报
1486
:184
–197
2Rone公司
MB(MB)
,风扇
J
,帕帕佐普洛斯
五
2009
类固醇生物合成中的胆固醇转运:蛋白质相互作用的作用和疾病状态的意义。
Biochim生物物理学报
1791
:646
–658
三米勒
禽流感
2007
类固醇生成性急性调节蛋白(StAR),一种新型线粒体胆固醇转运蛋白。
Biochim生物物理学报
1771
:663
–676
4帕帕佐普洛斯
五
,勒卡尼
L(左)
,棕色
钢筋混凝土
,汉族
Z
,姚明
ZX公司
2006
神经甾体生物合成、神经病理学和神经疾病中的外周型苯二氮卓受体。
神经科学
138
:749
–756
5帕帕佐普洛斯
五
,巴拉尔迪
M(M)
,吉拉尔特
信托收据
,努森
结核
,拉卡佩
JJ公司
,林德曼
P(P)
,诺伦伯格
医学博士
,螺母
D类
,魏茨曼
A类
,张
先生
,加维什
M(M)
2006
转定位蛋白(18kDa):基于其结构和分子功能的外周型苯二氮卓受体的新命名。
药物科学趋势
27
:402
–409
6加维什
M(M)
,巴赫曼
我
,舒克伦
R(右)
,卡茨
Y(Y)
,韦恩曼
L(左)
,魏辛格
G公司
,魏茨曼
A类
1999
外周苯二氮卓受体之谜。
药理学修订版
51
:629
–650
7锂
H(H)
,帕帕佐普洛斯
五
1998
外周型苯二氮卓类受体在胆固醇转运中的作用。确定假定的胆固醇识别/相互作用氨基酸序列和共识模式。
内分泌学
139
:4991
–4997
8杰明
N个
,诺依曼
吉咪
,奥斯图尼
妈妈
,Vu公司
TK公司
,姚明
ZX公司
,Murail公司
S公司
,罗伯特
JC公司
,贾扎基斯
C类
,帕帕佐普洛斯
五
,拉卡佩
JJ公司
2005
外周型苯二氮卓受体胆固醇识别氨基酸一致序列的表征。
分子内分泌学
19
:588
–594
9Murail公司
S公司
,罗伯特
JC公司
,成本
YM公司
,诺依曼
吉咪
,奥斯图尼
妈妈
,姚明
ZX公司
,帕帕佐普洛斯
五
,杰明
N个
,拉卡佩
JJ公司
2008
核磁共振测定转运蛋白TSPO跨膜结构域的二级和三级结构。配体结合后TSPO三级折叠的稳定性。
Biochim生物物理学报
1778
:1375
–1381
10锂
H(H)
,姚明
Z
,德根哈特
B类
,Teper公司
G公司
,帕帕佐普洛斯
五
2001
外周型苯二氮卓受体胆固醇识别/相互作用氨基酸共识(CRAC)上的胆固醇结合以及HIV TAT-CRAC肽对类固醇生成的抑制。
《美国科学院院刊》
98
:1267
–1272
11中村
K(K)
,山田
K(K)
,岩山
Y(Y)
,丰田
T型
,古鲁川
A类
,泷本
T型
,寺山
H(H)
,岩桥
K(K)
,塔凯
N个
,米纳贝
Y(Y)
,关根
Y(Y)
,铃木
K(K)
,岩手
Y(Y)
,皮莱
A类
,中本(Nakamoto)
Y(Y)
,池田
K(K)
,吉井
M(M)
,福西
我
,吉川
T型
,莫里
N个
2006
外周苯二氮卓受体(PBR)基因变异影响惊恐障碍易感性的证据
.美国医学遗传学杂志B神经精神病学遗传学
141B个
:222
–226
12科斯塔
B类
,皮尼
S公司
,马提尼
C类
,阿贝利
M(M)
,加贝洛尼
P(P)
,兰迪
S公司
,Muti公司
M(M)
,杰西
C类
,拉里
L(左)
,卡尔迪尼
A类
,加尔代里西
S公司
,穆奇
A类
,卢卡奇尼
A类
,卡萨诺
国标
2009
易位蛋白中的Ala147Thr替代与抑郁症患者的成人分离焦虑相关。
精神病学遗传学
19
:110
–111
13梅尔坎吉
钢筋混凝土
,加西亚-塞古拉
LM公司
,门萨-尼亚甘
AG公司
2008
神经活性类固醇:最新进展和新观点。
细胞分子生命科学
65
:777
–797
14博琉
受试者
1998
神经类固醇:大脑的一种新功能。
心理神经内分泌学
23
:963
–987
15克里斯滕森
斯里兰卡
,施特劳斯三世
JF公司
