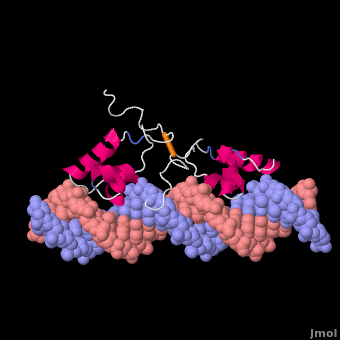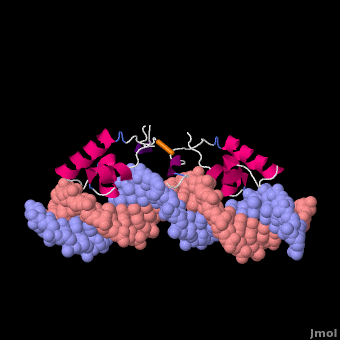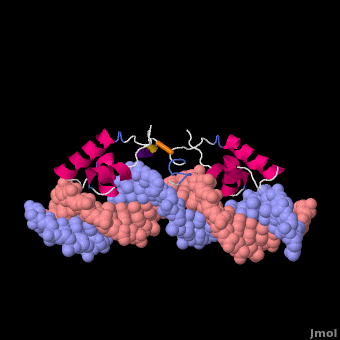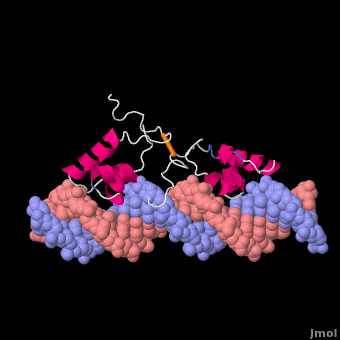
莫尔夫的紫胶阻遏物与DNA复合
显示了非特异性结合(直接DNA)与操作员序列的特异性识别(扭结DNA)之间的差异。这种结合是使DNA发生扭结,还是仅仅使预先存在的扭结稳定下来,尚不清楚。详细信息如下.
紫胶抑制因子是什么?
阻遏物是抑制基因; 也就是说,它们抑制信使RNA来自他们的目标基因。每一个阻遏物通过识别特定的DNA序列(称为操作人员在里面细菌抑制蛋白的编码管理的基因。
这个乳糖(“lac”)阻遏物控制参与糖乳糖代谢的细菌酶的表达。当lac阻遏物与乳糖结合时,它会变成一种不活跃的构象,不能抑制这些酶的产生。因此,使用乳糖所需的酶只有在乳糖可用时才能制造。lac阻遏物及其控制的一组基因,称为操纵子,是第一个被发现的这样的基因调控系统。1960年描述了歌剧院[1]弗朗索瓦·雅各布等。他还正确地提出了lac阻遏物调控的一般机制。这个1965年诺贝尔生理学或医学奖弗朗索瓦·雅各布(François Jacob)、安德烈·卢夫(AndréLwoff)和雅克·莫诺德(Jacques Monod)“因其在酶和病毒合成的遗传控制方面的发现”而被授予该奖项。
有关紫胶阻遏物的一般介绍,请参阅David Goodsell的紫胶阻遏物简介在他的系列中本月最佳分子以及维基百科上关于紫胶阻遏物Mitchell Lewis在2005年发表了一篇详细的评论[2]。另请参阅转录和RNA处理.
紫胶阻遏物的结构
紫胶抑制蛋白(显示链A1磅,分辨率4.8Ω),从N末端开始,以DNA-绑定“耳机”,后跟一个铰链区域,然后是N端配体结合子域和aC末端配体结合亚结构域,一个连接器和C端子四聚螺旋[3]. ()在缺乏DNA的情况下铰链区域没有形成此处所示的α螺旋。
当链条处于
每个配体结合亚结构域由两个不连续的片段组成。
紫胶阻遏物形成.二聚埋2200°2表面,包括,
疏水的,极地的
形成疏水核(如图所示1磅,分辨率2.7μl,由于混乱).
最高的是接触的表面DNA[4](此处仅显示α-碳原子,没有侧链,因为侧链在4.8℃下未被分解1磅型号。)二聚化表面是配体结合域[5](此场景显示侧链,使用中的2.7º模型1磅/平方英寸,由于无序,缺少DNA结合域。)
C末端四聚体螺旋系住两个二聚体,从而形成有两个DNA-结合位点。
DNA结合:操作员的一个扭结
非特定绑定
Lac阻遏物与DNA非特异性结合(派生的[6]从1盎司, 20核磁共振模型)使其能够沿着DNA双螺旋快速滑动,直到遇到lac算子序列(“促进扩散”[7]). DNA绑定域使用螺旋转螺旋基序(阿尔法螺旋,转动). 在非特定绑定期间铰链区域无序(由20个模型的位置范围表示)。这个DNA双螺旋在此处所示的模型中被描述为直线(参见方法),但实际上,直线度可能随顺序而变化(参见在下面). 右图所示的蛋白质模型(1盎司)有两个DNA-binding域副本铰链区域(以区分链条B铰链).这20个核磁共振模型模拟无序铰链区域的热运动。
特定绑定
在识别出特异性操作序列后,非特异性结合转化为(衍生的[6]从110万, 20核磁共振模型). 在这个转换过程中,铰链区域从无序循环变为阿尔法螺旋(:关闭旋转以查看高亮显示)与DNA的小凹槽结合。如下所述,此绑定使扭结(“弯曲”) DNA双螺旋构象。在缺乏结合的lac阻遏蛋白的情况下,该DNA序列处于扭结状态的时间百分比尚不清楚,但这可能是一个很大的百分比(见下文)。这20个核磁共振模型可以与非特异性结合的动画进行比较。请参见Lac阻遏物变形方法.
