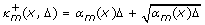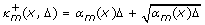滑动窗法 本文提出的算法的核心思想是计算近似值 属于 对 ( t吨 ) 以迭代方式,如等式( 6 ). 更准确地说,我们计算一系列近似值 这样对于子集 W公司
j个 状态空间的, j个 ∈ {1, ..., 第页 }, 为所有人 x个 ∈ W公司
j个 .集合 W公司 1 , ..., W公司
第页 被称为 窗户 ,我们假设 W公司
j个 包含时间间隔内概率质量(大部分)集中的状态[ t吨 j个 -1 , t吨
j个 ). 我们讨论窗户的构造 W公司
j个 稍后。
让 问
j个 是指的矩阵 W公司
j个 ,即我们定义 问
j个 ( x个 , x’ )= 问 ( x个 , x’ )如果 x个 , x’ ∈ W公司
j个 、和 问
j个 ( x个 , x’ )否则=0。 注意,为了演示的简单性,我们保留了一个固定的枚举 S公司 并假设每个 问
j个 大小与相同 问 然而,该方法的实现仅考虑 问
j个 包含中的状态条目 W公司
j个 。对于 τ
j个 = t吨
j个 - t吨 j个 -1 ,我们定义
哪里 = ( 1
年 ) T型 和 D类
j个 是对角线矩阵,其主要对角线项为 x个 ∈ W公司
j个 否则为零。 行向量( 1
年 ) T型 位置处有一个 年 否则为零。
在 j个 -第步,矩阵 包含入住的可能性 τ
j个 时间单位在 W公司
j个 从一个国家到另一个国家。 作为初始概率,公式( 7 )使用近似值 适用于所有州 x个 ∈ W公司
j个 对角矩阵确保概率质量位于 W公司 j个 -1 \ W公司
j个 在计算过程中忽略,即仅忽略交点的元素 W公司 j个 -1 ∩ W公司
j个 向量中可以有非零项 D类
j个 。这是必要的,因为 问
j个 不包含以外状态的转换速率 W公司
j个 (这些状态很吸引人)。 直观地,向量 描述了概率质量在内部移动后的位置 W公司
j个 .
尽管 问
j个 不是CTMC的发电机,等式( 7 )对所有状态都有一个简单的解释 x个 ∈ W公司
j个 .让我们修复 j个 现在,让CTMC 与…相同 X(X) ,除了所有州 x’ ∉ W公司
j个 正在吸收(即一次 x’ 已到达,无法离开)。 让初始概率分布 是这样的 为所有人 x个 ∈ W公司
j个 。那么 ,对于所有人 x个 ∈ W公司
j个 .面向所有人 j个 ,向量 是次随机的,并且在每个步骤中其条目的总和都会减少,即,
概率质量“丢失”,因为我们不考虑条目 对于 x个 ∈ W公司 j个 -1 \ W公司
j个 ,当我们与相乘时 D类
j个 此外,我们失去了离开的可能性 W公司
j个 在下一个 τ
j个 时间单位,因为 是一个超突变矩阵。如果 j个 ,在时间间隔内[ t吨 j个 -1 , t吨
j个 )大部分概率质量保持在 W公司
j个 ,然后是近似误差 对所有人来说都很小 x个 ∈ S公司 .之后损失的概率质量 j个 近似产生的步长由下式给出
因此,如果公式( 7 )精确求解后,滑动窗方法的总近似误差为 η
第页 注意,公式( 8 )是矢量所有分量的误差之和 .