2000
类固醇生成性急性调节蛋白(StAR)与胆固醇的线粒体内易位。
Biochim生物物理学报
1529
:175
–187
16亚力山大
比利时
,滚轮
E类
,科洛茨
U型
1992
人类淋巴细胞和淋巴瘤细胞系外周型苯二氮卓结合位点的特征及其在细胞生长中的作用。
生物化学药理学
44
:269
–274
17卡纳
X(X)
,卡拉永
P(P)
,布瓦博拉
M(M)
,卡阿尔
D类
,夏尔
D类
,罗克牌手表
C类
,勒弗尔
G公司
,卡塞利亚斯
P(P)
1993
外周型苯二氮卓类受体在人类造血细胞上的分布概况和性质。
生命科学
52
:107
–118
18别尔科维奇
A类
,费拉雷塞
C类
,卡瓦莱蒂
G公司
,Alho公司
H(H)
,马卓拉蒂
C类
,比安奇
G公司
,吉多蒂
A类
,科斯塔
E类
1993
人类淋巴细胞中两个DBI受体的拓扑结构。
生命科学
52
:1265
–1277
19前田
S公司
,宫崎骏
T型
,中西
T型
,多岐川
M(M)
,岛田
M(M)
1998
富含T淋巴细胞制剂中的外周型苯二氮卓受体。
生命科学
63
:1423
–1430
20韦恩曼
L(左)
,加维什
M(M)
2006
外周型苯二氮卓受体与心血管系统。对药物开发的影响。
药物治疗学
110
:503
–524
21库鲁马吉
A类
,野本
H(H)
,吉川
T型
,大久保
Y(Y)
,托鲁
M(M)
2000
在日本样本中,苯二氮卓受体(外周)基因的两种错义变异与精神分裂症之间的关联研究。
J神经传输
107
:491
–500
22博爱
A类
1968
从人类血液中分离单核细胞和粒细胞。用一次离心分离单核细胞,用离心和沉降相结合的方法分离粒细胞。
Scand J临床实验室投资供应商
97
:77
–89
23马洛尼语
W公司
,拉伊纳尔迪
G公司
,里瓦贝内
R(右)
,圣蒂尼
机器翻译
1994
不同的细胞死亡敏感性t吨-过氧化氢丁基可能依赖于细胞组织型相关的生长特征。
细胞生物毒素
10
:207
–218
24法布里齐
C类
,科拉桑蒂
M(M)
,佩尔西基尼
T型
,布西纳罗
R(右)
,Starace公司
G公司
,劳罗牌手表
总经理
1994
γ干扰素上调人星形细胞瘤细胞中α-2巨球蛋白的表达。
神经免疫杂志
53
:31
–37
25达·塞蒂莫
F类
,西莫里尼
F类
,塔利亚尼
S公司
,拉莫塔
C类
,马里尼
调幅
,萨勒诺
S公司
,贝兰迪
M(M)
,诺维里诺
E类
,格雷科
G公司
,科西梅利
B类
,达波佐
E类
,科斯塔
B类
,西莫拉
N个
,莫雷利
M(M)
,马提尼
C类
2008
抗焦虑作用N个,N个-二烷基-2-苯基吲哚-3-基乙醛酰亚胺通过调节转位蛋白促进神经甾体生物合成。
化学
51
:5798
–5806
26弗里德沃尔德
重量
,征收
RI公司
,弗雷德里克森
DS公司
1972
在不使用制备型超速离心机的情况下估算血浆中的低密度脂蛋白胆固醇浓度。
临床化学
18
:499
–502
27立方体
JF公司
,小林寺
K(K)
,Nagatsu公司
T型
,基德
KK公司
,基德
JR公司
,卡拉费尔
F类
,克兰茨勒
人力资源
,一之濑
H(H)
,格伦特尔
J
1997
多巴胺β羟化酶基因(DBH)功能变体的群体遗传学。
美国医学遗传学杂志
74
:374
–379
28库鲁马吉
A类
,野本
H(H)
,山田
K(K)
,吉川
T型
,托鲁
M(M)
2001
在日本样本中,苯二氮卓受体(外周)基因的两种错义变异与情绪障碍没有关联。
美国医学遗传学杂志
105
:172
–175
29哈德威克
M(M)
,费尔蒂克
D类
,可的
M(M)
,锂
H(H)
,维迪奇
B类
,帕帕佐普洛斯
五
1999
人类乳腺癌中的外周型苯二氮卓类受体(PBR):乳腺癌细胞侵袭性表型与PBR表达、核定位以及PBR介导的细胞增殖和胆固醇的核转运的相关性。
癌症研究
59
:831
–842
30奥卡
H(H)
