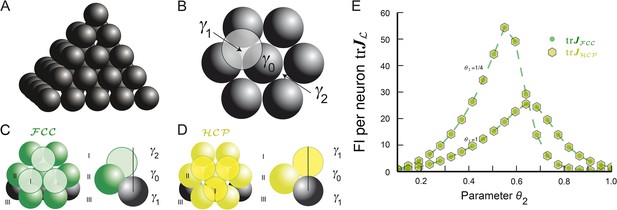
水果通常排列在形成(图5A). 一个人从一层六边形球体开始到达这个格子。这需要指定两个基向量,这是2D中最密集的填充。为了在3D中最大化填充比,下一层六边形排列的球体必须尽可能紧密地堆叠。第三个基向量和最后一个基向量有两种选择,表示为γ1和γ2在里面图5B(模数六角对称)。如果有人选择γ1,则下面两层没有中心位于该位置的球体γ1,但却有一个γ2(反之亦然)。层的堆叠如所示图5C并生成格子。
3D中的晶格和非晶格解决方案。
(A类)球体的堆叠,如格子。在这个3D中密度最大的晶格中,每个球体与其他12个球体接触,每个节点有四个不同的平面六角形晶格。(B类)在以γ0(黑色)可以从六个位置之一开始放置另一个六边形层,其中两个位置高亮显示,γ1和γ2. (C类)如果按照顺序(…,γ1,γ0,γ2,…)获得注意,层I中的球体与层III中的球体不对齐(D类)按照顺序(…,γ0,γ1,γ0,…)导致六角密封填料再次,每个球体与其他12个球体接触。然而,通过每个节点只有一个平面,其中球体中心的排列是正六边形晶格。此填料与,但不是晶格。(E类)用于缓冲功能Ω和用于各种参数组合θ1和θ2;θ1调节衰变和θ2支持。这两种填料具有相同的填充比,对于该调谐曲线,也提供相同的空间分辨率。FI:费希尔信息。
通过选择,可以达到相同的密度γ1对于顶层和底层以下的层。然而,正如这种排列,称为六角形密排(),不能用三个向量描述,它没有定义晶格(请参见图5D)尽管它和此类填料,定义为相等的非重叠球的排列(康威和斯隆,1992年;Hales,2012年),泛化格。
虽然可以为定义网格模块任何格子,如我们上面所示,一个不能由于缺乏对称性,以有意义的方式为任意封装定义网格模块。但对于任何给定的包装属于通过球在半径为1的情况下,可以通过推广格的定义来定义“网格单元”(公式7). 为此,考虑通过对于每个位置有一个独特的沃罗诺伊细胞V(V)第页带节点一个定义网格单元的调整曲线通过根据用于调节形状Ω和距离根据具体的包装,此调整曲线可能是周期性的,也可能不是周期性的。因为包装通常对称性比格子少,任意单元格中的“网格单元格”通常不能用于定义“网格模块”。要解释原因,请考虑任意包装和独特的Voronoi电池V(V)0包含点0的。选择M(M)均匀分布相c(c)1,…,c(c)M(M)在内部V(V)0.内的位置V(V)0然后将被移位的调谐曲线均匀覆盖然而,典型地,不同的Voronoi细胞既不一致,也不具有相似的体积。因此,Ω我通常会不用相同的密度覆盖每个Voronoi单元,因此无法定义合适的网格模块。格子不存在此问题。这里是等价类用相同的密度覆盖每个细胞。
另一方面,高度对称的填料确实允许定义网格模块。例如,六角密封填料可用于定义网格单元。使用相同的对称参数方程式9–11,意味着金融机构:
(18)
最大内半径R(右)对于具有网格大小λ=1等于1/2。与晶格一样,我们假设supp(Ω)=[0,R(右)]和B类R(右)(0) ⊂V(V)0然后被积函数在距离0大于1/2时消失。因此,我们得到:
(19)
考虑相同的调谐形状Ω和相位数M(M)对于点阵,其内半径也最大为1/2,式13为格子:
(20)
由于两个基本域具有相同的卷,即,,并且限制在这些球上的被积函数是相同的,即,,我们可以得出如下结论:网格模块包括或-相似的对称性具有相同的FI。我们还数值计算了网格单元格并将其与案例。对于凹凸型调谐曲线Ω,两个FI是相同的(图5E)从Ω的径向对称性可以看出。因此,网格单元定义为或对称性提供最佳分辨率。
图5D、E显示循环序列(γ0,γ1)和(γ1,γ0,γ2)导致和分别是。中心γ0,γ1、和γ2也可以用来说明填料的最后一点:有无限多不同的填料具有相同的密度它们可以由不相等的单词构成,这些单词是通过带字母的三角形有限遍历生成的γ0,γ1、和γ2(Hales,2012年),每个字母代表层的三个方向之一。例如(γ0,γ1,γ0,γ2)描述了具有相同密度的另一种填料。所有填料都有一个特点:每个球体周围正好有12个球体,排列在其中之一或格子式样(Hales,2012年). 这些填料也可用于定义网格模块,因为所有单元中的相密度都是均匀的。此外,在计算FI时和(方程式18–20)只有本地整合是必要的,这样的混合包装将有与纯或填料。
只有在最近几年,才证明没有其他安排比一个被称为开普勒猜想的问题(Hales,2005年,2012). 根据这些结果和我们对和(图5E),我们预测3D网格单元将对应于这些填料之一。虽然在3D中,填料密度与晶格密度相等,但在2D中却不是这样。从而证明了六角形晶格是所有平面填料中密度最大的一种(1910年,星期四); 2D中的网格单元应当具有六边形晶格结构。




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}