窗户构造 在迭代的每个步骤中,窗口 W公司
j个 必须选择错误 η
j个 保持较小。 如果是这样的话 W公司
j个 满足以下条件:(a)具有足够高的概率 X(X) ( t吨 j个 -1 ) ∈ W公司
j个 ,(b)离开的概率 W公司
j个 在时间间隔内[ t吨 j个 -1 , t吨
j个 )足够小。
要求(a)意味着 W公司
j个 包含支持 也就是说,一个子集 S公司
j个 ⊂ S公司 这样的话 很小。 在第一步中,我们设置 S公司 1 = { 年 }。 对于 j个 >1、窗户 W公司
j个 在之后构造 已计算。 我们修了一个小 δ >0并选择 S公司
j个 = { x个 | > δ }。 如果支持 较大且几乎均匀分布,可能需要构造 S公司
j个 这样的话 小于某个固定阈值。 然而,我们的实验结果表明,使用固定阈值会产生良好的结果,这使得对支持度进行排序的额外工作 在实践中是不必要的。 注意,要求(a)意味着 W公司
j个 和 W公司 j个 -1 横断。 因此,在每一步中,我们都会沿着概率质量移动的方向“滑动”窗口。
本节的后续部分重点讨论需求(b),其中有必要预测流程的未来行为。 找到集合的一种可能性 W公司
j个 满足要求的是对 t吨
j个 - t吨 j个 -1 初始状态为的时间单位 S公司
j个 。如果我们的目标是获得准确的近似值,那么这可能会很昂贵。 大多数模拟运行与系统的平均行为相对应。 然而,有些事件可能不太频繁,但仍有很大的可能性。 因此,我们提出了一种依赖于状态控制确定性近似的思想,即给定初始状态 z(z) ∈ S公司
j个 ,估计每个状态变量在下一个期间可以采用的最大值和最小值 τ
j个 = t吨
j个 - t吨 j个 -1 时间单位。 更确切地说,对于每个维度 d日 ∈ {1, 2, ..., n个 },我们计算值 , ∈ ℤ 这样的话
很小,其中 X(X)
d日 ( τ )是 d日 -随机向量的第个分量 X(X) ( τ ).
极值的计算 , 在几个州执行 z(z) 都是随机选择的。 我们的实验结果表明,当考虑10个以上的状态时,我们的结果的准确性没有提高。 让 A类
j个 ⊆ S公司
j个 是随机状态的集合。 通过计算 和 ,我们得到了时间间隔内每个状态变量的最大值和最小值的估计[ t吨 j个 -1 , t吨
j个 )在以下条件下 X(X) ( t吨 j个 -1 ) ∈ S公司
j个 .窗口 W公司
j个 现在被构造为 S公司
j个 以及其中的所有状态 和 也就是说, W公司
j个 等于
对于固定状态 z(z) ∈ A类
j个 我们利用马尔可夫链的规则结构来计算 和 .我们从状态开始 z(z) 并逐个更新状态变量。 我们假设在很短的时间间隔Δ内,反应类型的速率 R(右)
米 保持不变,即等于 α
米 ( z(z) ). 然后是 R(右)
米 -下一个Δ时间单位内的跃迁是带参数的泊松分布 α
米 ( z(z) )Δ. 我们可以通过预期来近似这个数字 α
米 ( z(z) )泊松分布的Δ。 注意,上述假设是有根据的,因为在耦合化学反应的情况下,倾向 α
米 ( x个 )在 x个 ,如果只考虑基本反应,即对应于单个机械步骤的反应,因此最多有两个反应物。 一般来说,反应可能有中间产物和/或平行反应路径。 然而,它们总是可以分解为基本反应。 由于我们对上限和下限感兴趣,我们还考虑了标准偏差 泊松分布。 我们假设,如果当前状态为 x个 ,在Δ时间单位内
至少 ,
至多
类型转换 R(右)
米 被带走了。 注意,例如, α
米 ( z(z) )Δ=1,则我们有91.97%的置信度,即反应的实际数量位于区间内
让 κ
米 ∈ 和 我 = 0, 1,.... 迭代
产生连续确定的最坏情况近似 X(X) ( t吨 + Δ), X(X) ( t吨 + 2Δ),... 在以下条件下 X(X) ( t吨 )= z(z) .对于函数 α
米 在状态变量中增长极快的情况下,迭代可能会产生错误的近似值,因为它基于这样的假设,即在[0,Δ)期间倾向是恒定的。
在生化反应网络的背景下, α
米 最多是二次的,因此公式( 11 )产生足够的结果。 对于给定的系统,我们执行等式中的近似( 11 )对于所有可能的组合 。可以跳过优先处理导致状态空间中相反方向的转换类型的组合,因为它们不会给出最坏情况边界。 例如,考虑示例(1) c(c) 1 = c(c) 2 = c(c) 三 = 1, z(z) =(10,10,100,0),Δ=0.01。 如果我们假设更多类型的反应 R(右) 2 和 R(右) 三 发生(比平均值少) R(右) 1 ,我们得到 、和 这意味着复杂分子的数量减少 x个 (1) = (14,12, 96, 2). 我们可以省略包含这两者的组合 和 .作为 R(右) 1 等同 R(右) 2 反之亦然,这些组合不会产生状态变量极值的良好近似值。 一般来说,反应网络的依赖关系图可能有助于识别那些最大化特定人群的组合(另请参阅实验结果部分)。
在续集中,每个选择的组合都被称为 分支 因为,对于固定 z(z) ,相应的迭代导致不同的后续 x个 ( 我 +1) 请注意,对于特定分支 米 ∈ {1, ..., k个 }我们修复 κ
米 = ,或 κ
米 = 为所有人 我 。迭代在以下时间结束 ⌈ τ
j个 /Δ ⌉ 步长(其中最后一个时间步长是余数而不是Δ),以及极值 和 由迭代期间状态变量的最小值和最大值给出。 更具体地说, 和 ,其中1≤ d日 ≤ n个 , x个 ( 我 ) = 、和 z(z) = x个 (0) .