,荣盛
Y(Y)
,林下(Hayashi)
Y(Y)
,野本
K(K)
2000
植入B16黑色素瘤的小鼠模型中CD4+T细胞中Th细胞免疫反应和类固醇生成CYP11A1表达的分解。
细胞免疫
206
:7
–15
31斯洛明斯基
A类
,戈梅兹·桑切斯
总工程师
,Foecking公司
MF公司
,沃茨曼
J
2000
正常大鼠皮肤中活跃的类固醇生成。
Biochim生物物理学报
1474
:1
–4
32特普柳克
NM公司
,张
Y(Y)
,卢
Y(Y)
,霍斯
JR公司
,哈桑
MQ公司
,特普柳克
不及物动词
,普拉塔普
J
,加林多
M(M)
,斯坦因
JL公司
,斯坦因
GS公司
,廉
接线盒
,范维恩
AJ公司
2009
成骨转录因子runx2控制参与固醇/类固醇代谢的基因,包括成骨细胞中的CYP11A1。
分子内分泌学
23
:849
–861
33涩屋
K(K)
,高田
N个
,霍霍
Y(Y)
,古鲁川
A类
,Yasumatsu公司
N个
,扩散膜
T型
,Enami公司
T型
,铃木
K(K)
,塔纳比
N个
,石井
H(H)
,无界
H(H)
,高桥
T型
,服部
助教
,卡瓦托
S公司
2003
海马细胞色素P450合成脑神经甾体,是突触信号转导的旁分泌神经调节剂。
Biochim生物物理学报
1619
:301
–316
34莫拉莱斯
A类
,线索
A类
,拉米雷兹
J
,维尔奇斯
F类
,迪亚兹·萨·切兹
五
1999
胰腺中类固醇的合成:细胞色素P-450scc活性的证据。
胰腺
19
:39
–44
35莱希纳
O(运行)
,迪特里希
H(H)
,威格斯
GJ公司
,瓦奇奥
M(M)
,灯芯
G公司
2001
鸡法氏囊和胸腺中的糖皮质激素生成。
国际免疫学
13
:769
–776
36西施
Y(Y)
,亚纳斯
T型
,亩
Y(Y)
,奥巴
K(K)
,一野
我
,斋藤
M(M)
,野村证券
M(M)
,穆卡萨
C类
,奥卡比
T型
,转到
K(K)
,Takayanagi公司
R(右)
,鹿岛
Y(Y)
,哈吉
M(M)
,纳瓦塔
H(H)
2001
表达功能性卵泡刺激激素受体的类固醇生成人类颗粒样肿瘤细胞系KGN的建立和鉴定。
内分泌学
142
:437
–445
37罗默茨
FF公司
,国王
SR公司
,跨度
PN编号
2001
MA-10细胞中孕酮代谢对准确测量类固醇生成率的影响。
内分泌学
142
:5236
–5242
38约翰逊
迈克科尔斯
,桑德森
JT公司
,隆德
BO公司
2002
3-MeSO2-DDE和一些CYP抑制剂对H295R人肾上腺皮质癌细胞株糖皮质激素类固醇生成的影响。
体外毒物
16
:113
–121
39坎皮亚尼
G公司
,纳奇
五
,菲奥里尼
我
,德菲利皮斯
最大功率
,加罗法洛
A类
,恰尼
性虐待
,格雷科
G公司
,诺维里诺
E类
,威廉姆斯
直流
,齐斯特尔
DM公司
,伍兹
美赞臣
,米哈伊
C类
,曼佐尼
C类
,梅尼尼
T型
1996
吡咯苯并氮杂卓衍生物的合成、生物活性和SARs,这是一类新的特异性“外周型”苯二氮杂卓受体配体。
化学
39
:3435
–3450
40Rone公司
MB(MB)
,风扇
J
,帕帕佐普洛斯
五
2009
类固醇生物合成中的胆固醇转运:蛋白质相互作用的作用和疾病状态的意义。
Biochim生物物理学报
1791
:646
–658
41杜布罗夫斯基
BO公司
2005
精神病理学中的类固醇、神经活性类固醇和神经甾醇。
神经心理药物生物精神病学进展
29
:169
–192
42曹
G公司
,赵
L(左)
,斯坦格尔
H(H)
,长谷川
T型
,理查森
青年成就组织
,帕克
吉隆坡
,霍布斯
HH(小时)
1999
小鼠B类1型清道夫受体的发育和激素调节
.分子内分泌学
13
:1460
–1473
版权所有©2009内分泌学会








{kind=link}
{kind=link}
{kind=link}
{kind=link}