DNA识别
蛋白质通常识别主槽通过直接读出也就是说,通过与暴露在主沟中的DNA碱基边缘形成序列特异性氢键[8].
DNA序列识别小凹槽通常伴随着DNA的扭结或弯曲,则更为复杂。直接读出不太重要,因为与主槽不同,四个碱基在次槽中没有独特的氢结合表面[8].识别DNA的形状似乎更重要[9][10]在许多情况下,人们认为阳离子精氨酸会被吸引到较小凹槽的一个区域,该区域的镍离子电荷密度较高,这是因为凹槽变窄所致[10]在这些情况下,蛋白质似乎能识别DNA小凹槽的形状(间接读出)[10].
在含有特定DNA的紫胶阻遏物复合体中,一对精氨酸(每条链中的Arg51)靠近小沟,但指向小沟之外(). 因此,精氨酸与狭窄的小沟的结合似乎与紫胶阻遏物的特定DNA识别无关。
DNA扭结
严格地说,弯管DNA中的扭结DNA据说是扭结的当两个相邻的碱基对之间的堆叠接触被破坏时[8]纽结两侧的DNA可能是直的或弯曲的。A类:单个CpG基对与相邻的CpG基对部分分离。嘧啶-嘌呤碱基对的堆叠相互作用最弱,最容易发生扭结[8]在lac阻遏物与特异性DNA的复合物中,(如果场景是空白的,)部分插在分离的CpG碱基对之间,这有助于稳定扭结。通常情况下,在蛋白质结合之前,DNA中存在序列相关的扭结和弯曲[8]DNA结构是动态的。例如,最近观察到Hoogsteen碱基配对与Watson-Crick碱基配对暂时平衡[11](请参见新闻和视图[12]). 此外,p53与部分但不是全部DNA序列的结合可以稳定Hoogsteen(而不是Watson-Crick)碱基配对[13]因此,中描述的“弯曲”(实际上是扭结)这一页上的变形可能会给人错误的印象:lac阻遏物结合可能只是稳定同源DNA序列中预先存在的扭结(或短暂扭结)。
DNA弯曲
严格地说,弯管在DNA中与扭结.弯曲是指分布在几个相邻的碱基对[8],而a扭结(见上一节)是堆叠中的中断,仅限于一对相邻的碱基对。安在与HPV E2蛋白复合的DNA中发现(由于[8]对于本例,1jj2(日本)).
转化形态
这个通过以下方式平滑设置动画时,可以更容易地看到它们变形.(中给出了用于创建此变形的方法Lac阻遏物变形方法.)
- 注意DNA的扭结,凸面上的中央小沟变宽。同源DNA序列在lac阻遏物结合之前是否(可能是暂时的)扭结尚不清楚(见上文)。
- 还要注意二硫键合铰链区域环向α螺旋的转换。(在变形插值中为每个模型计算显示的二级结构。)
尽管如此,DNA中lac算子序列的特异性识别在很大程度上发生了氢键.在变形的渲染中进行了说明。显示的氢键包括Arg22.N-eta2和Tyr18.OH与主沟中的DNA碱氧相互作用,以及Ala53.O与小沟中的DNA碱氮相互作用。(未显示所有相关氢键;参见方法.)
Powerpoint®幻灯片动画
这是一个动画multi-gif真正的电影已准备好插入Powerpoint®[14]幻灯片。如果下面的图像没有移动,请重新加载此页面(50个循环后停止)。
- 在Windows中,只需将电影拖放到Powerpoint幻灯片中。然后,您可以调整大小并定位它。当您将“视图”更改为“幻灯片放映”(“投影”)幻灯片时,应该播放电影。
- 在Mac OSX中,按住Ctrl键单击电影,然后单击“保存图像”。在Mac Powerpoint中,在所需的幻灯片上,使用插入菜单(位于顶部)并选择电影。。。,然后插入保存的.gif电影文件。插入影片后,请确保“工具箱”正在显示(由窗口顶部的图标按钮控制)。现在可以调整影片的大小和位置。单击幻灯片中的电影将其选中。现在,在工具箱/格式调色板的“电影”下,选中“循环直到停止”。现在,当您将“视图”更改为“幻灯片放映”(“投影”)幻灯片时,应该播放电影。
挑战你的理解力
以下是一些问题,挑战您的理解。
- 为什么lac阻遏物非特异性地与DNA结合?
- 当紫胶阻遏物非特异性地与DNA结合时,它与DNA双螺旋的哪一部分结合?
- DNA有净电荷吗?如果有,在pH值为7的水溶液中是负电荷还是正电荷?
- 抑制蛋白与DNA的非特异性结合可能涉及哪些化学键?
- 紫胶阻遏物与DNA的特异性结合是否破坏了DNA链中碱基对之间的Watson-Crick氢键?
- 像紫胶阻遏物这样的蛋白质如何识别DNA双螺旋中的特定核苷酸序列?
- 阻遏蛋白与DNA的特异性结合涉及何种化学键?
- 紫胶阻遏物能识别DNA大槽或小槽中的特定碱基吗?
- 当它识别其特定核苷酸序列时,lac阻遏物如何稳定DNA双螺旋中的扭结?
如有要求,可向 。如果您希望我们在Proteopedia中公开答案,请告知我们。联系我们时,请提供您的全名、职位、机构或学校以及位置。
。如果您希望我们在Proteopedia中公开答案,请告知我们。联系我们时,请提供您的全名、职位、机构或学校以及位置。
内容归属和确认
此处显示的变形最初是由埃里克·马茨2004年的网页与DNA结合的Lac抑制子,在蛋白质探索者。组织.
埃里克·马茨谢谢Remo Rohs公司感谢他对本文2010-2011年更新的亲切和专业建议。