计算 和 在表中的伪代码中进行了描述 1 (左栏),调用 ContDetApprox公司 。注意上标 我 指的是当前分支,而不是等式中的迭代( 11 )这在第19行中执行。 分支数量为2 n个 因为每个维度的最大值和最小值都是必要的。 在第17行中,我们根据当前分支决定 我 ,是否 κ
米 设置为 ,或 .
表1方法 ContDetApprox公司 (左)和主程序 s窗口 (右) 关于时间步长Δ的选择,我们建议动态选择Δ,以便 米 等式中的间隔( 10 )至少涵盖了相应泊松分布概率质量的80%。 显然,在更大的区间覆盖更多概率质量的情况下,该方法的准确性会提高。对于我们的实验结果,我们选择Δ,以便 λ
x个 ·Δ=1产生了足够准确的结果。
滑动窗口算法 在表的右栏中 1 我们描述了称为 s窗口 ,以伪代码表示。 第2-14行中的for循环实现了 通过连续计算向量 对
j个 从 对 j个 -1 输入ϵ是第13行中ODE的解引起的总近似误差的界。 阵列 t吨 包含时间实例 t吨 0 , ..., t吨
第页 对于我们的实验结果,我们比较了下面解释的两种不同的时间步进机制。 参数 δ 是用于删除这些状态以支持 对 j个 -1 概率小于 δ 。我们定义 S公司
j个 作为所有状态的集合 x个 为此 对 j个 -1 ( x个 )大于 δ 在第4行中。 请注意,对于 j个 =1组 S公司 1 仅包含初始状态 年 .在第6行中, 兰特 ( S公司
j个 , 国家数量 )返回一组 状态数 随机元素来自 S公司
j个 用于构造向量的 b条 + 和 b条 - 在第7-10行中。 窗口中所有状态的速率条目 W公司
j个 (参见公式( 9 ))在第11行中计算,所有剩余条目 问
j个 设置为零。 第13行调用了一个求解方法来计算 对
j个 从 对 j个 -1 例如,这可以是均匀化方法、ODE解算器或基于Krylov子空间近似的方法。 我们经过了一段时间 τ
j个 和相应的分数 近似误差。
我们可以从输出中计算概率质量的总损失 对
第页 通过 η
第页 = 1 - ∑
x个 对
第页 ( x个 ). 该值包括算法的两个近似误差:(1)离开窗口的概率 W公司
j个 在时间间隔内[ t吨 j个 -1 , t吨
j个 )(2)概率 这是由于窗口滑动而丢失的,通过与乘法得到 D类
j个 (参见公式( 7 )).
请注意,为了提高获得的精度,始终可以重复计算步骤。 更准确地说,我们可以通过增加等式中区间的置信度来确定更大的窗口( 10 )即,通过选择时间步长Δ 米 类型转换的最大/最小数目 R(右)
米 存在于具有一定置信度的区间内(例如,置信度为80%)。 然而,对于我们的实验结果,我们没有重复任何计算步骤,因为我们总是获得足够准确的结果。
时间间隔 对于我们的实验结果,我们比较了算法的两种不同的时间步进机制 s窗口 (见表 1 ,右侧)。 我们要么选择等距的时间步长 τ
j个 = τ ,对于所有人 j个 ,或者我们决定 τ
j个 在窗户施工期间 W公司
j个 (自适应时间步长)。 后一种方法产生更快的运行时间。 根据系统的动力学,长时间步长可能会导致三个问题:(1)窗口大,矩阵大 问
j个 可能超过工作记忆容量,(2)在长时间步长内,系统的动态可能会有很大差异 问
j个 (3)该窗口可能包含仅在更短的时间间隔内有效的状态。 另一方面,如果时间步长太小,则必须执行主循环的多次迭代,直到算法终止。 窗口几乎完全重叠,尽管每个步骤可能只需要很少的时间,但整个过程的计算成本可能很高。 一种可能性是固定窗口的大小并相应地选择时间步长。 但这也不一定导致算法运行时间短。 原因是,求解方法的时间复杂度不仅取决于表示窗口的矩阵的大小,还取决于其数学特性。
通过计算可以避免上述问题 τ 1 , ..., τ
第页 窗户施工期间 W公司
j个 如下所示。 我们计算了当时重要的状态数 t吨 j个 -1 并将其传递给 ContDetApprox公司 第9行(见表 1 ). 我们在Algorithm中运行while循环 ContDetApprox公司 (见表 1 ,left)直到(1)窗口至少有一定的大小,并且(2)窗口中的状态数超过当时有效状态数的两倍 t吨 j个 -1 第一个条件确保窗口超过某个最小大小,例如500个州。 第二个条件确保新窗口刚好足够大,可以将概率质量移动到 S公司
j个 更准确地说,它确保了 S公司 1 , S公司 2 ,... 不重叠,且后续集合相邻(如图所示 1 ). 请注意,这可以确保生成的窗口不包含许多仅在更短的时间间隔内有效的状态。
while-lop终止时,我们传递变量的值 时间 从 ContDetApprox公司 到 s窗口 并设置 τ
j个 价值 时间 显然,在 s窗口 我们添加了一个表示到目前为止经过的时间的变量,第2行中的for循环被替换为while循环,该循环在经过的时间超过时停止 t吨 随后,我们给出了滑动窗口方法的实验结果,其中我们以上述方式使用自适应时间步长。
数值求解方法 在本节中,我们介绍了两种数值求解算法的理论基础,即均匀化方法和Krylov子空间方法。 我们近似CME的全局解(参见方程( 5 ))以及算法第13行中要求的局部解决方案 s窗口 (另请参见等式( 7 )).
均匀化 均匀化方法可以追溯到Jensen[ 34 ]也称为Jensen方法、随机化或离散时间转换。 在计算机系统的性能分析中,这种方法很受欢迎,并且通常优于其他方法,如Krylov子空间方法和数值积分方法[ 19 , 39 ]. 最近,均匀化也被用于求解CME[ 40 – 42 ].
全局均匀化 让( X(X) ( t吨 ), t吨 ∈ ℝ ≥0 )是有限状态空间的CTMC S公司 一致化的基本思想是定义离散时间马尔可夫链(DTMC)和泊松过程。 DTMC随机地与 X(X) ,这意味着如果步数在[0, t吨 )给出,泊松过程跟踪时间,如下所述。
回想一下 λ
x个 是国家的退出率 x个 ∈ S公司 、和 我 是单位矩阵。我们定义了一个 均匀化率λ 这样的话 λ ≥最大值 x个 ∈ S公司 λ
x个 并构造 ,DTMC的转移矩阵 X(X) 注意 P(P) 定义自循环概率1- λ
x个 / λ 一个州的 x个 ,当且仅当 λ > λ
x个 。对于 k个 ≥1,随机矩阵 P(P) k个 包含 k个 -阶跃转移概率,如果 对 (0) 是初始分布 X(X) ,向量 周 ( k个 ) = 对 (0) P(P) k个 包含之后的状态概率 k个 DTMC中的步骤。 时间间隔[0, t吨 )具有带参数的泊松分布 λt 即。,
现在,方程( 5 )可以重写为[ 19 , 32 , 43 ]
等式( 13 )与等式相比,具有良好的性能( 5 ). 没有涉及负面指控,因为 P(P) 是一个随机矩阵,并且 λ > 0. 此外, 周 ( k个 ) 可以通过以下公式进行归纳计算
如果 P(P) 稀疏, 周 ( k个 ) 即使状态空间的大小很大,也可以有效地进行计算。求和下限和上限 L(左) 和 单位 可以获得每个状态的 x个 截断误差[ 44 ]
可以是预先定义的误差容限ϵ>0的先验界。 因此, 对 ( t吨 ) 可以通过以下公式以任意精度逼近
只要所需的汇总数量不是特别大。
时间复杂性和刚度 作为 λt 使泊松分布变平,左截断点变大 L(左) 在等式中( 16 )在中线性增长 λt ,而有效泊松概率项的数量为[ 44 ] O(运行) ( ). 如果向量 周 ( L(左) ) , 周 ( L(左) +1) , ..., 周 ( 单位 ) 使用以下公式计算 单位 矩阵-向量乘法(参见公式( 14 )),则均匀化过程的复杂性为 O(运行) ( νλt )其中 ν 是中非零元素的数量 P(P) .
如果基础模型为 僵硬的 在刚性模型中,底层系统的组件在不同数量级的时间尺度上运行,这在各种应用领域中都会出现,尤其是在系统生物学中。 对于刚性模型,均匀化率 λ ≥最大值 x个 ∈ S公司 λ
x个 将对应于最快的时间刻度。 相比之下,只有在对应于最慢时间尺度的一段时间内,才能观察到缓慢分量的显著变化。 因此,均匀化方法非常耗时,因为 刚度指数 [ 45 ] t吨 ·最大值 x个 ∈ S公司 λ
x个 .
在后续部分中,我们展示了如何以局部方式应用均匀化,从而使刚度对该方法的性能产生较少的负面影响。 换句话说,滑动窗口技术使均匀化即使对于刚性系统也能表现良好。
局部均匀化 我们现在结合了均匀化和滑动窗口方法。 假设 S公司 可能是无限的,我们迭代地应用均匀化来求解方程( 7 ). 更具体地说,在Algorithm的第13行 s窗口 (见表 1 ,右),我们调用均匀化方法来近似
因此, P(P)
j个 = 我 + 是次随机转换矩阵,其中 λ
j个 = .使用与等式相同的计算( 16 ),我们得到了一个亚稳态矢量
哪里 L(左) 和 单位 截断点取决于 λ
j个 τ
j个 、和 此外,作为 λ
j个 仅取决于 W公司
j个 ,均匀化率通常小于全局均匀化率sup x个 ∈ S公司 λ
x个 ,这意味着等式中所需的项更少( 17 )比等式中的( 16 ).
整个过程的计算复杂性为 O(运行) ( ),因此,与全局均匀化相比,如果 ,其中 λ =支持 x个 ∈ S公司 λ
x个 和 ν
j个 是中非零元素的数量 P(P)
j个 .
Krylov子空间 Krylov子空间方法广泛用于大型特征值问题、线性方程组的求解以及矩阵指数和向量乘积的近似[ 46 , 47 ]. 我们对后一种近似方法感兴趣,并展示了它如何用于求解CME,无论是以全局方式还是与滑动窗口方法相结合。 最近,Sidje等人将Krylov子空间方法应用于CME[ 21 ].
全局Krylov子空间方法 回想一下,CME的全局解决方案如下所示 对 ( t吨 ) = 对 (0) e(电子) Qt(数量) 在续集中,我们描述了 e(电子) 助教 v(v) ,其中 A类 是一个 N个 × N个 平方矩阵和 v(v) 是长度的列向量 N个 。我们得到了近似值 对 ( t吨 ) 通过选择 A类 = ( 问 ) T型 和 v(v) = ( 对 (0) ) T型 首先让我们假设 t吨 = 1. 其主要思想是生成 米 -th Krylov子空间
并寻求近似解 e(电子) A类 v(v) 从这个子空间。 让 q个 最小值 是最低次的非零一元多项式,如下所示 q个 最小值 ( A类 ) v(v) = 0. 我们选择 米 ∈ ℕ 这样 q个 最小值 大于或等于 米 在这种情况下,向量 v(v) , A类 v(v) , ..., A类 米 -1 v(v) 线性独立,对于每个元素 x个 ∈ K
米 存在一个多项式 q个 最多度 米 -1和 x个 = q个 ( A类 ) v(v) 注意,在实践中,我们选择 米 =30或 米 =20,因为 q个 最小值 通常大于30。 然而,如果没有,问题可以解决 确切地 在中 d日 -第个Krylov子空间,其中 d日 是的度数 q个 最小值 .直接与基础合作{ v(v) , A类 v(v) , ..., A类 米 -1 v(v) }数值不稳定。 因此,我们构造了一个正交基{ v(v) 1 , v(v) 2 , ..., v(v)
米 }的 K
米 通过将Arnoldi算法应用于 v(v) , A类 v(v) , ..., A类 米 -1 v(v) .让 H(H)
米 成为 米 × 米 用Arnoldi算法计算的上Hessenberg矩阵 小时 米 +1, 米 是最后一个规范化值。 由 V(V)
米 我们表示 N个 × 米 带列向量的矩阵 v(v) 1 , v(v) 2 , ..., v(v)
米 。那么
哪里 e(电子)
k个 是一个适当大小的列向量,其 k个 -th分量为1,所有其他分量为零。 直觉上,等式( 18 )(b) 声明 H(H)
米 是的矩阵投影 A类 到上面 K
米 w.r.t.依据 V(V)
米 .近似值 e(电子) A类 v(v) 在里面 K
米 表示使用 V(V)
米 是 e(电子) A类 v(v) ≈ V(V)
米 年 ,其中 年 是大小向量 米 .
我们选择
从而产生近似误差[ 46 ]
哪里 ρ ==========================================================|| A类 || 2 是的光谱范数 A类 .等式中的近似值( 19 )仍然涉及矩阵指数的计算 H(H)
米 ,但作为 H(H)
米 尺寸较小且具有特定结构(上海森堡),这需要较小的计算工作量。 对于小矩阵的矩阵指数,可以应用Schur分解和Padé逼近等方法[ 31 ].
现在假设时间瞬间 t吨 是任意的,即我们想要近似 e(电子) 助教 v(v) 对一些人来说 t吨 > 0. 为了控制近似误差,我们计算 e(电子) 助教 v(v) 逐步利用它 对于 τ 1 , τ 2 ≥ 0. 对于步长 τ ,我们近似 e(电子) τA v(v) 由|| v(v) || 2 因为Krylov子空间与 A类 和 τA 是相同的,并且 根据公式( 20 )如果|| Aτ || 2 很小。
总之,Krylov子空间方法近似于 e(电子) 在 v(v) 通过动态选择步长的时间迭代向前推进 τ 1 , τ 2 ,.... 在每个迭代步骤中,我们计算一个向量
其中最初是 u个 0 = v(v) .矩阵 和 结果来自 我 -构造子空间正交基的Arnoldi算法的第次执行 跨度 { u个 我 -1 , A类 u个 我 -1 , ..., A类 米 -1 u个 我 -1 }。 当经过的时间等于 t吨 ,我们得到了 e(电子) 在 v(v) .
对于Krylov子空间方法的步长,一种流行的启发式方法是选择 τ 我 +1 取决于误差的估计
我 上一步的。 让 托尔 >0是预先指定的公差。 一个通用方案由三个步骤组成[ 36 ]. (1) 定义 ,(2)计算 u个
我 和误差估计ϵ
我 .(3)如果ϵ
我 > 1.2 托尔 拒绝 u个
我 ,更换ϵ 我 -1 带ϵ
我 ,然后转至步骤(1)。 对于我们的实验结果,我们使用了Expokit软件[ 48 ]其中小指数, ,通过不可约Padé近似计算[ 49 ].
局部Krylov子空间方法 假设我们在算法的第13行中使用了Krylov子空间方法 s窗口 (见表 1 ,右),近似值 (参见公式( 7 )). 通过出租 v(v) = , A类 = 、和 t吨 = τ
j个 我们可以应用与全局情况相同的过程。 注意,这会产生嵌套迭代,因为时间步 τ
j个 通常比Krylov子空间方法的时间步长大得多。 对于Krylov子空间方法,使用矩阵 问
j个 而不是 问 提供了重要优势。 Arnoldi过程的速度比 问
j个 通常包含的非零条目少于 问 同样,滑动窗口方法可能提供具有较小谱范数的矩阵|| 问
j个 || 2 这允许在Krylov近似过程中使用更大的时间步长,如我们的实验结果所示。
实验结果 我们在表中对这两种算法进行了编码 1 在C++中,并在3.16 GHz Intel双核Linux PC上进行了实验。我们讨论了示例1和示例2的实验结果,以及Goutsias的模型[ 50 ]和一个双稳态拨动开关[ 51 ]. Goutsias模型描述了噬菌体中抑制蛋白的转录调控 λ 涉及六种不同的物种和十种反应。 双稳态拨动开关是基因开关的原型,具有两个相互竞争的阻遏蛋白和四个反应。 图中列出了所有结果 2 .
如下文所述,我们还实现了Burrage等人提出的方法[ 21 ]为了在运行时间和精度方面与我们的算法进行比较。 此外,对于有限示例,我们将我们的方法与全局分析进行了比较,即在每个步骤中都考虑了整个状态空间。 我们没有将我们的方法与Gillespie模拟或基于Fokker-Planck方程的近似方法进行比较。 前一种方法仅提供概率分布的估计,如果估计小概率,则不可行[ 52 ]. 后一种方法不考虑分子数的离散性,并且已知在这里考虑的小种群情况下提供了错误的近似值[ 53 ].
参数 我们修正了输入ϵ=10 -8 算法的 s窗口 所有实验(见表 1 ,右侧)。 我们选择了输入 δ 以动态方式确保 j个 -第十步我们损失的概率不超过10 -5 · τ
j个 /( t吨
第页 - t吨 0 )通过限制在重要的州,也就是说,我们减少 δ 直到算法第4行之后 s窗口 成套设备 S公司
j个 最多包含10个 -5 · 概率小于前一组 S公司 j个 -1 .在图中 2 ,我们列出了使用的平均值 δ .
在下文中,我们给出了用于我们获得的实施例1和实施例2的结果的参数的细节。 对于剩下的两个示例,我们列出了相应的化学反应和我们为图中的结果选择的参数 2 .
酶示例 对于示例1,我们尝试了不同的参数集,称为集合a)-c)(参见图 2 ). 对于参数组合a),我们有 c(c) 1 = c(c) 2 = 1, c(c) 三 =0.1,从1000种酶和100种底物开始。 在这种情况下,可到达状态的数量为5151。 对于参数集b)和c),我们有 c(c) 1 = c(c) 2 = c(c) 三 =1,从100个酶和1000个底物以及500个酶和500个底物开始,分别产生96051和125751个可达状态。 每次我们根据时间选择时间范围,直到大部分概率质量集中在所有底物分子转化为产物的状态。 对于时间步 τ
j个 在算法中 s窗口 ,我们应用上述条件。
我们考虑等式中迭代的四个分支( 11 )为了确定状态变量的上下限。 (1) 为了估计复合分子的最大数量(以及酶群的最小数量),我们实施了更多类型的反应 R(右) 1 比平均水平高( κ 1 = ),类型较少 R(右) 2 和 R(右) 三 ( κ 三 = 和 κ 2 = ). (2) 通过考虑较少类型的反应 R(右) 1 ( κ 1 = )和更多类型 R(右) 2 和 R(右) 三 ( κ 三 = 和 κ 2 = )复合群体变得最小(酶群体最大)。 (3) 最小类型数的估计 P(P) 分子(以及类型的最大数量 S公司 分子)通过执行更多类型的反应获得 R(右) 2 ( κ 2 = ),类型较少 R(右) 1 和 R(右) 三 ( κ 1 = 和 κ 三 = ). (4) 最后,更多类型的反应 R(右) 1 和 R(右) 三 ( κ 1 = 和 κ 三 = ),类型较少 R(右) 2 ( κ 2 = )最大限度地增加产物分子的数量(并使底物分子的数量最小化)。
以酶为例,如果初始条件是固定的,那么一种状态是由至少两个条目唯一确定的,比如复合物和产物分子的群体。 然而,如果复杂分子的预期数量较高,矩形窗口形状的结果较差。 原因是在这种情况下,概率质量位于对角线上(参见图 三 ). 如果有效状态集被矩形窗口捕获,则它可能包含许多不重要的状态。 通过考虑窗口构造过程中所有状态变量的边界以及守恒定律,可以避免这个问题。 更准确地说,图中的平行四边形 三 通过计算每个值来构造 属于 P(P) 上下限 锿 通过 和 ,其中 年 = ( 年 1 , 年 2 ,0,0)是初始总体向量 和 人口的上限和下限是 E类 , S公司 , 锿 、和 P(P) .
注意图中的平行四边形 三 是由系统的守恒定律引起的。 一般来说,应该考虑守恒定律,因为否则窗口可能与守恒定律不一致,即它可能包含无法到达的状态。
基因表达示例 在图中 2 我们给出了示例2的结果。 参数集a)和参数集b)(称为集合a)和集合b)的区别在于,对于a)我们从空系统开始,对于b)我们从100 mRNA分子和1000蛋白质开始。 对于这两种变量,我们都选择速率常数 c(c) 1 = 0.5, c(c) 2 = 0.0058, c(c) 三 = 0.0029, c(c) 4 = 0.0001. 我们使用的时间步长由时间间隔部分中的条件决定。 请注意,我们无法使用全局方法求解此示例,因为可达状态的数量是无限的。 专栏 错误 包含总错误 η
第页 (参见公式( 8 ))以及 次(秒) 指以秒为单位的运行时间。 在列中 百分比 。我们列出了用于窗口构造的总运行时间的百分比。 专栏 平均风 . 大小 指窗口中的平均状态数。
对于基因表达的例子,我们使用了四个分支:我们通过选择 和 并将其最小化 和 .反应 R(右) 2 和 R(右) 4 与这个物种无关。 我们通过选择最大化蛋白质群体 、和 并将其最小化 、和 .
Goutsias模型 该模型在图中称为Goutsias模型 2 ,由以下化学反应组成[ 50 ]:
1: RNA T RNA+M
2: M T?
三: DNA。 D T RNA+DNA。 D类
4: RNA T?
5: DNA+D T DNA。 D类
6: DNA。 D T DNA+D
7: DNA。 D+D T DNA.2D
8: DNA.2D T DNA。 D+D
9: M+M T D公司
10: D T M+M
我们使用了与Goutsias相同的动力学常数[ 50 ]和Sidje等人[ 21 ]以及相同的初始状态。
下面,我们列出状态变量上界的分支。 如果分别考虑相反的组合,则可获得下限。 我们参考图 4 以便于说明简化分支选择的反应之间的依赖性。 我们通过选择组合来最大化RNA种群 .我们通过选择组合来最大化单体数量 。我们通过选择组合来最大化二聚体分子的数量 注意,虽然反应5消耗二聚体,但选择 最大化系统中二聚体的数量。 这是因为反应5是生成单体和二聚体所必需的。 使用滑动窗口方法,我们永远不会耗尽内存,但在较长的时间范围内,运行时间可能会非常长。 原因是窗口很大,因为系统在稍后的时间包含许多单体和二聚体。 对于图中的结果 2 我们一直在考虑这个系统 t吨 =300,而Sidje等人[ 21 ],最长时间范围为 t吨 = 100. 在图中 5 我们绘制了物种的分布图 M(M) 和 D类 .
双稳态拨动开关 拨动开关涉及两种化学物质 A类 和 B类 和四种反应。 让 x个 = ( x个 1 , x个 2 ) ∈ 。反应如下 ∅ → A类 , A类 → ∅ , ∅ → B类 , B类 → ∅ 及其倾向函数 α 1 , ..., α 4 由提供 α 1 ( x个 )= c(c) 1 /( c(c) 2 + ), α 2 ( x个 )= c(c) 三 · x个 1 , α 三 ( x个 )= c(c) 4 /( c(c) 5 + ), α 4 ( x个 )= c(c) 6 · x个 2 注意,在本例中,倾向函数不是等式中描述的形式。 1 对于我们的实验结果,我们选择了与Sjöberg等人相同的参数[ 23 ]也就是说, c(c) 1 = c(c) 4 = 3·10 三 , c(c) 2 = c(c) 5 = 1.1·10 4 , c(c) 三 = c(c) 6 =0.001,以及 β = γ = 2. 初始分布为高斯分布 ( μ , σ 2 )带有 μ =(133133) T型 和 我们考虑了明显的四个分支,每个分支都旨在最小化/最大化这两个组件中的一个。 分支最小化 A类 例如将具有较少的第一反应和较多的第二反应。